
Soils
This compendium is, if nothing else, a testament to the key role soils must play if we are to preserve life on earth through the anthropocene. Soils, the engine of every terrestrial ecosystem, are themselves wildly diverse subterranean ecosystems providing habitat to countless trillions of micro- and macro-organisms. These organisms themselves create the soil and its functionality by ingesting dead organic matter, thereby breaking it down so that nutrients become available to plants which in turn feed everything else up the food chain. In the everyday processes of foraging, reproducing, exchanging, growing, and dying, the biology upon and within the soil regulates the climate by consuming, transforming, and burying molecules of carbon. This buried and embodied carbon would otherwise be atmospheric carbon dioxide. Thus, it’s not the soil as an apparently homogenous substrate that concerns us. It is the living, biodiverse soil ecosystem that matters for maintaining a global climate system hospitable to human life. In this section, we discuss the contents, processes and functions of the soil and the threats to it, with an eye to highlighting the absolutely critical role of soil biology.
Overview
A noteworthy perspective is that a teaspoon of healthy soil holds more microorganisms than there are people on earth. The biodiversity in that teaspoon rivals that of the Amazon rainforest. In fact, it’s these very microorganisms (along with their larger counterparts, like earthworms and mammals) that themselves create the content and structure of soil by breaking down organic material, secreting sticky biomolecules, and burrowing, while also depending on the soil for habitat and food. Because of the interrelatedness of diverse and vital roles played by soil organisms (roles such as fixing nitrogen, suppressing disease, creating channels for water and root penetration, and forming soil aggregates, for example), the soil as a whole could be viewed as a macro-organism, made up of myriad “organs” working together and becoming sick or healthy as if it were a living body (within a field, for instance) [Magdoff 2009].
Furthermore, the particular community composition of microorganisms in a given soil can affect the overall functioning of the soil. It is generally understood that high concentrations of fungi are associated with healthy soil, while soils regularly disturbed by tillage tend to be bacteria-dominant [Magdoff 2009]. Fungal populations are also associated with increased carbon sequestration.
Kallenbach et al [2016] demonstrate that distinct microbial communities, especially those with high fungal concentrations, are a better predictor than clay mineral content of Soil Organic Carbon (SOC) production. Morrien et al. [2016: 1] show “that during nature restoration the efficiency of nutrient cycling and carbon uptake can increase by a shift in fungal composition and/or fungal activity.” Specifically, the proportion of arbuscular mycorrhizal fungi[5] increases over time leading to greater carbon uptake by fungi, “without an increase in fungal biomass or shift in bacterial-to-fungal ratio.” Recent research by David Johnson of New Mexico State University shows that high fungal to bacterial ratios are correlated with strikingly high carbon sequestration and high plant productivity [Johnson n.d.; Johnson 2017] (see also Croplands section).
Due to the exceptional scope and complexity of its biological activity, soil is a major player in the self-regulating system of Earth’s environment by taking up and releasing large quantities of carbon dioxide, oxygen, methane and other gases [Brady 2008]. Indeed, it is the living organisms on and in the soil that breathe these gases in and out. Under favorable conditions balance is maintained due in part to soil organisms holding carbon in place, both within their own biomass and through ongoing carbon-rich soil creation.
Soils store more carbon than is found in the atmosphere and terrestrial vegetation combined [FAO 2017a]. The capacity for soils to store carbon depends on various factors, such as temperature, moisture level, soil type, and topography. Carbon-rich peatland and organic soil[6], which occupy 3% of land but contain an estimated 30% of the world’s soil carbon, are commonly located in cold environments or rainy, humid environments, where productivity is high and decomposition slow. Another third of the world’s soil carbon is in drylands (areas where potential evaporation/transpiration exceeds average rainfall) which cover approximately 40% of Earth’s land area, but tend toward lower productivity, slower carbon accumulation, and susceptibility to erosion when mismanaged [FAO 2017b].
Ecological processes have maintained a balanced carbon cycle over time, keeping the Earth at a relatively stable temperature for hundreds of thousand of years. Although there have been temperature fluctuations before, it is virtually certain that human activity (mainly burning fossil fuels, desertifying and clearing land) is upsetting the carbon cycle, causing atmospheric concentrations of heat-trapping carbon dioxide to have risen from about 280 parts per million (ppm) prior to the 19th century to more than 400 ppm today [NASA 2017]. This, in turn, is changing the temperature at a faster rate than at any time in at least 1,000 years [Smith 2015]. Since the start of industrial era, carbon emissions from the soil due to deforestation and land conversion is estimated to be 136 Gt (Gigatons, or one billion tons), a little less than half the amount of carbon emissions from fossil fuel combustion over the same time period [Lal 2016]. Estimates of carbon lost to the atmosphere from land use since the beginning of agriculture range as high as 537 Gt [Buringh 1984: 91].
It is important to note that soil that has been greatly depleted of carbon can become a carbon sink when managed according to ecological/restorative principles. A carbon sink is anything that absorbs more carbon than it releases as CO2. “Soils that are depleted of SOC have the greatest potential to gain carbon, but also the least propensity to do so.” [FAO 2017b: 7] This is because, while depleted soils can theoretically regain the entire massive amount of carbon they’ve lost, they first need to be biologically re-activated through restorative management practices. It is vital to note that degraded lands (and all lands) have this capacity for renewal as the global community seeks both to reduce CO2 emissions and to draw down excess atmospheric CO2 as quickly as possible. Conversely, organic soils, forests, and other intact ecosystems with large existing stores of carbon in the soil and vegetation have the potential to become new sources of emissions, and must therefore be properly managed and preserved.
According to the Rodale Institute, the fastest, least expensive and most efficient way to rebuild agricultural soils is through “regenerative organic agriculture,” designed to bring carbon and biodiversity back into the soils.
Recent data from farming systems and pasture trials around the globe show that we could sequester more than 100% of current annual CO2 emissions with a switch to widely available and inexpensive organic management practices, which we term “regenerative organic agriculture.” These practices work to maximize carbon fixation while minimizing the loss of that carbon once returned to the soil, reversing the greenhouse effect. [Rodale Institute 2014: 2] (See more in Croplands section.)
Management practices for building carbon in soils involve increasing input and reducing losses of soil organic matter (SOM). SOM is made up of fungi, bacteria, countless microorganisms, decaying plant matter, decomposing animals, and products formed from their decomposition. SOM also contains humus, an organic material believed to contain stable forms of organic carbon. However, the inherent resistance of humus and other compounds to decomposition and release of carbon dioxide into the atmosphere has recently come into question, as the stability of soil compounds may be an ecosystem property and not an intrinsic property of the compound itself [Schmidt 2011]. Thus the understanding of soils is transformed from a consideration of properties of isolated variables to properties of the system as a whole, with its exceedingly complex interactions, a transformation considered throughout this Compendium.
In sum, SOM provides food for soil organisms and plants, enhances microbial biodiversity, creates pore space, increases cation-exchange capacity (CEC)[7], and increases buffering capacity (ability to resist change in acidity or pH). All of these factors affect the soil’s ability to hold carbon [Fenton 2008].
Soil organic carbon (SOC) levels are directly related to the amount of SOM. The key factors in SOC levels are photosynthesis, respiration and decomposition. Photosynthesis is the process used by plants to harness energy from sunlight, combined with the CO2 they take from the air and water from the ground, and convert it into energy storage (in the form of sugars) for their own growth and survival. Respiration is the measure of CO2 released from soil microbes and plant roots into the surrounding soil.
Soil C results both directly from growth and death of plant roots and above-ground biomass, as well as indirectly from transfer of carbon-rich compounds from roots to soil microbes. The roots of almost all plants form symbiotic associations with arbuscular mycorrhizal fungi – the roots provide the fungi with energy in the form of carbon while the fungi provide the plant with nutrients. While decomposition of biomass by soil microbes results in carbon loss as CO2 from the soil due to respiration, a proportion of the original carbon is retained in the soil through the formation of numerous stable soil carbon molecules. Carbon is also stored in soil aggregates, which are formed in collaboration with microorganisms.
The multiple soil organisms providing vital ecosystem functions are sensitive to various practices and changing conditions, even those occurring above the soil surface. Applying synthetic nitrogen fertilizer, for example, can affect soil pH, which in turn can negatively affect soil organisms. Organic amendments such as manure and compost, on the other hand, provide direct carbon sources for soil organisms and an indirect carbon source via increased plant growth without negative side effects.
Temperature also affects soil microbes. For example, the scottnema lindsayae nematode cannot survive in its native habitat in Antarctica if temperatures become too warm or too cold, and research suggests that a 65% decline in activity of these nematodes over 12 years could lead to a 30% reduction of carbon cycling in soils [Wall 2014]. Similarly, earthworms, described by Darwin [1881] as “nature’s plow,” are damaged both by cultivation and by the use of toxic herbicides and other agrichemicals often used in “conventional” no-till.
A healthy biosphere has the power to draw down carbon and store water and through this support biodiverse life forms, including humans. Most importantly, in partnership with nature, humanity can restore ecosystems, including agro-ecosystems[8], to create a functional global biosphere once again. This begins with the soil, and requires at once protecting intact organic soils and their invaluable concentrations of stored carbon, and restoring carbon-depleted mineral soils so as to activate their capacity as massive carbon sinks. Remarkably, taking action to protect and rebuild soil is almost universally available, low-cost, safe, and has the power to mitigate and reverse climate change in a relatively short period of time.
Eco-restoration has numerous co-benefits such as restoring abundant food production, eliminating floods and droughts, restoring water supplies, building strong local economies and providing shade, beauty, and recreation.
Soil Article Summaries
Crowther et al. 2016. When this paper first appeared in Nature, it seemed to raise serious objections to the hypothesis that soils as carbon sinks could have a significant impact on climate. On further examination, however, it may be more about the perils of isolating variables than about the limitations of soils.
There is growing confidence that warming generally enhances fluxes to and from the soil, but the net global balance between these responses remains uncertain [Crowther 2016: 104].
The authors analyze data on the soil’s response to warming from 49 experimental sites in North America, Europe and Asia, across six biomes. They find that the level of carbon loss from the soil is contingent upon the size of the soil’s original carbon stock, and on the duration and extent of the warming. Extrapolating to a global scale, they estimate that an additional 1oC temperature rise will release anywhere from 30 ± 30 to 203 ± 161 of carbon Gt from the soil.
The authors themselves highlight several limitations in their analysis due to lack of data, including from experimental sites in the tropics, from soil at depths greater than 10cm, and on the effects from biotic responses to warming. It is noteworthy that, as the authors state:
Our current understanding of global feedbacks is dominated by the physical sciences, but changes in the physiology and community compositions of organisms have been shown to have strong effects on the strength of this feedback [107].
In other words, for a more precise prediction of how global warming will affect the soil’s net release of carbon, we need to understand better how plants, animals and microbes may interact and respond to that same warming with respect to their effect on the land-carbon climate feedback.
Like Amundson et al., these authors make projections about the soil’s response to climate change without consideration of the wide spectrum of possible land management practices, ranging from clearcutting and urbanization to reforestation and Holistic Management. Presumably, a more accurate picture of the soil’s response to future warming must incorporate measurements of the soil’s resilience capability, based on its level of exposure to oxidative or erosive elements vs. embeddedness in intact ecosystems.
Grindrod 2017. A brief review for non-scientists of soil microbiology and the growing understanding that soil health is, in many ways, all about microbes and their complex interactions with mineral, plants and animals. It likely that micro-organisms are the key to soil fertility as a result of the universe of microbial chemistry that affects, among many things, availability of nutrients and plant health, rendering other forms of dangerous inputs such as biocides unnecessary. See also David Johnson [2017].
