Grasslands have been estimated to cover approximately 40% of global land surface area, approximately 5.25 bn ha (~13 bn ac ), except for Greenland and Antarctica [Suttie 2005; White 2000:12]. Their deep soils are rich repositories of nutrients, especially carbon and water. Many grasslands are anthropogenic, i.e., resulting from various land management techniques to maintain land for grazing and crop production by humans. Virgin grasslands are increasingly rare, possibly leading to significant underestimations of their potential positive contribution to productivity, and to carbon and water storage. Grasslands are important repositories of biodiversity, and have significant impacts on weather and climate. Here we review research and articles that indicate soil carbon storage potentials of roughly 13 gigatons per year (the equivalent of 6.5 ppm) if global grasslands were managed regeneratively.
Overview
While we have separate sections for Grasslands, Croplands and Soils, there is inevitable overlap. Many croplands are modified grasslands, and both are, of course, based in soils. Yet there are enough differences in each area of study to merit separate sections, keeping in mind that systemic behaviors and interactions are broadly applicable.
Since the onset of agriculture over 10,000 years ago with land management techniques that expose soil to air, estimates of up to 537 gigatons of soil carbon have been oxidized to carbon dioxide and other greenhouse gases [Buringh 1984: 91]. Even so, soils (>2,300 Gt) currently hold almost as much carbon as plants (550 Gt), atmosphere (800 Gt) and ocean surface waters (1,000 Gt) combined [NASA 2011], and almost surely retain the potential to store enough atmospheric carbon to return to pre-industrial levels.
Typical soil studies examine the first 30 cm (1 ft) of soil depth, but more recent investigations indicate that major soil carbon storage takes place deeper than that, often in a more stable form [Liebig 2008, Follett 2012, Harper 2013]. A USDA paper found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr) [Follett 2012]. A study of switchgrass for bioenergy found rates of SOC increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft) [Liebig 2008]. On an intensively grazed former row-crop agriculture land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions [Machmuller 2015]. In addition, the ultimate methane emissions may have been markedly less than measured since the report did not consider methane breakdown into CO2 from methanotrophic bacteria and atmospheric hydroxyl radical oxidation, with a significant reduction of methane’s ultimate greenhouse gas impacts.
These reports demonstrate the potential for massive amounts of soil carbon storage, significant cooling of the biosphere, and dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Grassland Evolution
Grasslands have long been a rich repository of carbon, both stable and labile. The co-evolution of grasslands with grazing ruminants has contributed to dramatic global cooling over the past 50 million years as a result of significant photosynthetic carbon drawdown into grassland soils [Retallack 2013]. Thus, grasslands are more than a consequence of geophysical changes, they are
. . . a biological force in their own right (Retallack 1998), in some ways comparable to the human rise to dominance of planetary resources (Vitousek et al. 1997). Grasslands have long been considered products of the coevolution of grasses and grazers (Kovalevsky 1873). Few plants other than grasses can withstand the high-crowned, enamel-edged teeth and hard hooves of antelope and horses. Yet these same animals are best suited to the abrasive gritty opal phytoliths and dust of flat, open grasslands. Grasses recover readily from fire and nurture large herbivores such as elephants: both fire and elephants promote grassland at the expense of wood land (Retallack 1997b; Jacobs et al. 1999). Grasses suppress insect and fungal attack with secondary metabolites such as cyclic hydroxamic acids (Frey et al. 1997). Grasses create Mollisols, unique soils with fine crumb clods rich in organic matter (Retallack 1997b). (Retallack 2001:407) [Emphasis added.]
Occupying such vast areas of planetary land surface, grasslands have a major influence on the global climate:
CO2 and CH4 (which rapidly oxidizes to CO2 ) are important greenhouse gases, and mechanisms for burial of their C may result in climatic cooling (Berner 1999). The most important long-term C sink from grasslands is their supply by erosion to sedimentary basins of crumb peds, which are unusually rich in organic matter intimately admixed with clay (Pawluk and Bal 1985). Tropical forests, in contrast, yield highly oxidized spherical micropeds with virtually no organic content (Retallack 1991a).” (Retallack 2001:414)
While there are other soils that are greater carbon sinks, such as peat bogs, wetlands and coastal habitats (e.g., mangroves, seagrasses), for volume and depth of carbon storage on vast areas of land, grasslands have enormous potential:
Grasses themselves are C sinks, especially considering their mass of roots and rhizomes underground. . . . [Numerous soil investigations] indicate that past estimates of organic C in tropical grassland soils have been low, in part because soils were not analyzed to sufficient depths. . . . Grassland and woodland soils may have comparable amounts of organic C in the surface 15cm. Beyond that depth, organic C values drop off dramatically in woodland soils but remain high in grassland soils to a meter or more. The fine structure and fertility of grassland soils is in large part due to this large C reservoir. (Retallack 2001:415)
Conventionally, it is estimated that approximately 40% of global land surface area is grasslands (52.5 million square kilometers, or ~5.25 billion hectares, or ~13 billion acres [Suttie 2005]), except for Greenland and Antarctica [White 2000:12; see Figure 1, below]. This is likely a significant underestimation of soil surface area and volume, since grasslands are not uniformly flat, with topographical variations adding carbon, water, etc. to areas that are typically calculated on the basis of a two-dimensional map projection [Blakemore 2016: fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regeneration, carbon capture and water storage than is conventionally assumed (see the section, “Do We Have More Soil for Carbon Storage than We Thought?”).