Hart 2015. In a year of severe heat and drought, cover crops and no-till proved their value for North Carolina Farmer Russell Hedrick, reports Southeast Farm Press. In a county where average SOM is 1.5%, Hedrick’s farm has 4.8% SOM after just four years of no-till and cover crops, which have increased the water infiltration in the soil while also cooling the soil surface as much as 20 degrees F. Hedrick also introduced cattle into his system to graze on the cover crops, a strategy his NRCS[9] advisor told him he “couldn’t afford not to do,” due to the ruminants’ stimulating effects on the life of the soil. During this drought year, while Hedrick’s neighbors lost their crop, he was still able to harvest 110 bushels per acre of corn although it is twice that much in a normal year. Furthermore, Hedrick controls weeds with less herbicide due to the thick mat and allelopathic[10] effects of the five-plant cover crop mix, and he no longer uses any insecticides. He’s also started to see earthworms on his land for the first time, helping to break down organic matter and carry carbon deeper into the soil.
Kallenbach 2016.
Soil organic matter (SOM) and the carbon and nutrients therein drive fundamental submicron- to global-scale biogeochemical processes and influence carbon-climate feedbacks. Consensus is emerging that microbial materials are an important constituent of stable SOM, and new conceptual and quantitative SOM models are rapidly incorporating this view. However, direct evidence demonstrating that microbial residues account for the chemistry, stability and abundance of SOM is still lacking. Further, emerging models emphasize the stabilization of microbial-derived SOM by abiotic mechanisms, while the effects of microbial physiology on microbial residue production remain unclear. Here we provide the first direct evidence that soil microbes produce chemically diverse, stable SOM. We show that SOM accumulation is driven by distinct microbial communities more so than clay mineralogy, where microbial- derived SOM accumulation is greatest in soils with higher fungal abundances and more efficient microbial biomass production. [Kallenbach 2016: Abstract]
Schmidt 2011. Complex interactions, not intrinsic chemical properties, may determine the persistence of soil organic carbon molecules and their climate effects:
Most soil carbon derives from below-ground inputs and is transformed, through oxidation by microorganisms, into the substances found in the soil. By moving on from the concept of recalcitrance and making better use of the breadth of relevant research, the emerging conceptual model of soil organic carbon cycling will help to unravel the mysteries surrounding the fate of plant- and fire-derived inputs and how their dynamics vary between sites and soil depths, and to understand feedbacks to climate change. We argue that the persistence of organic matter in soil is largely due to complex interactions between organic matter and its environment, such as the interdependence of compound chemistry, reactive mineral surfaces, climate, water availability, soil acidity, soil redox state and the presence of potential degraders in the immediate microenvironment. This does not mean that compound chemistry is not important for decomposition rates, just that its influence depends on environmental factors. Rather than describing organic matter by decay rate, pool, stability or level of ‘recalcitrance’ – as if these were properties of the compounds themselves – organic matter should be described by quantifiable environmental characteristics governing stabilization, such as solubility, molecular size and functionalization. . . . Because many, if not most, organic molecules in soils are of microbial origin, experiments are needed that identify the long-term drivers of microbial-cell and microbial-product decomposition, rather than focusing on the immediate fate of fresh plant material. (emphasis added) [Schmidt 2011: 52-3]
This study opens many possibilities for new approaches to soil carbon, including assessment from a systemic as opposed to a reductionist perspective:
More generally, though, the major advances in our understanding of soils will come from research grounded in the theory of many disciplines and in the practice of many approaches. The future research agenda for soils will integrate many different fields and have broader goals than it might have had in the past, with longer time horizons, wider spatial coverage, and an imperative to connect carbon, water and nutrient cycles, so as to understand the soil-plant system as a crucial part of our biosphere. [Schmidt 2011: 55]
Do We Have More Soil for Carbon Storage than We Thought?
This is a discussion of a hypothesis by oligochaetologist[11] Rob Blakemore, Ph.D. He suggests that accounting for varying planetary topography may dramatically increase the projections for soil-carbon storage potential. This has significant implications for the importance of eco-restoration efforts to address climate, and the speed with which we can draw carbon from the atmosphere.
We believe that this hypothesis is both reasonable and logical, but support is largely inferential at the current time. It is our hope that this discussion will stimulate further research into the potential of photosynthetic carbon capture and sequestration in soils, and that this development will motivate an intensification of eco-restoration efforts.
Introduction
Area measurements of the earth’s terrestrial surface are conveniently two-dimensional (2-D), yet the true surface has a certain amount of three-dimensional undulation (3-D). Therefore the actual surface and volume of soils on Earth may be underestimated by an unknown factor using current measurements and models.
This leads to the proposition that global soils may have the capacity to store greater quantities of atmospheric carbon than previously assumed. The role of eco-restoration, with soils as the primary storage medium, with above-ground and below-ground biodiversity as the process for incorporating carbon into soils, is therefore a key to any successful climate strategy.
Why “Flat-Earth” soil estimates may be incorrect
The surface of the earth is conventionally calculated based on an assumption of two-dimensional areas within a set of coordinates. Yet, despite decades of satellite imagery, accurate information on the actual surface area of the earth is surprisingly elusive: the land has undulating terrain adding to its extent. The importance of this distinction is that the total volume of global soils, as well as primary productivity, i.e., photosynthesis, relate to actual soil surface area, not to a two-dimensional model projection thereof.
Actual three-dimensional surface area is most important for calculations of our total topsoil resource or, as Darwin [1881: 49] describes in his ground-breaking book on earthworms: “The vegetable mould . . . covers, as with a mantle, the surface of the land . . .”
It is not unreasonable to estimate a flat projection of photosynthesizing terrestrial surface area of 12 billion hectares (gigahectares or Gha).[12] Some multiple will account for surface irregularities that increase that surface area, and for the sake of discussion we will use a conservative factor of 2.
A practical example of an applicable mathematical estimate of irregular surface areas is from a paint manufacturer [Resene, n.d.], who estimates that compared to a flat surface, a 200 m2 corrugated sheet has 10.5% larger surface area (= 221 m2), and that Anaglypta or Stucco textures, i.e., bumpy like Earth, have surface area 40-100% greater than that of the base area.
One can also use geometry and knowledge of fractals in order to estimate a reasonable multiplier of the available flat topography for illustrative purposes. As a paradoxical (i.e., counter-intuitive) fractal, the actual true land surface area may be infinitely expanded at increasingly finer scales of observation as in a 3-D version of the 2-D “Coastline Paradox” that, in practice, increased the linear distance estimate of Britain’s coastal outline more than six fold[13].
Therefore, this revised estimate of a true land surface takes into account the area exposed to the Sun’s irradiation, and includes all topsoil that supports the plants upon which we depend. If that surface area is doubled by irregularities in the surface of the land, so proportionately is capture of the Sun’s energy and resultant soil activity, including carbon and water storage.
These are very rough approximations: we can say with certainty only that current “Flat-Earth” surface area, and therefore soil volume calculations, are under-representations, and likely significant ones. We look forward to more detailed studies from researchers with the resources to pursue them.
Leaf Area Index
The primary productivity providing for most life on Earth operates at the biological scale of a leaf.[14] Average leaf sizes reportedly range from 0.011 to about 39.5 cm2 but no data are readily available for the topographical surface area and volume of underlying topsoil that supports these plants. An alternative estimate of effective terrestrial surface area is possible if we apply a Leaf-Area-Index (LAI).
LAI is a dimensionless quantity that characterizes plant canopies defined as the one-sided green leaf area over the flat unit ground surface area (LAI = leaf area / ground area). In other words, LAI is a factor that derives the effective ground area for which the plant is productive based on how much photosynthesis is actually turning atmospheric carbon dioxide molecules into above- and below-ground biomass. For example, if the surface area on the ground under a tree occupies 10 m2 and the total leaf surface area is 45 m2, the tree is accomplishing 45 m2 worth of photosynthesis which would be underestimated by only accounting for the tree’s ground-level two-dimensional footprint, as is conventionally reported.
LAI’s range from 0 (bare ground) to ~18 (dense forests) and a global average is 4.5. The authors of this source state that “LAI is a key variable for regional and global models of biosphere-atmosphere exchanges of energy, carbon dioxide, water vapour, and other materials.” [Asner et al. 2003: 195]
For our purposes, we apply LAI to the recalculated undulating and rough-surface topography. If we therefore take our hypothetical but reasonable estimate of a flat 12 Gha of photosynthesizing land and multiply it by 2, we have 24 Gha of non-flat photosynthesizing land surface area. If we include the LAI multiplier of 4.5 to those 24 Gha, we arrive at the equivalent of 108 Gha of photosynthesis, or 9 times more carbohydrate production by green plants than would be estimated from flat-surface-area measurements.
Why does it matter?
We are rapidly losing soils, with global topsoil erosion rates reportedly greater than 2,000 tonnes per second[15] [Pimentel 2013: 447]. Soil is further depleted by agri-chemical pollution and urbanization [Blakemore 2017a], that is, land degradation is due in no small measure to loss of natural soil fertility and excess synthetic nitrogen [Rockström 2009: 472, Fig. 1]. Therefore it is clearly in our best interests, and in the interests of the remaining living organisms on planet Earth, to get accurate information about the 3-D topography of the land, which will alter calculations about surface area and volumes of soils. This in turn will increase the potential for sequestration of carbon and for water storage in soils, vital knowledge that could allow more rapid and effective restoration efforts.
How Much Soil Is There on Earth?
In addition to terrain considerations, we now consider Soil Organic Carbon (SOC) weight in Gigatons, as opposed to previously considered surface area in Gigahectares. Blakemore [2016a: 11] noted that:
Soil carbon values require allowance for intractable glomalin adding a further 5-27% to almost all SOC tallies (Comis, 2002). Plus data from deep soils may increase budgets: e.g., Harper & Tibbett (2013) found C up to five times greater in Australian soils at depth >1 m and down to 35 m in some cases. The Walkley-Black method itself underestimates total C by about 20% with a correction factor of ca. 1.3 often required, whereas latest techniques using mid-infrared (MIR) spectroscopy give more accurate readings. These three factors combined would surely increase SOC totals.
Glomalin, only discovered in 1996, is a stable fungal molecule tightly-bound to soil particles as a major component of soil organic matter. Glomalin adds up to 27% of total SOC [Khursheed 2016], and is stable for 7 to 42 years, depending on conditions [Comis 2002: 4].
Thus an answer to “How much soil is there on Earth?” is still elusive. How much soil can be built on Earth through the activity of healthy biological systems and how quickly these transformations can take place remains an intriguing question. Allowing for glomalin, deep soil data and carbon in living or dead roots [Jackson 1997: T1], soil carbon quantities are likely considerably higher than conventional estimates, as is the potential for future carbon sequestration in soils.
Conclusion
More than two millennia ago Aristotle told us the Earth was not flat and he also concurred with Plato in recognizing that soil erosion and loss of humus and earthworms is catastrophic to civilization [Montgomery 2008: 51]. Leonardo da Vinci’s observation 500 years ago that “We know more about the movement of celestial bodies than about the soil underfoot” seemingly still rings true. An essential feasible and achievable solution is to apply what Sir Albert Howard termed Nature’s Law of Return, i.e., to vermi-compost all organic ‘wastes” to restock the topsoil [Howard 1945].
Most crucially vital, we must determine the amount of living topsoil remaining and its potential restoration through organic regenerative land management worldwide applying principles and practices of Permaculture (Mollison 1988; Blakemore 2017). The challenge now is for professional geographers, astronomers and others to provide actual topographic values for land and topsoil contribution to global photosynthesis and the carbon cycle.
Earthworms
Although often overlooked, ignored or taken for granted, earthworms are nevertheless keystone soil species, mediators and moderators for rebuilding healthy, biodiverse, high carbon and moisture rich topsoil [Darwin 1881; Blakemore 2016c]. We depend on soils for more than 99% of our food and 100% of our timber and natural fibres [Blakemore 2012, Pimentel 2013]. As an integral part of organic production, earthworms are key to agricultural sustainability and global ecosystem stability. Ancient in origin (probably pre-Cambrian but certainly more than 500 million years old), the 7,000 known species of earthworms are ubiquitous and invariably associated with topsoil humus. Earthworms are a basis of terrestrial food webs and the ultimate detritivor [Blakemore 2016c], recently reinstated as key players in the International “4 per 1000 Initiative” [4p1000.org, n.d.] to increase soil organic matter to store carbon. In this section, we discuss the abundance and variety of earthworms and their role in soil health and functionality.