Natural grasslands are typically areas of low and seasonal rainfall. Unlike temperate environments with year-round rainfall, semi-arid and arid grasslands are dependent on grazing animals as a keystone species. The habits of grassland plants are as dependent on grazing animals as the animals are dependent on the plants that grasslands provide as food.
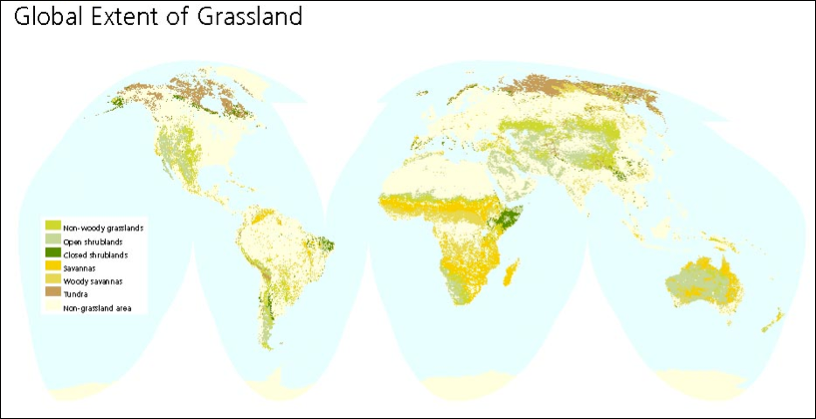
Figure 1: Global Extent of Grassland (White 2000:12).
Grazing animals are ruminants and as long as they are grazed in herds that move frequently, as they do in natural habitats in the company of predators, ruminant species are often interchangeable insofar as grassland health is concerned. Bison and antelope, for example, may be the wild ruminants that roamed the prairies and savannahs, but domestic cattle, when properly managed (human herders are the equivalent of wild predators), serve the same ecosystem functions.
Pioneering Work Of Allan Savory
Allan Savory, a wildlife biologist from Zimbabwe, began studying desertification in the 1950s. He pioneered an approach that he has termed Holistic Planned Grazing (HPG) for regenerative management of grasslands. He noted that there are essential differences between temperate grasslands, which he termed “non-brittle” environments, and arid and semi-arid grasslands, which he referred to as “brittle.” These distinctions are critical in understanding how different habitats require different management approaches.
Non-brittle environments, because of year-round rainfall, are relatively forgiving of mismanagement that destroys soil biota and exposes soil to sunlight, air and the elements. Recovery from soil degradation can be be rapid. Brittle environments, to the contrary, are fragile and easily desiccated, and when poorly managed, either from overgrazing or undergrazing, may take decades or centuries to recover or even ultimately turn to desert.
Brittle environments particularly need ruminant evolutionary partners. Their hooves are designed to open the soils to air and water, and their digestive systems deposit a feast for soil organisms. The ruminant gut is a moist refuge during the dry season for soil microbes which are essential to the health of the land. Grasses need to be bitten lest they shade out their own new growth.
What Savory discovered is that the same land may either flourish or die depending on how it is grazed. When ruminants are kept in check by predators they graze an area in tight herds for protection and then move to the next patch of fresh grasses and other plants, providing up to two years of recovery and regrowth time for the recently grazed pasture. On the other hand, when they are provided the safety of fencing and left to graze large areas at will, they return to their favorite plants and overgraze those areas, eventually compacting the soil, preventing water infiltration and proper aeration, killing the plants, and leading to desertification.
The difference in land health is dramatic.[19] The pictures below illustrate:


Mexico


Arizona


Zimbabwe
Fig. 1. These pictures are of neighboring properties in Mexico, Arizona and Zimbabwe. In each area they were taken on the same day, have similar soils, and the same precipitation. The pictures on the right are examples of properly managed livestock through Holistic Planned Grazing to restore grasslands. On the left we see examples of improperly managed livestock as well as exclusion from grazing (“resting the land”) [Savory Institute 2015:12]
Savory’s work, after decades of successful application on ranches in Africa, Asia, Australia and North and South America, garnered global attention (and controversy) after his TED Talk in 2013 [Savory 2013].
The primary point is that If well-managed, grassland soils can not only sequester annual greenhouse gas emissions but can also begin to draw down legacy atmospheric carbon as well. In addition, they provide human and other predator food, converting grasses inedible to non-ruminant mammals to meat.
Grasslands As Ecosystems
It is useful to understand how grasslands work as intact ecosystems, thereby providing a solid theoretical basis for observations of grasslands as vast carbon and water sinks. What appears above-ground is only a hint of grassland ecosystem dynamics. Soils are the planet’s most complex and least understood terrestrial ecosystem, yet soils are where most of the action takes place on grasslands as well. While all soils are built on the foundation of minerals provided by weathering of rock, the soils on grasslands are primarily biological soils. The kingdoms of life are the active agents in soil creation and it is the interactions among life forms that create the rich and productive grassland soils (see Soils section).