Overview
Extrapolating data from Darwin [1881], their population numbers around 1.3 x 1015 or 1.3 quadrillion globally with biomass of 0.4 t/ha x 9.5 Gha of productive land = 3.8 Gt. This is about ten times the biomass of all humanity, and twice that of both all domesticated stock and total global fish [Blakemore, 2017]. Forming possibly the largest beneficial animal resource on the planet, earthworms are yet apparently severely depleted by cultivation and agrichemical excesses of industrial farming, often being absent from such soils [e.g. Lee 1985] with both their populations and biodiversity in decline [Blakemore 2016a, b, c].
In comparison to intensive agrichemical farming, studies by Blakemore, [2000, 2016a, b] show a diverse array of up to 23 earthworm species per organic farm site (mean 13 spp), implicated in 16-80% increased crop or pasture yield (mean +39%) plus an average of 12% extra soil moisture storage (range 7-91%) compared to conventional neighbour farms. Carbon sequestration is restored at rates two to three times higher in pasture. Such findings are highly relevant due to looming species extinction and climate change with requirement to meet the needs of a growing population. Organic farming can thus produce higher yields and sequester more carbon.
Earthworms may number up to 1,000~2,000/m2 (10-20 million/ha, or 4-8 million/ac) in fertile soils with biomass as high as 3-5 t/ha, (1.2-2 t/ac ) so earthworm stocks may outweigh the above ground stock [Lee 1985; Blakemore 2016c, 2017]. Earthworm abundance and diversity increase in a truly sustainable system as they convert all organic ‘wastes’ into humus-rich compost while processing all atmospheric CO2 in 12 yr cycles [Blakemore 2016a]. Their burrows, as long as 9,000 km/ha (2250 mi/ac) [Kretzchmar 1982] and up to 15 m in depth (49.2 ft) [Sims & Gerard, 1999: 27, as cited in Blakemore 2016c] aerate, improve water infiltration and, importantly, provide habitats for many other beneficial organisms and microbes that they help distribute throughout the entire soil profile. All rainfall is filtered through their burrows and water is stored in worm-worked humus. Blakemore [2000] found up to 90% extra water in pasture compared to adjacent arable fields, and organic arable soil stored 40% more water than chemically farmed arable soil [Blakemore, Hochkirch 2017].
Wormless soils are economically and ecologically expensive: they need to be plowed regularly, and require extra irrigation plus subsidized artificial chemical nitrogen fertilizers and biocide sprays to fight off plant infections and infestations [Howard 1945; Balfour 1975]. This toxic burden has severe impact upon non-target organisms and any organism fed the crops – including humans – as well as poisoning the soil, air, waterways and oceans. Such findings are summarized in Lady Eve Balfour’s IFOAM presentation in 1977 [Balfour 1977]. Another compelling reason for earthworm conservation is that it is impossible to “geoengineer” by addressing isolated variables the many benefits and essential irreducible systems services that earthworms freely and relentlessly provide. In other words, we have no viable alternative to earthworms.
Soil and Earthworm Relationships
We face a complexity of inter-relating ecological problems. Intensive chemical agriculture is a major GHG contributor (28-50%) and a major source of extraneous CO2 (currently 10-25% and in total historically up to 40%) [Houghton 2010: 338, 348]:
Globally, the conversion of lands to croplands has been responsible for the largest emissions of carbon from land-use change. . . From 1850 to 2000, land use and land-use change released an estimated 108–188 Gt (billion tons) of carbon to the atmosphere, or about 28–40% of total anthropogenic emissions of carbon (274 Gt C from fossil fuels) [Strassmann 2008].
The FAO [Gerber 2013] found that intensive industrial livestock farming (rather than organic husbandry) contributed 14.5% of human-induced GHG emissions. A newspaper report [Bryce 2013] comments:
The FAO’s last livestock report, a 2006 assessment titled Livestock’s Long Shadow, found that farms breeding chickens, pigs, and cows for meat and dairy products, produced a disconcerting 18% of global greenhouse gas emissions . . . Around 30% of global biodiversity loss can be attributed to livestock production, such as the spread of pasture land or turning over forests and savannahs.
Although these figures vary due to different formulas for budgeting, it’s clear that agriculture in all its forms, including the practice of forest clearance, is a major contributor to GHG emissions.
The traditional, innovative & scientific methods of non-chemical, organic farming and Permaculture appreciate the importance of earthworm conservation [Howard 1945; Balfour 1975; Mollison 1988]. As a key player in natural processes and crucial issues, Darwin’s “lowly earthworm”, although neglected, warrants re-ascendency to its former position as premier farm livestock [Howard 1945]. For our own health and for that of our planet, we urgently need wholly natural vermi-composting at all scales (from kitchen to continent) in order to replace synthetic fertilizers and to facilitate rapid transition to broad-acre organics that also has earthworm livestock at its core. Enabling earthworms to restore healthy soils is vital to stabilizing climate. All organic ‘wastes’ and manures should be recycled via vermi-composting and appropriate management employed to enhance populations of field-working worms.
Earthworm Article Summaries
van Groenigen et al. 2014. In a recent meta-analysis, while not considering organic farming or carbon per se, this study confirmed earthworm presence corresponding to crop yield increases of 25%, which is comparable to average ~39% extra organic yield in soils with earthworm proliferations determined by Blakemore [2000, 2016b]. This supports earlier studies by Wollny [1890: Forschungen auf der Gebiet der Agrikultur-Physik, 13, s. 381] that found addition of earthworms to soil led to a marked increase of cereal grain by 35-50% and of straw by 40%.
Solomon 2013.
Although earthworms are beneficial in gardens and agricultural fields, they are harmful to Michigan’s forests where they are an invasive species. . . . Earthworms are not native to Michigan and the Great Lakes region, at least not since before glaciers covered the region; they were brought here during European settlement in the 1800s or possibly earlier. Plants, wildlife and forests evolved without any of these creatures around. They are now an invasive species that harms forests.
Hardwood forests without earthworms have a thick layer of slowly decomposing leaves, or “duff” that promotes a rich community of wildflowers, tree seedlings and small animals. Earthworms change that environment dramatically by essentially consuming the duff, thereby destroying habitat and reducing fertility. In contrast to their effect in gardens, earthworms cause forest soils to become more compacted. As a result of habitat loss, fertility declines and soil compaction, these forests may be less productive and have poorer new tree regeneration in the long run.
Another view, from oligochaetologist (worm scientist) Rob Blakemore, is as follows:
Regarding popularized concerns about alien Asian invasive worms threatening to destroy American native forests, this may reasonably be regarded as part of a process that is commonly known as Ecological Succession [Odum 2005].
Ironically, the ecological concept of succession started with Thoreau and Cowles on studies of forest succession and on the Lake Michigan dunes. Large parts of the northeastern North America were glaciated up to about 10,000 years ago completely destroying all land surfaces and forming the Great Lakes. When the ice retreated Nature returned in successive waves and, gradually, the soil, vegetation, and animals communities re-established and species continue to evolve.
According to Darwin [1881] earthworms are supremely important for natural productivity and for the recent progress of human civilizations. In this context the woodlands of Michigan seem a relatively minor issue compared to species extinction and climate change. Healthy soils generally harbour earthworms and it appears there had been insufficient time for these slow-moving and flightless organisms to colonize without fast-track via incidental intervention of most-recent human settlers, often as anglers on the Great Lakes.
When exotic crops and stock were introduced around the world 10,000 years ago, so too were attendent earthworms and these have now spread to “pristine,” albeit transitional, woodlands. The many benefits earthworms have for agricultural and unmanaged soils may cause some changes in more natural habitats but this is a virtually unavoidable and irreversible force majeure and fact-of-life.
Certainly there will be a new ecological balance in time, possibly at a different level of productivity and biodiversity. That is the definition of succession.
Héry et al. 2008. Earthworms have been observed to increase methanotrophy (methane metabolic breakdown) in soil covering a landfill; this is most likely “due to the stimulation of bacterial growth or activity than to substantial shifts in the methanotroph community structure” [Hery 2008: 92].
Earthworm-mediated bioturbation has been linked to an increase in methanotrophy in a landfill biocover soil (AC Singer et al., unpublished), but the mechanism of this trophic interaction remains unclear. The aims of this study were to determine the composition of the active methanotroph community and to investigate the interactions between earthworms and bacteria in this landfill biocover soil where the methane oxidation activity was significantly increased by the earthworms [Hery 2008: 92].
And
We proposed the hypothesis that earthworms could stimulate the growth or the activity of methanotrophs. We showed that the earthworm-mediated increase of methane oxidation in the landfill biocover soil only weakly correlated with a shift in the structure of the active methanotroph population. Future work needs to focus on the relationship between this earthworm effect on enhanced methane oxidation in landfill cover soil and this effect on bacterial activity and growth. The possible contribution of an enriched population of nitrifying bacteria to methane oxidation also requires further investigation [Hery 2008: 101].
Water
With the rise of civilizations, humans began having significant impacts on bodies of water and the water cycle. The early “hydraulic civilizations” appeared along major rivers (Nile, Tigris-Euphrates, Indus, Yellow River and others), changed watercourses and built canals for agriculture and transportation. As populations and cities expanded, demand for food led to soil depletion while the built environment created growing areas of impermeable surfaces. Disruption of water cycles has reached a peak since the industrial revolution, with large areas of land covered with impermeable surfaces, and rainwater and waste rapidly shuttled away from land into the oceans. [Kravčik 2007: 42 ff.]
Eco-restoration requires a shift in thinking about water management and fortunately there are many successful water restoration projects under way, along with a strong theoretical basis to guide them. Water management is the key to cooling the biosphere, regenerating photosynthesis and drawing down carbon on billions of acres. Included in this section is a brief discussion of wetlands, which will be expanded in the next Compendium release.
Several eco-restoration examples are included here, and many were explored at our 2015 Restoring Water Cycles to Reverse Global Warming Conference at Tufts University – all speaker videos are available online. https://bio4climate.org/program-tufts-2015/.
Overview
Healthy Soils Australia, Walter Jehne 2015. Walter Jehne is a microbiologist, soil and climate scientist who has spent the past several years teaching and promoting the use of nature’s tools to address destruction of land and water cycles, and educating on methods important in addressing global warming. The text below represents excerpts (condensed and edited) from the paper, “Restoring Regional Rainfalls: Background Brief for Outcomes, Australia Forum on Vegetation-Rainfall Relationships”:
Contrary to the dominant assumptions that global warming is due to elevated atmospheric carbon concentrations,
- Systemic climate changes such as aridification began in the 1970s well before CO2 levels or its temperature effects increased abnormally.
- Water-based processes govern most climate effects and over 95% of the earth’s heat dynamics for billions of years, including some 80% of the natural greenhouse effect.
- These hydrological heat dynamics have been ignored or deemed “secondary feedbacks” to the CO2 greenhouse effect because water is assumed to be a dominant determinant of our climate, and humans could not possibly have altered these global cycles to cause the anthropogenic climate changes
- The hydrological processes are highly complex and difficult to model, and were therefore excluded in IPCC (Intergovernmental Panel on Climate Change) assumptions and models, whereas the minor CO2 component of the greenhouse effect is more readily modelled, and provides a simple marketable explanation of its “cause.”
- Because of these IPCC assumptions, policy and response options have largely ignored the dominant hydrological determinants on climate, the effects of land management, and impacts these changes may have on climate, water and bio-system stability.
Yet we have greatly altered the earth’s natural hydrology and thus heat dynamics by:
- Clearing over 75% (6.3 billion hectares or 15.75 billion acres) of the earth’s primary forest, greatly altering the land’s albedo and heat reflectance as well as transpiration and latent heat fluxes that cooled vast regions.
- Oxidizing and eroding organic matter from some 10 billion hectares of soils thereby reducing the ability of landscapes to infiltrate, retain and supply water to sustain cooling transpiration and latent heat fluxes and the drawdown of carbon from the air by plants via photosynthesis.