The basis for all life is the microbial kingdom. These smallest of cells, with their complex biochemistry, morphology and behavior, are active players in creating stable soil molecules, storing abundant carbon and water. In addition to microbes, soil ecosystems are built from exchanges among fungi, insects, worms, green plants, birds and small and large mammals. It is this set of rich interactions that creates the biodiverse, abundant and resilient environment of global grasslands.
For the sake of illustration, let us start the discussion with the ruminant gut during a dry season. Microbes survive in a warm, moist environment while constantly being cycled into the soil through digestion and elimination. The short-term hoof disturbance with minimal compaction while animals are constantly moving opens the soils to available moisture from precipitation, urination and condensation. During the rainy season the water is more effectively absorbed into opened soils, nourishing plants, raising the water table and eventually even leading to perennial streams and ponds. Even limited rainfall goes a long way in spongy soils that are covered with grasses and other plants to keep the ground cool and moist [Byck 2014: 8’38”]
Methane
Methane, a relatively short-lived but powerful greenhouse gas, is often raised as a serious concern with beef production. This is surely true when animals are left to roam freely and overgraze, and then moved to concentrated animal feeding operations with large manure lagoons. However, it is important to consider the whole of ecosystem functions in assessing methane emissions {Savory Institute 2015]. This includes the conversion of the methane molecule into carbon dioxide by bacteria (metanotrophs) that live in healthy soils and literally eat energy-rich methane, and the oxidation of methane by hydroxyl radicals present in the lower atmosphere. It may include other ecosystem processes, such as the effects of earthworms increasing methanotrophic bacterial activity in landfills and pastures [Héry 2008; Kernecker 2014]. The result is a virtuous cycle where plants can then take up that carbon dioxide through photosynthesis and send some of the carbon back underground through their root systems. Throughout their life cycle under conventional industrial management, cattle are rarely if ever exposed to such healthy, biodiverse soils.
Historical methane data indicates that in the United States, for example, pre-settlement wild ruminants generated approximately 86% of the methane of current farmed ruminants (Hristov 2012:1371). Yet even with vast numbers of ruminants on grasslands across the planet, atmospheric methane remained constant until the global dependence on widespread use of fossil fuels and its effects on agriculture and animal husbandry began to grow rapidly in the 18th century (Fig. 2).
Thus, results are very different with animals grazed in a manner that mimics nature. Rowntree et al. describe the importance of accounting for the beneficial ecosystem services that well-managed grazing systems can provide.
. . . LCA’s [Life Cycle Assessments] often consider soil C to be in dynamic equilibrium. However, empirical data suggest otherwise (e.g. Machmuller et al., 2015; Teague et al., 2011). Recent studies such as Ripple et al. (2014) and Eshel et al. (2014) have reported the emissions from ruminants in food production without accounting for the beneficial ecosystem services that well-managed grazing systems can provide. In our study, we used 3 tC/ha/yr (1.2 tC/ac/yr) as a potential C sequestration figure, which is relatively high (Conant et al., 2001) but viable based on existing studies (Teague et al., 2011; Delgado et al., 2011; Machmuller et al., 2015; Teague et al., 2016). Importantly, the results presented here suggest that with appropriately managed grazing, a grass-finished beef model can not only contribute to food provisioning but also be ecologically regenerative as well. [Rowntree 2016:36]
This excerpt illustrates a paradigm shift in action, wherein investigators within the dominant paradigm are constrained from evaluating the possibilities offered by “beneficial ecosystem services” because it doesn’t occur to them to consider them. Such biological processes are invisible due to limiting assumptions of the paradigm.
Research into systems implications of holistically managed grass-finished beef is growing. It is only recently that mainstream researchers are beginning to understand that the biological function of animals in an ecosystem is as dependent on the ecosystem as it is on the biology of the animal.[20] Studying animals in isolation or as part of a synthetic system such as industrial agriculture often leads to incorrect conclusions.
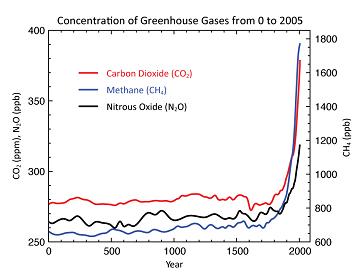
Fig. 2. Historical Concentrations of Greenhouse Gases [IPPC 2007]
Review of some studies of grasslands
Soil creation (pedogenesis) is conventionally defined as the weathering of rock; it may take 3,000 years or longer to create a foot of soil through geological processes. Soil created through biological activity, on the other hand, happens orders of magnitude faster, up to several inches per year.
Australian soil scientist Christine Jones notes that
The rates of soil formation provided in the scientific literature usually refer to the weathering of parent material and the differentiation of soil profiles. These are extremely slow processes, sometimes taking thousands of years. Topsoil formation is different and can occur rapidly under appropriate conditions. . . .