- Exposing vast areas of such degraded, cropped and bare soils to erosion which has dispersed 3 billion tonnes of additional dust aerosols into the air where it nucleates warming humid hazes that retain heat in the biosphere.
- Heating bare exposed soils to greatly increase their re-radiation of heat which massively increases greenhouse warming effects.
- Increasing the absorption of solar radiation by humid haze micro-droplets [resulting in] global dimming (while in the liquid phase), as well as the absorption of re-radiated heat (while in the gaseous phase) to warm the lower atmosphere via the water vapour greenhouse effect.
- Reducing regional rainfalls often by up to 30% due to the increase in persistent haze micro-droplets which are too small to coalesce into raindrops and precipitate by themselves.
- Increasing surface humidity due to the persistent humid hazes, thus lowering evaporation rates by up to 10% and reducing latent heat fluxes which transfer heat out of the biosphere into space.
- Reducing the production of the biological precipitation nuclei from forests that would help coalesce the humid haze micro-droplets to form dense clouds with high albedos that reflect 33% of solar radiation out to space, thereby regulating global temperatures.
- Preventing the nucleation of haze and cloud droplets into raindrops which lowers rainfalls and the level and longevity of transpiration, photosynthesis and cooling latent heat fluxes.
- Impairing the night-time escape of re-radiated heat to space via natural “radiation windows” due to the impaired nucleation and precipitation of such “blocking” humid micro-drop hazes.
- Increasing sustained high pressures above the cleared, bare heated soils that prevents the inflow of cool moist air from oceans, its precipitation and the associated cooling heat fluxes.
- Extending such high pressure over vast regions and periods to accentuate the aridification of bio-systems which readily collapse to deserts with further human land degradation. [Healthy Soils Australia 2015: 1-2]
Given this reality we need solutions that go beyond just reducing future CO2 emissions but also:
- Cool regions and the climate so as to offset dangerous warming and its feedback effects.
- Draw down carbon back into its safe soil sinks so as to reduce its greenhouse effect.
- Restore the resilience of agro-ecosystems and communities to the extremes and secure their essential water, food and bio-material needs via just, safe low carbon futures. [Healthy Soils Australia 2015: 8]
- Regenerate natural hydrological processes by land management which captures water in soils, wetlands, aquifers and biomass
- Maintain healthy biodiverse soils to Restore microbial drivers that govern these cooling hydrological processes by emitting condensation nuclei that lead to rainfall.
- Support the biological sequestration of carbon from the air into stable soil humates and glomalin to enhance the water held in the soil reservoirs that sustain the cooling latent heat fluxes.
- Support the production of microbial precipitation nuclei that coalesce the warming humid hazes into dense high albedo clouds that cool regions and generate critical cooling rainfalls.
- Promote the nucleation and enhancement of rainfall in key regions to maintain the latent heat fluxes, green vegetated habitats and the radiation windows that enhance nighttime cooling effects.
Only by regenerating our forests, soils and landscapes can we now restore the hydrological cooling processes that helped govern the natural heat dynamics and buffered climate of the blue planet. Such regeneration is now our only option to offset the dangerous climate feedbacks resulting from the warming induced by our landscape degradation and its associated abnormal rise in CO2 levels.
Fortunately viable practical options exist to enable us to do and directly benefit from this, at grass roots community level: tree by tree, hectare by hectare, region by region. While we face a global emergency and must all take responsibility for it, it can only be addressed locally via practical action on the ground by communities driven by their own self interest in securing a safe climate and future.
The good news is we can still avoid the pending extremes and collapse provided we focus on direct local action urgently to regenerate the health of each square metre of soil and each forest and tree. We have the abundant degraded land, sunshine, CO2, waste biomass and nutrients to do it with. To grow more green areas; by regenerating our soils, forests, rangelands and even re-greening deserts.
We can do this if we enhance the infiltration, retention and availability of each raindrop in our soils so that the regenerated ‘in soil reservoirs’ sustain healthy green growth over larger areas for longer. This will happen naturally, synergistically, as plant growth enhances the structure of the soil by increasing its carbon content which in turn aids its water holding capacity and nutrient dynamics.
Just as nature did over the past 420 million years in colonizing and greening the earth’s land surface, these same processes are now our only option to regenerate our soils, forests and landscape and re-secure our safe climate and future. [Healthy Soils Australia 2015: 7-11]
Water Article Summaries
Ellison 2017. “Trees, forests and water: Cool insights for a hot world” may be one of the few articles in the mainstream literature relating to climate that puts hydrological cooling effects before carbon in importance for addressing global warming, although dynamics of water and carbon are closely intertwined.
Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. [Ellison 2017: 51]
This paper is discussed further under Forests.
Evans, Griggs 2015. Carol Evans is a fisheries biologist and Jon Griggs is a rancher in northeastern Nevada. They have worked together over twenty-five years to restore overgrazed lands to health through planned grazing of cattle, which also brings water, trout, beavers and biodiversity to the riparian areas of Maggie and Susie Creeks. In the driest state in the U.S., with less than ten inches of rain a year, they now have perennial streams and wet meadows after five years of the worst drought in memory.


Susie Creek, ca. 1989 (left) and 2015 (right) after five years of drought. Elko, Nevada
Kravčík 2007. Michal Kravčík and co-authors are Slovakian hydrologists who have developed what they call a new water paradigm for managing water cycles, floods and drought.
In a healthy water cycle, while some rain enters streams and rivers directly and is carried off to sea, most rain water is absorbed by the soils in situ, where it lands. The rain gives life to the soil and sets many biological processes in motion, where it is essential for stable soil carbon storage and cooling the biosphere. This includes evapotranspiration from plants which returns water as vapor to the atmosphere where the water condenses and falls as rain. The cycle then begins anew. Kravčík et al. call this the “small water cycle” (i.e., local water cycle) where most water goes through its cycles in a regional area or smaller. The “large water cycle” is the exchange of water between oceans and land, and “above land water circulates at the same time in many small water cycles which are subsidized with water from the large [continental or global] water cycle.” [Kravčík 2007: 16]
Civilizations disturb healthy water cycles and accelerate the runoff from land by creating impermeable surfaces (including degraded farmlands and rangelands), and preventing water from remaining in place to sink into soils or to forcing it to run off the land, causing floods and often carrying valuable topsoil with it. Furthermore, water systems have been engineered to move water away from its source to the oceans. Water, with its growing use as a means to dispose of farming, industrial and human wastes, is even seen as a nuisance. As a result, less water returns to continents from the oceans than is lost from continents to oceans, which leads to desiccation of soils, severe drought, wildfires, desertification, and a measure of sea-level rise. There is a growing understanding that these phenomena, often attributed to climate change, may in fact also be a function of disrupted water cycles.


Restored urban land, Kosice. November 2005 (left), September 2006 (right).
Heat from the sun drives these earthly water cycles. Small water cycles are local, circulating water within a relatively small area. Latent heat causes water to evaporate; heat is absorbed in the process of evaporating water and does not result in an increase in local temperature. We thus do not experience latent heat as an increase in temperature. However, when there is less water available for evaporation, less solar energy is transformed into latent heat and more solar energy is transformed into sensible heat, heat you can feel as increased temperature. This is the heat that we are increasingly experiencing as global warming.
A great deal of heat is moved from the surface of the earth to the upper atmosphere by evaporation and transpiration of water by plants, contributing to significant cooling of the biosphere – to illustrate it takes 540 calories to turn 1 gram of water to vapor; by comparison it takes only 80 calories to melt 1 gram of ice.
Draining of land, that is, runoff and floods, can be reversed through comprehensive conservation of rainwater which maintains the sponge-like absorption capacity of soils and maintains many aspects of soil health, resilience, biodiversity and productivity. Renewal of small water cycles over land can temper extreme weather events and ensure a growth in water reserves by eliminating heat islands and problematic distribution of atmospheric moisture.
Nobre 2010. Antonio Nobre is an Amazon scientist who has studied the biotic pump (see also Makarieva), and tells how he was once told by an indigenous wise man,
“Doesn’t the white man know that, if he destroys the forest, there will be no more rain? And that if there’s no more rain, there will be nothing to drink, or to eat?” I heard that . . . [ and thought], “Oh, my! I’ve been studying this for 20 years, with a super computer; dozens, thousands of scientists, and we are starting to get to this conclusion, which he already knows!” A critical point is the Yanomami have never deforested. How could they know the rain would end? This bugged me and I was befuddled. How could he know that?
Some months later, I met him at another event and said, “Davi, how did you know that if the forest was destroyed, there’d be no more rain?” He replied: “The spirit of the forest told us.”
The equatorial region, in general, and the Amazon specifically, is extremely important for the world’s climate. It’s a powerful engine for evaporation. From a satellite viewpoint, atmospheric water flow can look like a geyser, which is underground water heated by magma transferred into the atmosphere. There are no geysers in the Amazon but trees play the same role. They, like geysers, transfer an enormous amount of water from the ground into the atmosphere. Nobre continues:
There are 600 billion trees in the Amazon forest, 600 billion geysers. That is done with an extraordinary sophistication. They don’t need the heat of magma. They use sunlight to do this process. On a typical sunny day in the Amazon a large tree manages to transpire 1,000 liters of water. If we take all of the Amazon, which is a very large area, and add up all the water that is released by transpiration, “the sweat of the forest,” an incredible amount of water is evaporated into the atmosphere: 20 billion metric tons of water per day. . . . This river of vapor that comes up from the forest and goes into the atmosphere is greater than the Amazon River.”
The Amazon River itself is the largest river on Earth, it carries one fifth of all the fresh water, it releases 17 billion metric tons of water a day into the Atlantic Ocean, smaller than “the river in the sky.” To evaporate the 20 billion tons of water released by trees it would take 50,000 of the largest hydroelectric plant in the world, Itaipus, which generates 14 GW of electricity, 30% of Brazil’s power. The Amazon does this with no technology, at no cost.
Schwartz 2016. Judith Schwartz once again travels the world to collect stories of lands restored, of lives revived, this time to glean insight from restorers of water. She demonstrates that many of our assumptions about managing water are derived from engineering, not biology. When biology is the focus of the water and rainfall question the problem is redefined, and clarified. Solutions that had been invisible become apparent, and provide the opportunity for far more effective responses – even in some of the driest places on earth. Floods and droughts become manageable, even preventable entirely. Two of the innovators mentioned in Water in Plain Sight, Michal Kravčík and Rajendra Singh, spoke at Biodiversity for a Livable Climate’s 2015 Restoring Water Cycles conference, as did Judy.
Singh 2007. Rajendra Singh, the “Water Man of India,” tells the story of how he helped over 1,000 villages restore water and abundance through the use of ancient, low-technology land management. Providing water for people, farms and animals, such efforts countered the ill effects of industrialization and reversed flight to the cities. Says Singh:
I am neither a scientist, nor a professional water engineer nor a climate change expert. I am a small constructive worker of Gandhi and I mobilize the civil society and the community for action on natural resources management and conservation for rural uplift in India. Here I am recording the impact of the above work on the ecology of 6,500 square km area in Alwar district from 1985‐2007. Since 1985, 8,600 small water harvesting talabs [a form of check dam] in 1,068 villages of Alwar district covering 6,500 square km area have been built. This has resulted in the shallow aquifer recharge in groundwater bringing up the water table from about 100‐120 meters depth to 3‐13 meters at present. The area under single cropping increased from 11 per cent to 70 per cent out of which area under double cropping increased from 3 per cent to 50 per cent bringing prosperity to the farmers. The forest cover, which used to be around 7 per cent increased to 40 per cent through agro‐forestry and social forestry, providing sufficient fuel wood and sequestering carbon from atmosphere [Singh 2007: 5].
A dramatic example of how large restoration efforts are built from small, local efforts. In the ten years since this paper, Tarun Bhagat Sangh has continued to expand its work.

Wetlands
Note: As mentioned in the Release notes, we have a small staff and therefore have had to postpone some important material to the next release, scheduled for January 2018. This will include a more thorough exploration of the importance of wetlands in addressing climate.