The late P.A. Yeomans, developer of the Keyline system of land management, recognised that the sustainability of the whole farm was dependent on living, vibrant topsoil. The formation of new topsoil using Keyline principles, at rates not previously considered possible, was due to the use of a tillage implement designed to increase soil oxygen and moisture levels, combined with a rest/recovery form of grazing and pasture slashing, to prune grass roots and feed soil biota, especially endemic earthworms. Yeomans was able to produce 10 cm of friable black soil within three years, on what was previously bare weathered red shale on his North Richmond farm (Hill 2002).
Bennett (1939) calculated a rate of topsoil formation of just over 11 t/ha/yr (4.4 t/ac/yr) for soils in which organic material was intermixed into surface layers. In situations where plant root mass is high, rates of topsoil formation of 15-20 t/ha/yr (6-8 t/ha/yr) have been indicated (Brady 1984). Healthy groundcover, high root biomass and high levels of associated microbial activity, are fundamental to the success of any technique for building new topsoil.
If the land management is appropriate, evidence of new topsoil formation can be seen within 12 months, with quite dramatic effects often observed within three years. Many people have built new topsoil in their vegetable or flower gardens. Some have started to build new topsoil on their farms. If you have not seen new soil being formed, make a point of doing so. (Jones 2003:19-20)
Healthy biodiverse grasslands with abundant animal populations provide favorable circumstances for biological soil accumulation and carbon sequestration, including opening soils to air and water, fertilizing soil life and stimulating growth of grasses.

Fig. 2. “Root Systems of Prairie Plants,” Heidi Natura, Conservation Research Institute, n.d., http://kmlandtrust.org/pdf/NPGpp5-6-11×17.pdf
Using current best land management practices, recent research has begun to confirm the importance of studying soil organic carbon accumulation on grasslands. In a 9-year study of bioenergy crops, investigators found that switchgrass and maize stored 50% of their soil organic carbon (SOC) below 30 cm (1 foot), up to 4 times more than used in models in use at that time (Follett 2012:866):
Most of the research on SOC in agricultural production systems focused on C in the 0 to 30 cm depth [22–27]. A few studies in which soil sampling has been conducted at greater depths indicate that production agriculture affects soil C deeper in the soil profile [28,29]. (Follett 2012:867)
Concerns about soil depth measurement are not new [Liebig 2008]; depth of soil measurement in estimating soil carbon storage potential is a significant issue. Conventional soil science, which largely addresses agricultural soils managed in industrial agricultural contexts, typically measures soil carbon down to around 30 – 40 cm (12 – 16 inches). Yet roots of native prairie plants may reach 5 times that depth (see Fig. 1), storing carbon in stable molecules for centuries and millennia as long as the soils are undisturbed and not exposed to light or desiccation. [See Soils section.]
Methodological issues for assessment of SOC have thus been problematic, and have likely led to serious overall underestimation of soil sequestration capacity. This is particularly relevant because these soils will not be able play their appropriate critical role in addressing climate until mainstream science and policy recognize and promote the potential of best practices in land management in all ecosystems. Harper & Tibbett found up to five times more soil carbon in Australian soils at depths greater than 1 meter (~3 feet) than is conventionally estimated:
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths (Table 1), it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5 m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm −2 or 21 % of the total store to bedrock. [Harper 2013: 645]
We discuss the dynamics of water cycling and forests elsewhere, but it is worth noting a recent paper suggesting that a more holistic view of ecosystem dynamics is in order. A paradigm shift prioritizing water over carbon as the driving climate force more accurately and effectively guides climate recovery strategies and offers more tactical and regenerative options [Ellison 2017; s.a. Schmidt 2017]. Water is more tangible to most people than carbon, and water recovery is more visible and rapid, offering hope and encouragement in a generally grim scenario. In addition there are numerous other benefits to people and landscapes with improved water management.
THE IMPORTANCE OF ANECDOTAL EVIDENCE
While anecdotal evidence is often disdained in academic science, in many scientific pursuits it forms the foundation of future study. This is especially true in such predominantly observational pursuits such as naturalist biology, ecology, rangeland science and agronomy, where study of the visible vagaries of the natural world inspires questions that may reach far beyond what a happenstance occurrence would imply. It is the weight of such observations that leads to formal protocols, hypotheses and theories to explore the details of a field. Isolating variables can be a very helpful tool; unfortunately, modern science practice has generally lost sight of systems contexts, and that system behavior can be very different from the behavior of any of its isolated parts.
Therefore, it is essential to embrace both analytical and holistic evidence in order to build a full understanding of how environmental systems work as wholes. Together these complementary approaches provide a more comprehensive picture of the systems in question, as well as much clearer guidance for how to proceed in current global ecosystem urgencies.
A small selection of a growing literature of informative anecdotal reports of grassland eco-restoration is included among formal studies below. They reflect the extensive experience of farmers, ranchers and other land managers, and demonstrate the potential positive effects of regenerative land management and eco-restoration on climate and the biosphere as a whole. See for example Stigge 2016, Oppenheimer 2015, Byck 2014, Brown n.d., Brown 2016.
Grassland Article Summaries
Byck 2014. This 12-minute video relates the experiences of three ranchers who manage cattle and land according to regenerative land management principles. They discuss their transition to Holistic Planned Grazing, where for two of them, in areas of ~15 inches of rainfall, their alternative had been bankruptcy. The video vividly illustrates the benefits of cover-cropping and organics, the improvement in lifestyle and economics, and the dramatic improvement in biodiversity and water management. [Byck 2014]
Follett 2012. A USDA study found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr). Ausmus reports that
. . . a 9-year project that evaluated the effects of nitrogen fertilizer and harvest treatments on soil organic carbon sequestration in switchgrass and no-till maize crops managed for biofeedstock production [found that] more than 50 percent of the soil carbon was found between 1 and 5 feet below the soil surface. The average annual increase of soil organic carbon throughout the first 5 feet of subsoil also exceeded 0.9 tons per acre per year [Ausmus 2014: 4-5].
Of interest were the difficulties the authors faced in getting the study through peer review and published since their results were so unexpected, as reported in Ausmus 2014.[21] It originally appeared in Bioenergy Research in 2012. [Follett 2012].
Note too that the Follett study was performed on already degraded soils, “Perennial grasses could be used as bioenergy crops on about 20 million ha (ha = 10,000 m2 or 2.5 acres) of marginal or idle cropland in the USA alone [18]” (Follett 2012:867). [Emphasis added.] A well-supported inference is that healthy, biodiverse soils will yield even better results.
Degraded soils may be less effective carbon sinks than virgin soils even though they have lost most of their carbon because the soil life that creates long-lasting stable carbon molecules is damaged or destroyed by synthetic inputs, tilling and other forms of mismanagement. Nonetheless, results were dramatic:
In the first 9 years of a long-term C sequestration study in eastern Nebraska, USA, switchgrass and maize with best management practices had average annual increases in SOC per hectare that exceed 2 tC/yr for the 0 to 150 cm soil depth. For both switchgrass and maize, over 50 % of the increase in SOC was below the 30 cm depth. SOC sequestration by switchgrass was twofold to fourfold greater than that used in models to date which also assumed no SOC sequestration by maize. (Follett 2012:866) . . . .
Our results clearly show that significant amounts of C were sequestered deep in the soil profile by switchgrass grown and managed as a biomass energy crop and maize grown continuously in a no-tillage production system for the cultivar Trailblazer array of N fertility and harvest treatments for a 9-year period. For almost all other C sequestration reports used in bioenergy models, studies designed for other purposes were adapted to obtain soil C sequestration estimates, initial soil samples were not available, and control samples were from adjacent fields or non-treatment areas. Our results are supported by similar results reported by Liebig et al. [28] for four switchgrass fields managed with uniform N rates and harvest treatments for 5 years in the USA western Corn Belt. They are supported by the recent work of Varvel and Wilhelm [29] for maize in which significant increases in soil C occurred in soil layers up to 150 cm in depth in maize no-till plots as compared to tilled plots. The soil C that is sequestered deeper than 30 cm is expected to be more stable over time since it is below the tillage zone. Even in the top 30 cm of soil, sequestered C may be stable for extended periods in no-till production systems as reported previously by Follett et al. [33]. (Follett 2012:873)
Harper 2013. This study suggests that the standard sampling depth of 30cm vastly underestimates the global store of soil organic carbon, and thus, presumably, the potential of future soil carbon storage that could result from eco-restoration efforts. “Hypothesizing that SOC retained in soils below the top half metre (in highly weathered deep profiles) would account for the major proportion of SOC in the landscape” [Harper 2013: 642], this southwestern Australia study took deep soil samples in 38 spots across 5 locations. They found that 79% of total carbon store to bedrock occurred below a half meter depth, and 41% occurred below 5 meters depth. “There are two possible sources for the deep carbon; that produced in situ by roots or dissolved carbon that has moved downward from nearer the surface” [Harper 2013: 645]. The sample sites were recently reforested or were under agriculture, and previously had been “covered in a range of xerophytic plants, with root systems that extended to depths of 40 m, such as reported for a Eucalyptus marginata forest” [Harper 2013: 642]. The authors note that more research is needed to understand how deep SOC is affected by land-used changes and climate change.
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths, it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm−2 or 21 % of the total store to bedrock. If this is adjusted to 0.3 m depth, using an exponential function based on the samples in the surface metre, the value decreases to 5.6 kgCm−2. For the individual sites this ranged from 3.6 to 8.0 kgCm−2, or 14 –37 % of the total store. …the surface 5 m contained 16.3±1.38 kgCm−2 or 59 % of the total store to bedrock, with this proportion varying from 47 to 77% across the five sampling locations. The amount of carbon stored in the soils can also be contrasted with the biomass carbon storage of 11.0–16.0kg Cm−2 expected at equilibrium following reforestation for these sites (Harper et al. 2007) and likely previously removed from the sites by deforestation in advance of agriculture [p.645].
Liebig 2008. A study of switchgrass for bioenergy found rates of SOC (Soil Organic Carbon) increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft). “In this study, switchgrass significantly affected change in SOC. . . Across sites, SOC increased significantly at 0–30 cm (1 ft) and 0–120 cm (4 ft), with accrual rates of 1.1 and 2.9 tC/ha (0.44 and 1.16 tC/ac), respectively.” [Liebig 2008:215] This indicates the chronic underestimation of soil carbon capacity in the many studies which by convention only measure SOC down to 30 cm (1 ft).