Wetlands only cover only a small proportion of the terrestrial surface area, with estimates ranging from 5-8% [Mitsch 2007]. Despite this, they store a disproportionate amount of Earth’s soil carbon, with estimates ranging from 20-30% [Lal 2008]. Soil carbon in wetlands can be as high as 40% [Vepraskas and Craft 2016; cf. Nahlik 2016: 2], compared to typical estimates for agricultural soils that range up to 2% [Lal 1995; see the Croplands section of this Compendium for a detailed discussion of agricultural soil carbon]. High carbon storage in wetlands is the result of anoxic conditions in wetland soils that lead to slowed decomposition and a resulting increase in stored organic matter. Wetlands have significant potential to act as carbon sinks under the right circumstances. Potentially reducing the carbon sequestration potential for wetlands are the methane emissions that they produce. Notwithstanding the uncertainty of methane emissions, wetlands globally likely serve as a significant net carbon sink. With the complexity of processes that contribute to wetland carbon exchange and resulting function as GHG sinks or sources, management decisions may be a critical factor in determining the carbon sequestration outcome of wetland ecosystems. [See Nahlik 2016 for review of quantitative data]
Overview
The importance of wetlands cannot be overstated. They exist in all biomes ranging from the tropics to the tundra, and on every continent except Antarctica . Wetlands also tend to host much higher concentrations of biological diversity than other ecosystems. The fact that the oldest intergovernmental environmental convention, the Ramsar Convention of 1971, was focused explicitly on wetland conservation is a testament to this importance [Mitra 2005].
The comparison of carbon dioxide storage to methane emissions is a source of uncertainty in determining the carbon sequestration potential for wetlands. Given methane’s stronger radiative forcing as a greenhouse gas, the IPCC currently uses a ratio of 25:1 to indicate the methane to carbon global warming potential. Despite the high global warming potential presented by methane, it has a relatively short atmospheric “lifetime”, often reported in the range of 8-12 years. Although methanotrophic (methane metabolizing) bacteria are not typically calculated into the equations regarding the atmospheric lifetime of methane, bacteria that metabolize atmospheric methane (e.g., high-affinity oxidation methanotrophs) provide an additional mechanism hastening methane attenuation [Jardine 2009].
Several trends appear to be instructive in wetland carbon storage. Tropical and temperate wetlands generally hold more carbon than northern boreal peatlands [Mitsch 2012: 7-9]. Freshwater inland wetlands may hold more carbon than coastal saline wetlands, though this may be based primarily on their surface area extent. More carbon storage occurs at depths from 30 – 120 cm (1-4 ft), at least as reported by an analysis of U.S. conterminous wetlands in which 65% of the total carbon was deeper than 30 cm (1 ft) [Nahlik 2016: 2-3]. A correlation exists between increased anthropogenic disturbance and decreased carbon storage, although this may be an artefact of settlement patterns rather than a causal relationship [Nahlik 2016: 4]. One study reported high carbon sequestration and low methane emissions in constructed wetlands [Mitsch 2012]; this is an encouraging result considering the many wetlands created as mitigations to compensate for human development impacts.
Wetland Article Summaries
Apfelbaum 1993. Steve Apfelbaum of Applied Ecological Services, Inc. is a restoration ecologist with several decades of experience around the world. This brief paper, “The Role of Landscapes in Stormwater Management,” describes the historical condition of wetlands in the upper midwest, the degrading effects of agriculture and urbanization on water cycles, vegetation and the resultant pollution. Included are recommendations for restoration of healthy wetlands and methods for slowing the movement of water so that it may keep soils hydrated and feed local ecosystems. In addition, restoration of wetlands includes high-capacity carbon storage due to low-oxygen conditions in wetland soils, with subsequent low rates of oxidation and loss of carbon to the atmosphere.
This paper presents evidence that many existing streams did not have conspicuous
channels and were not identified during pre-settlement times (prior to 1830s in the Midwestern United States). Many currently identified first, second, and third-order streams were identified as vegetated swales, wetlands, wet prairies, and swamps in the original land survey records of the U.S. General Land Office.
The data presented show that significant increases in discharge for low, medium, and high flows have occurred since settlement. Stream channels have formed inadvertently or were created to drain land for development and agricultural land uses. Currently, discharges may be 200 to 400 times greater than historical levels, based on data from 1886 to the present for the Des Plaines River in Illinois, a 620-square-mile watershed. Historic data document how this river had no measurable discharge or very low flow conditions for over 60 percent of each year during the period from 1886 to 1904.
This study suggests that land-use changes in the previous upland/prairie watershed have resulted in a change from a diffuse and slow overland flow to increased runoff, concentrated flows, and significantly reduced lag time. Preliminary modeling suggests the following results: reduced infiltration, reduced evaporation and evapotranspiration, greatly increased runoff and hydraulic volatility, and increased sediment yields and instream water quality problems caused by destabilization of streambanks.
The opportunity to emulate historical stormwater behavior by integrating upland landscape features in urban developments and agricultural lands offers stormwater management options that are easier to maintain, less expensive over time, attractive, and possibly more efficient compared with many conventional stormwater management solutions and the use of biofiltration wetlands.
and
Diverse and productive prairies, wetlands, savannas, and other ecological systems occupied hundreds of millions of acres in presettlement North America. These ecological systems have been replaced by a vast acreage of tilled and developed lands. Land-use changes have modified the capability of the upland systems and small depressional wetlands in the uplands to retain water and assimilate nutrients and other materials that now flow from the land into aquatic systems, streams, and wetlands. The historical plant communities that were dominated by deep-rooted, long-lived, and productive species have been primarily replaced by annual species (corn, soybeans, wheat) or shallow rooted non-native species (bluegrass lawns, brome grass fields). The native vegetation was efficient at using water and nutrients, and consequently maintained very high levels of carbon fixation and primary productivity. Modern communities, in turn, are productive but primarily above ground, in contrast to the prairie ecosystem where perhaps 70 percent of the biomass was actually created below ground in highly developed root systems. These changes in the landscape and vegetation coupled with intentional stormwater management have changed the lag time for water to remain in uplands and consequently increased the rate and volume of water leaving the landscape.
Mitsch et al 2012. This study evaluated the carbon storage and methane exchange potential for seven wetlands based on field data collected over several years, and used field data collected at 14 other wetlands globally to model the carbon sequestration and methane emission potential out to 300 years. A total of 21 wetlands were examined. The modeling accounted for the anticipated half-life of methane oxidative degradation in the atmosphere. Results indicated that methane emissions become unimportant within the 300 year model simulation time range, with most wetlands making the shift to net carbon storage by year 100 of the model. The study supports the potential for wetlands as carbon sinks.
Nahlik and Fennessy 2016. The objective of this article was to quantify the carbon stocks present in wetlands of the conterminous United States. To do so, the authors examined empirical field data collected during the 2011 National Wetland Condition Assessment conducted by the U.S. Environmental Protection Agency, and used this to quantitatively extrapolate to larger scale carbon estimates. These estimates were developed at regional and national scales. Results were evaluated by region, wetland type, freshwater or tidal status, and level of anthropogenic disturbance. The article indicated that 11.52 gigatons of carbon are present in the U.S., much of which is in soils deeper than 30 cm (1 ft). Freshwater wetlands located inland held nearly ten-fold as much carbon as intertidal wetlands overall, although this is at least partly due to the much greater aerial extent of inland freshwater wetlands; tidal wetlands still had higher concentrations of carbon storage. The authors also indicate a possible relationship between anthropogenic disturbance and carbon stocks, wherein less disturbed sites store more carbon. Insufficient data was available to determine whether this was a causal effect or an artefact of some kind, such as human preference in settlement patterns. The authors conclude that, due to the substantial carbon stocks that wetlands represent and the potential for anthropogenic impacts, existing intact wetlands should be protected to avoid the risk of further contributing to climate change.
Croplands
Cultivated land covers 1.6 billion hectares globally [FAO 2011]. About 62% of cropland produces food directly for human consumption, while 35% is dedicated to producing animal feed, and 3% to biofuel feedstock, seed and other industrial products [Foley 2011: 338]. Agriculture is a major source of emissions of carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O), contributing 10-12% (including crop and livestock production) of total greenhouse gas emissions [Smith 2007]. Agricultural emissions are driven by the globally dominant industrial model, which favors monocultures and fossil-fuel intensive inputs, and results in soil organic carbon loss and overall soil degradation. However, rather than being a source of carbon emissions globally, agriculture can become a powerful carbon sink. This section looks at the carbon sequestration outcomes of farming practices, such as cover cropping, agroforestry and no-till, which are designed to minimize erosion and boost soil biodiversity, thus restoring soil ecosystems to health and resilience. While more research is needed on holistic approaches that combine multiple soil-building practices, such as permaculture and agroecology, recent research suggests that restorative agriculture could sequester “more than 40% of annual emissions (an estimated 21 Gt CO2 each year [5.7 Gt C/year])” [Rodale 2014], and likely far more, as indicated below.
Cultivation thus began an ongoing slow ignition of
Earth’s largest surficial reservoir of carbon [16]
Overview
The purpose of this compendium, once again, is to emphasize possibilities, the “positive deviants” which lead us to expand our conceptual limits. Only when we can conceive of exceptional and inspiring outcomes may we find the motivation to overcome obstacles to attain them. Fortunately the evidence that supports regenerative land management is rapidly growing, and there are indications that it may outpace climate disruption and provide us with the time and opportunity to address the many difficult circumstances resulting from widespread eco-destruction, including the poster child, global warming. In this section we address the challenges of croplands and their ability to capture atmospheric carbon and recover quickly from millennia of mistreatment.
Under careful human management it is possible for soil organic carbon to reach amounts greater even than under natural, pre-agricultural conditions. A classic example is the Terra Preta soils of the Amazon, “where intensive management and high levels of organic matter additions were practiced over many years, resulting in greatly enhanced soil C” [Paustian 1997: 231].
In spite of a long history of soil carbon loss and a body of scientific literature that views carbon-poor soils as “normal,” many examples of building high levels of soil carbon exist among today’s ecologically minded land managers. California Farmers Paul and Elizabeth Kaiser, for instance, use 5-10 times more compost than average, never till, rotate fields with an extremely diverse mix of vegetable varieties, surround their crops with native trees, shrubs and flowers and have thus built up a thick topsoil containing 10% SOM [Oppenheimer 2015; Kaiser 2017].
In most scientific studies, carbon sequestration rates for croplands measure below 1t C/ha/yr (0.4t C/ac/yr), despite some exceptions as highlighted below. Leading soil scientist Rattan Lal [2016] estimates the global sequestration potential for cropland soils to be 0.8 to 1.2 t C/ha/yr, or “as much as 62 t/ha over the next 50 to 75 years … with a total C sink capacity of ~88 Gt on 1,400 Mha” [Lal 2016: 20A]. That amounts to an average annual global sequestration rate between 1 and 2 Gt C/year, compared to annual carbon emissions from fuel combustion and land use conversion of more than 10Gt C/yr [Lal 2016]. Similarly, Smith [2008] estimates that, under improved management, agriculture could offset 20% of global emissions. Both authors note that conservation-oriented agricultural is a small, albeit crucial, piece of the whole climate mitigation puzzle.
Yet, for a couple of important reasons, these estimates likely greatly underestimate the potential of global croplands to absorb carbon. First, samples are commonly taken to a depth of 30cm or less [Torres-Sallan 2017; Minasny 2017]. This is the default sampling depth recommended in the 2006 IPCC Guidelines for National Greenhouse Gas Inventories, despite acknowledgement in these same guidelines that land use and management is likely to have a major impact on deeper soil layers [FAO 2017b].
Indeed, significant amounts of carbon sequestration occurs in deeper soil profiles – even beyond a 1 m (3 ft) depth [Follett 2012, Liebig 2008, Schmidt 2011: 51]. Harper et al. found that half to three-quarters of total SOC to bedrock was in the surface 5 m with the remainder below that depth. The authors speculate that deep carbon may have been deposited directly by deep-rooting plants. “Where deep soils coincide with deep rooting the biological deposition of carbon from roots (and their associated biota) is inevitable at depths at which SOC has rarely been measured” [Harper 2013: 642].