Machmuller 2015. On intensively grazed former row-crop agricultural land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions. The study “sought to determine how fast and how much soil C accumulates following conversion of row crop agriculture to management-intensive grazed pastures in the southeastern United States. . . . The highest rates of belowground C accumulation occur when land is converted to grassland ecosystems” [Machmuller 2015: 2]. These intensively grazed managed systems led to an approximately 75% increase in soil carbon within six years,
[a] high C accumulation rate [that] stems from year round intensive forage/grazing management techniques on sandy soils with an initially low soil C content due to past conventional-till row crop agriculture. . . . These forage-management techniques are precisely those suggested to increase SOM in pasture systems and when they are applied to soils with degraded SOC content, such as soils in the southeastern United States, rapid C accumulation ensues. . . .
On the basis of a whole farm C-cycle analysis, C accumulation appears to offset methane emissions during the rapid soil C accumulation phase . . . As the C accumulation rate declines these farms will become net C-emitting—similar to all dairy production—because of ruminant methane emissions. However, the substantial soil-quality benefits of higher organic matter remain and will likely increase the sustainability of dairy production using management-intensive grazing [Machmuller 2015:3].
The eventual methane emissions may be markedly less than suggested, however, since the report did not consider methane breakdown from methanotrophic bacteria and atmospheric hydroxyl radical oxidation.
The authors conclude
that pasture-based intensively grazed dairy systems may provide a near-term solution for agricultural lands that have experienced soil-C loss from previous management practices. Emerging land uses, such as management-intensive grazing, offer profitable and sustainable solutions to our needs for pairing food production with soil restoration and C sequestration. [Machmuller 2015: 2-3]
McCosker 2000. A discussion of the introduction of what the author calls “cell grazing,” framed as a paradigm shift in Thomas Kuhn’s terms [Kuhn 1962] over the years 1990-1999. McCosker reviews the dichotomy between researchers and producers, and travels to see actual results in the U.S., Zimbabwe, Namibia, and South Africa. He states, “Only after seeing the outcomes time and again in all possible environments was I finally convinced that the principles could not be faulted.” This kind of paradigm shift has been experienced repeatedly as practitioners must develop the courage to challenge prevailing assumptions in order to take the requisite transitional steps.
Oppenheimer 2015. Farmers Paul and Elizabeth Kaiser own eight acres in Sebastopol in Northern California and they farm three of them, developing a model that other farmers are beginning to use.
[Kaiser] farms a mere eight acres, and harvests fewer than three of them. Nonetheless, his methods are at the forefront of a farming movement that is so new (at least in the U.S.), and so built for a climate-changed world of diminishing rains, that it opens up gargantuan possibilities. One might call this methodology sustainability on steroids, because it can generate substantial profits. Last year, Kaiser’s Sonoma County farm grossed more than $100,000 an acre, which is 10 times the average per-acre income of comparable California farms. This includes Sonoma’s legendary vineyards, which have been overtaking farmland for decades, largely because wine grapes have become much more lucrative these days than food, at least the way most farmers grow it.
Kaiser manages all of this without plowing an inch of his ground, without doing any weeding, and without using any sprays—either chemical or organic. And while most farmers, even on model organic farms, constantly tinker with various fertilizer cocktails, Kaiser concentrates on just one: a pile of rotten food and plants, commonly known as compost, and lots of it. Kaiser then adds this compost to a rare blend of farming practices, both old and new, all aimed at returning dirt to the richest, most fertile seedbed possible.
They use permaculture, agroforestry and other intensive techniques, have built deep, healthy soils by keeping the ground covered and spongy to capture water and carbon, all of which provide solid protection from droughts and floods.
Retallack 2001, 2013. “Grassland expansion initiated increased organic C storage in soils, soil water retention, speed of nutrient exploitation, surface albedo, and C burial in sediments eroded from their soils. These changes had many consequences, including long-term global cooling.” [Retallack 2001:422] and “This climatic zone is not only the most widespread, but also the most fertile region of our planet.” [Retallack 2013:78] The paleohistory of grasslands provides the basis for considering the potential of grasslands as huge biological carbon sinks that may be realized again with regenerative land management
Rodale Institute 2014.
Rodale reports that regenerative grazing practiced on a global scale could sequester 71% of annual emissions of 14 Gt C/yr. Combined results from regenerative grazing and agriculture techniques could, if practiced globally, lead to a net reduction of atmospheric carbon dioxide of 1.7 gigatons per year, or 0.85 ppm/year. This represents a potential sequestration of approx. 3t C/ha/yr (1.2t C/ac/yr) on grasslands and croplands. [Rodale Institute 2014:9] This does not include the considerable contributions of non-agricultural lands, nor recent developments in intensive regenerative practices such as permaculture and biochar.
Moreover, Rodale’s side-by-side trial after 30+ years showed that, following the three-year transition period, organic yields match conventional yields, except in drought years, when organic yields surpass conventional yields. Furthermore, energy input and greenhouse gas emissions were lower in organic systems, and profits were higher.