Second, many studies measure sequestration rates for just one or two soil-building techniques, isolating them from additional, potentially synergistic, practices. In fact, intact ecosystems are based on countless synergistic relationships among organisms and their environment. In other words, many studies measure minor tweaks to conventional, industrial cropping systems.
For example, Minasny et al. [2017] compiled sequestration rates from around the world to assess the viability of the France-led “4 per 1000” initiative (seeking to halt the annual increase in atmospheric CO2 by increasing soil carbon by 0.4% per year). The authors estimate “that an annual rate of 0.2-0.5t C/ha/yr is possible after adoption of best management practices such as reduced tillage in combination with leguminous cover crops.” However, most of some 40 studies of best management practices on arable land assessed only one or two of many – often minimally improved – practices, such as “reduced use of summer fallow,” “rice-rice with NPK,” “inorganic fertilizer,” and “pasture.”
Similarly, an often-cited study by West and Post [2002], compiling 276 paired treatments from 67 long-term experiments, analyzes the sequestration rates for either increased rotation complexity (0.2+/-0.12t C/ha/yr) or a change from conventional tillage to no-till (0.57+/-0.14t C/ha/yr). While both practices were used at some sites, the data were not analyzed according to whether a single practice or combined practices were used. However, the authors suggest that using both practices together can be additive:
Data used in this analysis was stratified separately with regard to a change in tillage or a change in crop rotation. In practice, these changes could occur simultaneously. It can be inferred from our results that if of a decrease in tillage and an enhancement in rotation complexity occur simultaneously, the short-term (15–20yr) increase in SOC will primarily be caused by the change in tillage and subsequent decrease in the rate of SOC decomposition, while the long-term (40–60yr) increase in SOC will be primarily caused by the rotation enhancement and residue input and composition [West & Post 2002: 1943].
If moving to a combination of two restorative practices can increase carbon sequestration somewhat above the use of a single improved practice, then what is possible when many restorative practices are stacked one upon another within an agroecosystem? Permaculture, a design framework with “two broad conceptual criteria: ecosystem mimicry and system optimization,” where multiple restorative practices are indeed combined, represents a counterexample to industrial farming. Yet, sequestration rates from such a system have rarely, if ever, been measured. In fact, very little scientific study of any kind has been conducted in permaculture systems, despite the concept having been developed by scientist Bill Mollison, and adopted to favorable results globally for over 30 years [Ferguson and Lovell 2013].
In light of the centrality of agriculture’s role in ecosystem restoration due to the vast surface area it occupies, we present some literature representing agriculture’s maximum potential contribution to climate change mitigation, and argue that the focus of future research do the same.
Specifically, future studies should consider the effects of greater ecological intensity, diversity and potentially additive and synergistic interactions that can exist among multiple soil-building practices, rather than continuing to pursue measurement of their individual effects, which yield minimal outcomes. Future research must also measure SOC changes to greater depths in the soil horizon in order to capture the full benefit of any given practices. Such changes would likely present both a more accurate and more promising real-world potential for the climate mitigation potential of agriculture.
For a relevant and effective comprehensive assessment of regenerative management practices, one that supports the profound shift necessary in conventional 21st century agriculture, a scientific paradigm shift is necessary so that economics and policy will follow.
For 10,000 years, humans have been clearing patches of forest and grassland to plant crops. While clearing land by burning it visibly turns organic carbon into smoky CO2, plowing and tilling releases soil organic carbon by breaking up soil aggregates that protect carbon. Exposed soil organic carbon is consumed by microbes, and converted to CO2 through respiration. Tilling soil also subjects it to erosion. “Since tillage-based farming began, most agricultural soils have lost 30% to 75% of their soil organic carbon (SOC), with industrial agriculture accelerating these.” [Teague 2016: 157]
Agriculture is a source not only of CO2 emissions, but also of greenhouse gas emissions of methane (CH4) and nitrous oxide (N2O). In cropland soils, CH4 is produced by anaerobic decomposition of organic matter, usually in waterlogged soils like rice paddies. However, soils can also be a methane sink due to the presence of methanotrophic bacteria, which oxidize methane. N2O is produced by microorganisms, which transform excess ammonia fertilizer into nitrate and then N2O. “Upon conversion to NO2– or NO3–, excessive fertilizer N becomes subject to denitrification and thereby contributes to terrestrial emissions of N2O, which have been found to increase with the rate of N fertilization” [Mulvaney 2009: 2296].
Industrial agriculture compensates for soil carbon loss by abandoning degraded land or using chemical inputs for the nutrients and pest resistance that an otherwise carbon-rich, biologically active soil provides. However, the farming methods that rebuild topsoil without relying on synthetic inputs, while also ameliorating the worst effects of drought, are the same ones that can make agriculture a major sink for atmospheric CO2. Such methods, which can be used together as a complementary suite of practices include: no-till; cover-cropping; agro-forestry; diverse crop rotations, including integrating livestock grazing; use of compost, manure, and biochar; and use of deeper-rooting plants and perennials.
Cropland Article Summaries
Cover crops
Cover crops protect the soil during a time of year when no cash crops are growing and the soil would otherwise be bare. “Cover crops, also named inter-crops or catch crops, are crops that replace bare fallow during winter period and are ploughed under as green manure before sowing of the next main crop.” [Poeplau 2015: 34] Cover crops can also be rolled and crimped or mowed, instead of plowed, in preparation for the main crop.
Using cover crops reduces erosion, nutrient leaching, and drought stress, and add carbon through continued plant cover and growth as well as increase biodiversity. Leguminous cover crops also fix nitrogen. Furthermore, “in contrast to other organic amendments, a large part of the C input from cover crop is added as roots, which was found to contribute more effectively to the relatively stable carbon pool than aboveground C-input” [Poeplau 2015: 38].
Vick 2016. This Montana study demonstrates that leaving farmland fallow “depletes carbon stocks and thereby soil quality” [Vick 2016: 129], thus illustrating the importance of keeping land continuously covered with living vegetation. “Fallow” is the stage of crop rotation where no crop is grown.
In this study, a CO2 emissions rate of 1.35 tC/ha/yr (0.54 tC/ac/yr) was measured from land left fallow during the 2014 summer growing season; an adjacent field planted in winter wheat (summer 2013) and spring wheat (summer 2014) was a net carbon sink, measuring carbon input from the atmosphere into the soil at ~2 tC/ha/yr (0.8 tC/ac/yr) and ~1 tC/ha/yr (0.4 tC/ac/yr), respectively. Other parts of this study show a dramatic effect on area cooling as well as increased moisture and rainfall. These results occur only from ending the practice of fallowing.
The researchers observed that a widespread decline of land left fallow in agricultural areas of the Canadian Prairie Provinces coincided with a summertime cooling trend since the 1970s. They noted that extreme temperature events now occur less frequently than in the recent past, maximum summer temperatures have decreased by ca. 2° C (3.6° F), relative humidity has increased by some 7% and summer precipitation has increased by an average of 10 mm/ decade across parts of the Canadian Prairie Provinces. A remarkable 6 W/m2 summer cooling has been observed compared to a ca. 2.5 W/m2 warming globally since the dawn of the Industrial Era.
Even in degraded croplands, relatively small changes can lead to significant differences in rainfall, soil carbon sequestration, and ambient temperature. A 2016 study in Montana demonstrated the effects of reversing the practice of fallowing of wheat fields in the upper midwest. Fallowing is “the practice of keeping a field out of production during the growing season” (Vick 2016:129):
Fallow is a common management practice in the dryland wheat-growing regions of the northern North American Great Plains to conserve water for subsequent crops (Lubowski et al., 2006). Fallow however also increases erosion (Wischmeier, 1959) and soil carbon loss (Cihacek and Ulmer, 1995), and fallow-small grain management strategies are not considered sustainable from the soil conservation perspective (Merrill et al., 1999). [Vick 2016:130].
As a result of farmers’ experience, fallowing has progressively decreased across many areas of the northern midwestern plains since the 1970s, providing an environment suitable for comparison study:
The area of fallow in the Prairie Provinces of Canada has decreased from over 15 Mha in the 1970s to under 2 million ha at the present (Fig. 1) as producers have realized that the water-savings benefit of fallow is outweighed by the economic losses of not planting (Dhuyvetter et al., 1996). The area under fallow in the United States has likewise decreased from 16 Mha to 6 Mha across the same time frame (Lubowski et al., 2006), largely in the northern Great Plains and other areas of the semiarid West . . . Despite the decreasing trend in fallow area across the North American northern Great Plains, fallow remains common in many regions including major land resource area
(MLRA) 52 in north-central Montana – the largest wheat-growing region in the state – where some 40% of agricultural lands may remain in fallow in any given year. In contrast, fallow has been reduced in northeastern Montana (MLRA 53) by hundreds of kha over the past decade (Long et al., 2014, 2013) as producers have adopted continuous cropping or alternate cropping practices. [Vick 2016:130].
The effects of this relatively simple change of practice led to some remarkable results:
The widespread decline of fallow in agricultural areas of the Canadian Prairie Provinces (Fig. 1) has coincided with a summertime cooling trend since the 1970s (Betts et al., 2013a, 2013b; Gameda et al., 2007; Mahmood et al., 2014). Extreme temperature events now occur less frequently than in the recent past, maximum summer temperatures have decreased by ca. 2০C, relative humidity has increased by some 7% (Betts et al., 2013b), and summer precipitation has increased by an average of 10 mm/decade across parts of the Canadian Prairie Provinces (Gameda et al., 2007). A remarkable 6 W/m2 summer cooling has been observed (Betts et al., 2013a); for reference, anthropogenic greenhouse gasses are responsible for a ca. 2.5 W/m2 warming globally since the dawn of the Industrial Era (IPCC, 2007). These climate benefits have only occurred during the growing season; fall, winter, and early spring temperatures have followed global trends (Betts et al., 2013b) . . . In other words, the observed regional climate cooling is broadly consistent with the effects of fallow avoidance on climate processes. [Vick 2016:130-131]
As dramatic as some of these changes are with only reduced fallowing, there are other land-management practices with significant impacts on water cycles, soil carbon, biodiversity and productivity. Such practices hold additional potential, and include cover-cropping and green mulches, pasture cropping, elimination of synthetic inputs which encourage renewed activity of important soil biota, especially worms – and perhaps most importantly on grasslands that co-evolved with animals, the reintroduction of animals themselves.
Finally, it is worth noting that there may be a significant underestimation of surface area and volume of soils in grasslands, as well as in other ecosystems, since natural topographies are not uniformly flat. Topographical variations would add volumes of soil carbon, water, etc. to prior estimates of areas that are typically calculated on the basis of two-dimensional map projections [Blakemore 2016: Fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regenerative management, carbon capture and water storage than is conventionally assumed. Such adjustments to soil volume calculations would positively affect carbon drawdown estimates in considering the potentials of eco-restoration in climate (see section, Do We Have More Soil for Carbon Storage than We Thought?).
Pimentel 2011. Arguing for cover crops as an effective way to reduce erosion and conserve nutrients in soil, Pimentel notes that “Growing cover crops on land before and after a primary crop nearly doubles the quantity of solar energy harvested in the agricultural system per hectare per year. This increased solar energy capture provides additional organic matter, which improves soil quality and productivity.” [Pimentel 2011: 41]
Crop rotation
Crop rotation diversification can enhance pest resistance, nitrogen input (when leguminous crops are added), soil penetration for better water infiltration (when deeper rooting plants are added), and residue input (when crops that produce more biomass are added). The effects on carbon sequestration from increases in crop rotation diversity vary depending on what crops are included. “Crop species can vary significantly in growth patterns, biomass production, water requirements, and decomposition rates, all of which affect net GHG emissions. Therefore, many rotations could be adapted with alternative species or varieties of annual crops to promote soil C sequestration—increasing root and residue biomass, increasing root exudates, or slowing decomposition—or otherwise reduce emissions” [Eagle 2012: 13].