Rowntree 2016. Examining ruminant methane and net carbon sequestration for grassfed beef in a systems context, Rowntree found net sequestration rates of up to 2.11 tons/ha/yr (0.84 tC/ac/yr) for non-irrigated, lightly stocked grazing.
Therefore, including soil carbon sequestration (SCS) potential brings the differences in grazing environments into focus, and significantly changes the outcome. When comparing two well-managed grazing strategies, grass-finished (MOB) and conventional (IRG), each strategy could be an overall carbon sink, but the MOB grazing would only need sequester half as much carbon (1 tC/ha/yr) as IRG grazing (2 tC/ha/yr) for a net zero greenhouse gas footprint. Methane emissions were similar in both grazing environments, but MOB grazing offered significant benefits in increased carbon sequestration. [Rowntree 2016:36]
It is unlikely that such SCS would take place in the absence of a healthy biodiverse ecosystem, one that is supported through MOB grazing. The result is higher net methane emissions under conventional grazing practices. The opposite occurs with properly managed grazing practices, where grasslands as a system actively build more soil carbon for years, leading to a net increase in soil carbon despite enteric methane production by ruminants.[22]
The recent call for improved management of grazing systems as part of an international climate change mitigation strategy is critical, particularly in light of many existing beef LCAs [Life Cycle Assessments] that have concluded that beef cattle produced in grazing systems are a particularly large sources of GHG emissions. To identify the best opportunities to reduce GHG emissions from beef production, a systems approach that considers the potential to increase soil C and reduce ecosystem-level GHG emissions is essential… [W]e generated an LCA that indicates highly-managed grass-finished beef systems in the Upper Midwestern United States can mitigate GHG emissions through SCS while contributing to food provisioning at stocking rates as high as 2.5 Animal Units (AU) per hectare. From this data, we conclude that well-managed grazing and grass-finishing systems in environmentally appropriate settings can positively contribute to reducing the carbon footprint of beef cattle, while lowering overall atmospheric CO2 concentrations. [Rowntree 2016:36]
Schwartz 2013. Cows Save the Planet was a landmark book that helped launch the regenerative agriculture and eco-restoration climate movements. The author gathers stories from practitioners around the world and paints a picture of broad possibilities for addressing global warming, floods, droughts, desertification, malnutrition and many other seemingly unrelated problems that have a single common cause: human mismanagement of lands across the planet. An excellent introduction for a general readership as well as for scientists unfamiliar with the potentials that nature provides.
Shinn n.d. Ridge Shinn is a rancher in Central Massachusetts (Big Picture Beef) who introduced Allan Savory’s methods to the state. For over ten years he has been investigating the holistic context for bringing healthy meat through the local supply chain, and its relationship to human health and global warming. He states,
Since the 1990’s, science has discovered important connections between rotational grazing, soil health, and healthy food. Big Picture Beef’s methods for raising 100% grass-fed cattle offer huge benefits for the environment and for society. The long term goal of the program is Northeast beef for Northeast markets, carbon sequestration, soil fertility and biodiversity, energy savings, and a revitalized rural economy.
Teague et al. 2016. In a review of the literature, the authors conclude that regenerative conservation cropping and adaptive multi-paddock grazing can turn agricultural soils from a carbon source in conventional agriculture into a carbon sink at rate of ~3 tC/ha/yr (~1.2 tC/ac/yr). Key factors include the use of no-till, cover crops, managed grazing, organic soil amendments and biotic fertilizer formulations. These practices can result in elimination of soil erosion and loss, the greatest agricultural contribution to global warming (1 Gt C/yr). Benefits may include “increased water infiltration, improved water catchment, greater biodiversity, increased ecosystem stability and resilience, and improved C sequestration.” [Teague 2016:158]
Conclusion
These reports demonstrate the promising potential for storage of massive amounts of soil carbon to address both the atmospheric and eco-destruction aspects of climate, along with dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Ausmus, Steven, A Surprising Supply of Deep Soil Carbon, Agricultural Research, USDA, February 2014, 4-6, https://agresearchmag.ars.usda.gov/ar/archive/2014/feb/february2014.pdf (s.a. Follett 2012). [Grasslands]
Blakemore, R.J. 2016a, Veni, Vidi, Vermi – I. On the contribution of Darwin’s ‘humble earthworm’ to soil health, pollution-free primary production, organic ‘waste’ management & atmospheric carbon capture for a safe and sustainable global climate, July, 2016, VermEcology Occasional Papers, 2(1): 1-34, https://veop.files.wordpress.com/2016/09/vvv-i.pdf. [Soils, Grasslands]
Blakemore, R.J. 2016b, Veni, Vidi, Vermi – II. Earthworms in organic fields restore SOM & H2O and fix CO2, July, 2016, VermEcology Occasional Papers, 2(2): 1-26, https://veop.files.wordpress.com/2016/09/vvv-ii.pdf. [Soils, Grasslands]
Blakemore, R.J. 2016c, Cosmopolitan Earthworms - An Eco-Taxonomic Guide to the Peregrine Species of the World, 6th ed., VermEcology, Yokohama 2016 https://vermecology.wordpress.com/2016/07/20/cosmopolitan-earthworms-6th-edition-2016-is-now-available/. [Soils]
Byck, Peter, 2014, Soil Carbon Cowboys, https://vimeo.com/80518559. [Grasslands]
Ellison, David, et al., Trees, forests and water: Cool insights for a hot world, Global Environmental Change 43, January 2017, 51–61, http://www.sciencedirect.com/science/article/pii/S0959378017300134. [Grasslands, Forests, Water]
Follett, Ronald F., Kenneth P. Vogel et al. 2012, Soil Carbon Sequestration by Switchgrass and No-Till Maize Grown for Bioenergy, Bioenerg. Res. May 4, 2012, 5:866–875, http://link.springer.com/article/10.1007/s12155-012-9198-y (s.a. Ausmus 2014). [Grasslands, Croplands]
Harper R.J., M. Tibbett 2013, The hidden organic carbon in deep mineral soils, Plant and Soil 368 (1-2): 641-648, http://link.springer.com/article/10.1007%2Fs11104-013-1600-9. [Grasslands]
Héry, Marina, Andrew C Singer, Deepak Kumaresan, et al. 2008, Effect of earthworms on the community structure of active methanotrophic bacteria in a landfill cover soil, International Society for Microbial Ecology 2, 92–104, https://www.ncbi.nlm.nih.gov/pubmed/18049457.