Clearly, crop rotation is something of an umbrella term, describing a variety of practices, and even leaving space for practices that would not seem to offer much in the way of soil restoration. For example, as West and Post [2002] state,
. . . enhancement of rotation complexity refers to (i) a change from monoculture to continuous rotation cropping, (ii) a change from crop–fallow systems to continuous monoculture or rotation cropping, and (iii) an increase in the number of crops used in a rotation cropping system. In this analysis, continuous cropping is a cropping system without a fallow season, monoculture is a system with only one crop grown, and rotation cropping indicates two or more crops rotated over time on the same unit of land. [West & Post 2002: 1931]
Thus, even “continuous monoculture” can be considered as a crop rotation meant to increase carbon sequestration capacity. On the other hand, crop rotation can also involve great diversity, such as at Paul and Elizabeth Kaiser’s farm, where 3-7 crops/year rotate through vegetable beds, sometimes intercropped two crops at a time [Kaiser 2017].
Teague 2016. This study argues for greater use of no-till, cover crops, and crop rotation, including integrating livestock rotation into cropping systems.
Crop production can be managed to maintain permanent ground cover through the rotation of forage and row crop mixes, including cover crops, and legumes to increase soil fertility by fixing N. Grazing livestock can accelerate nutrient cycling through the consumption and decomposition of residual aboveground biomass.” [Teague 2016: 159]
The authors present a set of testable hypothetical scenarios suggesting the adoption of conservation cropping and adaptive management grazing (including grass-finishing cattle).
No-till
No-till (NT) allows farmers to plant without disturbing the soil, thus protecting it from water and wind erosion, leaving soil aggregates intact, and preventing a flush of oxygen from activating microbial breakdown of organic matter and releasing CO2. No-till can contribute to climate mitigation both by reducing emissions from the turnover of soil organic matter caused by tillage, and by sequestering carbon, especially in the surface layer [Mangalassery 2015].
Brown 2016. North Dakota Farmer Gabe Brown began practicing no-till in 1994. Since then, he has added cover crops (a diverse mixture of 70 species), complex crop rotations, orchards, livestock grazing (including cattle, sheep, pork and chicken), vegetable production, and bees. Through a long-term commitment to building the soil through no-till, keeping the ground always covered, and favoring as much biodiversity as possible (including a wide diversity of cash crops), Brown reports SOM has increased from 1.7% in 1993 to 11% in 2013. Furthermore, water infiltration has increased from ½ inch to more than 14 inches over the same time span.
Follett 2012. Measured to a depth of 150 cm (~5 ft), no-till continuous maize grown in eastern Nebraska, fertilized with 120 kg/ha of nitrogen and stover left on the field after grain harvest, sequestered 2.6 tC/ha/yr (1 tC/ac/yr). Notably, more than 50% of sequestered carbon was found below 30 cm (1 ft), illustrating that studies failing to sample below this depth (a common practice) risk greatly underestimating sequestration rates.
Organic vs. synthetic inputs
Organic farming uses “cultural, biological, and mechanical practices that support the cycling of on-farm resources, promote ecological balance, and conserve biodiversity” according to the USDA, which prohibits the use of most synthetic pesticides and fertilizers on certified organic farms. Organic farmers must find alternatives to synthetic inputs for managing pests and fertility. For example, vermi-composting is commonly used in organic farming. It is a natural and proven enhancement of the humification process that uses specific earthworms (e.g. Eudrilus eugeniae [Blakemore 2015]) to rapidly convert all organic “wastes.” Returning this vermicompost to soil renders synthetic fertilizers and pesticides unnecessary, as vermicompost often confers natural resistance to pests [Howard 1945, Balfour 1975] and it enhances resident earthworms [Blakemore 2000, 2016a; see also Earthworms section].
While the organic law provides a baseline for organic practices, the term “organic” encompasses a wide range of approaches to farming. For instance, some organic farmers may do little more than substitute naturally occurring inputs into an otherwise conventional, industrial operation, likely leaving the soil similarly depleted. Other organic farmers put into practice several of the methods mentioned in this section, aiming to truly build the functionality of the soil to resist pests and provide fertility. The studies included below highlight benefits from organic inputs and problems that come with using synthetic fertilizers with respect to soil carbon and biodiversity.
Johnson 2017. Using fungal-dominant compost in a 4.5-year trial at Leyendecker Field Research Site in New Mexico, researchers recorded an annual carbon sequestration rate of 10.7t C/ha/yr (4.8t C/ac/yr). Based on the observed trajectory of increasing productivity, they estimate a potential rate of 19.2t C/ha/yr (7.67t C/ac/yr). Chief investigator David Johnson found that increased plant growth is correlated most closely with the fungal to bacterial ratio. At a fungi:bacteria ratio of 0.04, only 3% of carbon flow went into plant biomass production, with the remainder of the carbon going into other functions, including nitrogen fixation, the soil, and respiration. At a fungi:bacteria ratio of 3.68, plant growth was more efficient with 56% of carbon flow going to biomass production.
Rodale 2014. Compiling data collected from around the world, Rodale Institute concluded that if all cropland were converted to their regenerative model[17], it would sequester 40% of annual CO2 emissions. Adding pastures to that model would add another 71%, effectively exceeding the world’s yearly carbon dioxide emissions.
On-farm soil carbon sequestration can potentially sequester all of our current annual global greenhouse gas emissions of roughly 52 gigatonnes of carbon dioxide equivalent (GtCO2e). Indeed, if sequestration rates attained by exemplar cases were achieved on crop and pastureland across the globe, regenerative agriculture could sequester more than our current annual carbon dioxide (CO2) emissions. Even if modest assumptions about soil’s carbon sequestration potential are made, regenerative agriculture can easily keep annual emissions within the desirable lower end of the 41-47 GtCO2e range by 2020, which is identified as necessary if we are to have a good chance of limiting warming to 1.5°C. [p.5]
Ryals and Silver 2013. This study examined the effects on plant growth and respiration from compost application on annual grassland in both coastal and valley sites in California. They found that a single application of compost during the three-year study resulted in a carbon sequestration rate of 1.45t C/ha/yr (0.58t C/ac/yr) and 0.54t C/ha/yr (0.22t C/ac/yr) at the valley grassland and coastal grassland, respectively. This enhanced net primary productivity was partially offset by CO2 emissions from increased respiration, but the compost did not affect CH4 or N2O fluxes. The authors conclude that:
Our results have important implications for rangeland management in the context of climate change mitigation. Urban and agricultural green waste is often an important source of greenhouse gas emissions (IPCC 2001). Here we show that an alternative fate for that material can significantly increase NPP and slow rates of ecosystem C losses at the field scale. This approach provides important co-benefits to landowners, such as the sustained increase in forage production measured here [Ryals & Silver 2013: 56].
While these results are low compared to some of the other studies noted here, this study illustrates positive use for green waste, and a potential tool that may contribute to climate-positive management.
Khan 2007. This five-decade study of nitrogen fertilization effects on SOC in Illinois shows that, despite progressively greater corn crop residue input during the second half of the 20th Century (increasing from 20,000 or 30,000 to 69,000 plants/ha since 1955), partly due to synthetic fertilizer use, SOC content did not increase, and in most cases declined. SOC declines were most pronounced in subsurface (16-46cm) of the soil horizon, compared to the surface layer (0-15cm). These results are despite crop residue being incorporated, rather than removed, in most plots since 1955, and in all plots since 1967.
These findings implicate fertilizer N in promoting the decomposition of crop residues and soil organic matter and are consistent with data from numerous cropping experiments involving synthetic N fertilization in the USA Corn Belt and elsewhere, although not with the interpretation usually provided. [Khan 2007: 1821]
Perennial systems, agroforestry, and permaculture
Unlike annual plants, perennials live for many years – thousands of years in some cases. Because of their deep (>2m, or 6 ft) and extensive root system, and longer growing seasons, perennials are likely to sequester carbon better than annual cropping systems [Glover 2007].
Agroforestry is the practice of integrating trees (a type of perennial) into a cropping system, including alley cropping, windbreaks, riparian buffers, silvopasture, and forest farming [Eagle 2012; Nair 2009]. Agronomic practices are notable for adding significant amounts of carbon to aboveground biomass, which is often measured separately from soil organic carbon sequestration [Nair 2009]. One of the strengths of agroforestry is its enhancement of an agroecosystem’s functional diversity:
The utilization of the environment by species includes three main components: space, resources, and time. Any species utilizing the same exact combination of these resources as another will be in direct competition which could lead to a reduction in C sequestration. However, if one species differs in utilization of even one of the components, for example light saturation of C3 vs. C4 plants,[18] C sequestration will be enhanced.” [Udawatta 2011: 19]
Toensmeier 2017. Compiling carbon sequestration rates from individual studies, reviews, and expert estimates, and organizing them into groups of annual versus perennial systems, woody versus herbaceous crops, and polyculture versus monoculture, Toensmeier observes that “the general trend is that systems that incorporate trees sequester more carbon.” The highest sequestration rate listed, 18 tC/ha/yr (7.2 tC/ac/yr) falls into the perennial woody polyculture group, and more than half of all sequestration rates listed under perennials are more than 6 tC/ha/year (2.4 tC/ac/yr), while most rates for annual cropping systems are less than 1t C/ha/yr (0.4 tC/ac/yr).
Lawton 2016. On 10 acres of the Arabian Desert in Wadi Rum, Jordan, Permaculture Designer Geoff Lawton built an organic, multi-species food forest on what had previously been bare desert ground. Using wastewater from a nearby irrigated farm to get started, he designed a microclimate that would prevent evaporation in every way possible. Key elements included: date palm trees for wind protection and shade; smaller fruit tree and trellised grapevines for additional shade; a succulent ground cover, which also catches nutrient-rich desert dust; a shaded swale for irrigation; and cut legume trees for mulch.

From https://www.facebook.com/greeningthedesert2/.
Lawton sought to “build organic matter within the system as quickly as possible with any living elements that will achieve those ends.” Once the soil came alive, it became productive. Lawton explains that strategic arrangement of the space is especially important in the desert. That’s why crops were grown in two rows in between three slightly wider rows of mixed fruit trees for protection. After four years, this orchard/farm was producing an abundance of fruits and vegetables, showing that it is possible to work with nature and avoid industrial inputs to achieve a productive landscape even in the harshest environment.
DuPont 2010. A Land Institute study measured the effect on soil properties and biota from perennial polyculture systems as compared to annual grain crop systems. Since the latter are typically intensively managed, “the effects of tillage and plant community composition are often confounded” [DuPont 2010: 25]. To control for management effects, this study compared the soil carbon and root biomass outcomes from no-tilled annual crops (rotation of soybean, sorghum and wheat) versus a perennial polyculture. Total root biomass in no-till annual plot measured at only 43% of that in a perennial grass plot in the top 1m of soil. Also, the authors found significantly higher levels of readily oxidizable carbon (ROC) and microbial biomass in the perennial plots compared to the annual crop plots. ROC measures soil carbon that is more available to soil microbes.
Small changes in ROC and other labile fractions of SOC may provide an early indication of soil degradation or improvement in response to management practices. Changes in active carbon pools can be two to four times greater than changes in total C after the initiation of new management practices and they are more highly correlated with other soil quality indicators including microbial respiration, aggregate stability and plant productivity [DuPont 2010: 28].
The authors conclude that “even in the absence of tillage and under best management practices, annual cropping can reduce soil carbon and impact soil biota and food webs important in nutrient cycling after just three years” [DuPont 2010: 25].
Soto-Pinto et al. 2009. In this southern Mexico study of land-use change in various agroforestry systems, the authors show that converting “traditional fallow” (secondary growth woods following cropping, averaging 23.4 years in age) to maize (with beans, squash and pepper) production results in 94% loss of living biomass carbon. However, transitioning to (a) “taungya” (maize, beans, squash and peppers intercropped between rows of timber and multipurpose trees), (b) shaded coffee systems, or (c) “improved fallow” (adding timber trees to traditional fallow plots) preserves living biomass carbon. This study points to the mounting relevance of agroforestry systems that can provide economic benefits to small-scale farmers, while avoiding carbon emissions from land use change from forest to agriculture and livestock production, which accounts for 35% of total emissions in Mexico, according to the authors.