Kernecker, Maria, Joann K. Whalen, Robert L. Bradley 2014, Endogeic earthworms lower net methane production in saturated riparian soils, Biology and Fertility of Soils 50 (7), https://link.springer.com/article/10.1007/s00374-014-0965-0. [Grasslands]
Kuhn, Thomas 1962, The Structure of Scientific Revolutions, U. Chicago Press, 1962; full text available (2d edition, 1970) at http://projektintegracija.pravo.hr/_download/repository/Kuhn_Structure_of_Scientific_Revolutions.pdf. [Introduction]
Liebig, M.A., et al., Soil Carbon Storage by Switchgrass Grown for Bioenergy, Bioenerg. Res. (2008) 1:215–222, http://naldc.nal.usda.gov/download/28132/PDF [Grasslands, Croplands]
Machmuller, Megan B., et al., 2015, Emerging land use practices rapidly increase soil organic matter, Nature Communications, April 30, 2015, p. 1-5, http://www.nature.com/articles/ncomms7995. [Grasslands]
NASA Earth Observatory 2011, The Carbon Cycle, http://earthobservatory.nasa.gov/Features/CarbonCycle/. [Soils, Grasslands]
Retallack, Gregory J. 2001, Cenozoic Expansion of Grasslands and Climatic Cooling, The Journal of Geology, 2001, 109 (407–426), http://www.journals.uchicago.edu/doi/abs/10.1086/320791?journalCode=jg. [Grasslands]
Retallack, Gregory J., 2013, Global Cooling by Grassland Soils of the Geological Past and Near Future, Annu. Rev. Earth Planet. Sci. 41:69–86, http://www.annualreviews.org/doi/abs/10.1146/annurev-earth-050212-124001. [Grasslands]
Rodale Institute White Paper 2014, Regenerative Organic Agriculture and Climate Change: A Down-to-Earth Solution to Global Warming, https://rodaleinstitute.org/regenerative-organic-agriculture-and-climate-change/ [Soils, Grasslands]
Rowntree, Jason, et al., Potential mitigation of midwest grass-finished beef production emissions with soil carbon sequestration in the United States of America, Future of Food: Journal on Food, Agriculture and Society 4 (3) Winter 2016, http://futureoffoodjournal.org/index.php/journal/article/view/232 [Grasslands]
Savory Institute 2016, Restoring the Climate through Capture and Storage of Soil Carbon through Holistic Planned Grazing, http://savory.global/assets/docs/evidence-papers/RestoringClimateWhitePaper2015.pdf [Grasslands]
Savory Institute, An Exploration of Methane and Properly Managed Livestock through
Holistic Management, 2015, http://savory.global/assets/docs/evidence-papers/exploration-of-methane.pdf [Grasslands]
Savory, Allan, How to fight desertification and reverse climate change, TED Talk, February 2013, https://www.ted.com/talks/allan_savory_how_to_green_the_world_s_deserts_and_reverse_climate_change [Grasslands]
Suttie, J.M., S.G. Reynolds, C Batello, eds. 2005, Grasslands of the World. Food and Agriculture Organization of the United Nations, Ch. 1, http://www.fao.org/docrep/008/y8344e/y8344e05.htm - bm05. [Grasslands]
Teague, W.R., Steve Apfelbaum, Rattan Lal et al. 2016, The role of ruminants in reducing agriculture’s carbon footprint in North America, J. Soil and Water Conservation, March/April 2016, 71:2, http://www.jswconline.org/content/71/2/156.abstract. [Grasslands, Croplands]
White, R., S. Murray, M. Rohweder 2000, Pilot analysis of global ecosystems: Grassland ecosystems, World Resources Institute, Washington D.C., http://www.wri.org/publication/pilot-analysis-global-ecosystems-grassland-ecosystems.
[Croplands, Grasslands]