Association for Temperate Agroforestry 2004:
Agroforestry practices are intentional combinations of trees with crops and/or livestock which involve intensive management of the interactions between the components as an integrated agroecosystem.
Intentional: Combinations of trees, crops and/or animals are intentionally designed and managed as a whole unit, rather than as individual elements which may occur in close proximity but are controlled separately.
Intensive: Agroforestry practices are intensively managed to maintain their productive and protective functions, and often involve annual operations such as cultivation, fertilization and irrigation.
Interactive: Agroforestry management seeks to actively manipulate the biological and physical interactions between the tree, crop and animal components. The goal is to enhance the production of more than one harvestable component at a time, while also providing conservation benefits such as non-point source water pollution control or wildlife habitat.
Integrated: The tree, crop and/or animal components are structurally and functionally combined into a single, integrated management unit. Integration may be horizontal or vertical, and above- or below-ground. Such integration utilizes more of the productive capacity of the land and helps to balance economic production with resource conservation.
Liebig 2008. Measured to a depth of 120 cm (~4 ft), switchgrass grown for bioenergy at 10 farms across the Great Plains in the United States sequestered 2.9 tC/ha/yr (1.16 tC/ac/yr). Of that, only 1.1 tC/ha/yr (0.44 tC/ac/yr) was found in the first 30 cm (1 ft) depth, with the remainder measured below 30 cm. The authors explain what makes switchgrass effective in carbon sequestration:
Increases in SOC [soil organic carbon] under switchgrass were likely caused by belowground C input from root biomass and rhizodeposition and decreased soil organic matter losses by erosion. Research conducted by ecologist John Weaver and his graduate students over 60 years ago provide ancillary support for increased SOC under switchgrass. Their detailed surveys of prairie grass roots indicated switchgrass to have the deepest root system of all grasses examined, with roots extending to a soil depth of 3m (~10 ft). This finding, coupled with observations that prairie grass roots regenerate by replacing dying roots with new, live roots indicates the potential for significant C input to the soil under switchgrass.
Montagnini & Nair 2004. Agroforestry systems are multifunctional with respect to carbon capture. Agroforestry can: increase the soil carbon content and fertility of cropland, while allowing for continued food production; create greater sequestration efficiency through diversity of vegetation; and allow for harvest of forest products, potentially keeping carbon sequestered in wood products for many years, and thereby also decreasing pressure on natural forests. And because of the mixed use of agroforestry systems:
[T]he amount of biomass and therefore carbon that is harvested and ‘exported’ from the system is relatively low in relation to the total productivity of the tree (as in the case of shaded perennial systems). Therefore, unlike in tree plantations and other monocultural systems, agroforestry seems to have a unique advantage in terms of C sequestration [Montanigni & Nair 2004: 285].
A few sequestration rates highlighted in this article include: A Costa Rica study of cacao grown under two different species of shade trees Erythrina (a leguminous tree) and Cordia (a timber tree), measured C sequestration in perennial plant biomass at an average of 4.28t C/ha/yr (1.7t C/ac/yr) for the cacao-Cordia system, and 3.08t C/ha/yr (1.2 tC/ac/yr) in the cacao-Erythrina system . In another study, tropical smallholder agroforestry was projected to sequester 1.5-3.5t C/ha/yr (0.6-1.4 tC/ac/yr).
Onim 1990. Tropical agroforestry was observed to increase SOC (soil organic carbon), at the 0-30 cm depth, to a maximum of 8.34 tC/ha/yr (3.38 tC/ac/yr) and minimum of 0.73 tC/ha/yr (0.30 tC/ac/yr).
Biochar
Biochar is organic matter that has been decomposed through pyrolysis (burning) under controlled, low-oxygen conditions, where it emits relatively little CO2. Biochar is then added to the soil for long-term carbon storage and/or enhancing availability of soil nutrients, oxygen and water to plants and microbes. Because charred biomass has been observed to persist in the soil for centuries or millennia, biochar is seen as a stable or recalcitrant form of carbon that that may prove to be a useful tool for reversing climate change. Not only is the biochar itself a stable form of carbon that can remain in soils long-term, but also it helps build healthy soil structure which increases plant growth and therefore photosynthetic capacity, resulting in carbon being removed from the atmosphere and stored in biomass or soils. [McLaughlin 2017; Taylor 2010; Paustian 2016; Weng 2017; Remediation Magazine 2017]
It is worth noting that depending on the pyrolysis technique, the resulting biochar may range in quality from poor to excellent. One hopes that as the industry matures, the understanding of the importance of biochar quality in assessing results will grow as well.
McLaughlin 2017. Hugh McLaughlin, Ph.D., P.E. is an expert on the properties and production of chars created by pyrolyzing biomass, and the subsequent conversion to activated carbons. He has published extensively on biochar and biomass-derived heat production. In this video he gives a short but comprehensive review of the qualities and use of biochar.
Paustian 2016. Biochar application to soils is considered in this article among several activities (such as compost application, cover cropping, residue retention, no-till, and others, as previously mentioned in this compendium) designed to increase soil C stocks by increasing organic matter inputs or reducing decomposition rates. Biochar acts as a soil amendment stimulating plant growth, thereby allowing for greater C storage through greater biomass production, while also embodying a generally stable form of buried carbon.
Biochar mineralizes 10–100 times more slowly than uncharred biomass. Thus a large fraction of added C … can be retained in the soil over several decades or longer, although residence times vary depending on the amendment type, nutrient content and soil conditions (such as moisture, temperature and texture).
However, because the organic matter originates from outside the ecosystem ‘boundary’, a broader life-cycle assessment approach is needed, that considers the GHG impacts of: (1) offsite biomass removal, transport, and processing; (2) alternative end uses of the biomass; (3) interactions with other soil GHG-producing processes; and (4) synergies between these soil amendments and the fixation and retention of in situ plant-derived C. In many cases, net life-cycle emissions will largely depend on whether the biomass used as a soil amendment would have otherwise been burnt (either for fuel, thereby offsetting fossil fuel use, or as waste disposal), added to a landfill, or left in place as living biomass or detritus [Paustian 2016: 50].
Remediation Magazine 2017. A popular report on Weng 2017, quoting the authors:
The project’s leader, DPI [Department of Primary Industries] researcher and SCU [Southern Cross University] adjunct professor Lukas Van Zwieten said the research threw up some unexpected results. “We immediately saw an increase in soil carbon from the biochar, as expected, but what we didn’t expect was that soil carbon content continued to increase. This research demonstrates the ongoing benefits of biochar in farming systems to improve pastures and grasslands and increase farmers’ production and profitability . . . the researchers found that biochar enhanced the below-ground recovery of new root-derived carbon by 20% – that is, more of the carbon photosynthesised by plants was retained in the biochar-amended soil. Biochar accelerated the formation of soil microaggregates via interactions between organic matter and soil minerals, thus stabilising the root-derived carbon. . . . The increased microbial activity and improved physical structure of the soil would also ultimately improve the effectiveness of fertiliser use, making the application of biochar particularly beneficial for high-end, intensive crop production”
“[T]he improved structure of the soil protected the naturally occurring carbon, as well as the carbon added”, said Southern Cross University’s associate professor Terry Rose, a co-author of the study. “Importantly, the biochar also slowed down the natural breakdown of native soil organic carbon by more than 5%.
Taylor 2010. An anthology of articles written by biochar pioneers. Covers biochar history, testing, production, challenges and uses. Suitable reading for general audiences as well as land management and industry professionals.
Weng 2017. Biochar can increase the stable C content of soil. However, studies on the longer-term role of plant–soil–biochar interactions and the consequent changes to native soil organic carbon (SOC) are lacking. . . . We found that biochar accelerates the formation of microaggregates via organo-mineral interactions, resulting in the stabilization and accumulation of SOC in a rhodic ferralsol (s.a. Remediation Magazine 2017).
Grasslands
Grasslands have been estimated to cover approximately 40% of global land surface area, approximately 5.25 bn ha (~13 bn ac ), except for Greenland and Antarctica [Suttie 2005; White 2000:12]. Their deep soils are rich repositories of nutrients, especially carbon and water. Many grasslands are anthropogenic, i.e., resulting from various land management techniques to maintain land for grazing and crop production by humans. Virgin grasslands are increasingly rare, possibly leading to significant underestimations of their potential positive contribution to productivity, and to carbon and water storage. Grasslands are important repositories of biodiversity, and have significant impacts on weather and climate. Here we review research and articles that indicate soil carbon storage potentials of roughly 13 gigatons per year (the equivalent of 6.5 ppm) if global grasslands were managed regeneratively.
Overview
While we have separate sections for Grasslands, Croplands and Soils, there is inevitable overlap. Many croplands are modified grasslands, and both are, of course, based in soils. Yet there are enough differences in each area of study to merit separate sections, keeping in mind that systemic behaviors and interactions are broadly applicable.
Since the onset of agriculture over 10,000 years ago with land management techniques that expose soil to air, estimates of up to 537 gigatons of soil carbon have been oxidized to carbon dioxide and other greenhouse gases [Buringh 1984: 91]. Even so, soils (>2,300 Gt) currently hold almost as much carbon as plants (550 Gt), atmosphere (800 Gt) and ocean surface waters (1,000 Gt) combined [NASA 2011], and almost surely retain the potential to store enough atmospheric carbon to return to pre-industrial levels.
Typical soil studies examine the first 30 cm (1 ft) of soil depth, but more recent investigations indicate that major soil carbon storage takes place deeper than that, often in a more stable form [Liebig 2008, Follett 2012, Harper 2013]. A USDA paper found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr) [Follett 2012]. A study of switchgrass for bioenergy found rates of SOC increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft) [Liebig 2008]. On an intensively grazed former row-crop agriculture land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions [Machmuller 2015]. In addition, the ultimate methane emissions may have been markedly less than measured since the report did not consider methane breakdown into CO2 from methanotrophic bacteria and atmospheric hydroxyl radical oxidation, with a significant reduction of methane’s ultimate greenhouse gas impacts.
These reports demonstrate the potential for massive amounts of soil carbon storage, significant cooling of the biosphere, and dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Grassland Evolution
Grasslands have long been a rich repository of carbon, both stable and labile. The co-evolution of grasslands with grazing ruminants has contributed to dramatic global cooling over the past 50 million years as a result of significant photosynthetic carbon drawdown into grassland soils [Retallack 2013]. Thus, grasslands are more than a consequence of geophysical changes, they are
. . . a biological force in their own right (Retallack 1998), in some ways comparable to the human rise to dominance of planetary resources (Vitousek et al. 1997). Grasslands have long been considered products of the coevolution of grasses and grazers (Kovalevsky 1873). Few plants other than grasses can withstand the high-crowned, enamel-edged teeth and hard hooves of antelope and horses. Yet these same animals are best suited to the abrasive gritty opal phytoliths and dust of flat, open grasslands. Grasses recover readily from fire and nurture large herbivores such as elephants: both fire and elephants promote grassland at the expense of wood land (Retallack 1997b; Jacobs et al. 1999). Grasses suppress insect and fungal attack with secondary metabolites such as cyclic hydroxamic acids (Frey et al. 1997). Grasses create Mollisols, unique soils with fine crumb clods rich in organic matter (Retallack 1997b). (Retallack 2001:407) [Emphasis added.]
Occupying such vast areas of planetary land surface, grasslands have a major influence on the global climate:
CO2 and CH4 (which rapidly oxidizes to CO2 ) are important greenhouse gases, and mechanisms for burial of their C may result in climatic cooling (Berner 1999). The most important long-term C sink from grasslands is their supply by erosion to sedimentary basins of crumb peds, which are unusually rich in organic matter intimately admixed with clay (Pawluk and Bal 1985). Tropical forests, in contrast, yield highly oxidized spherical micropeds with virtually no organic content (Retallack 1991a).” (Retallack 2001:414)
While there are other soils that are greater carbon sinks, such as peat bogs, wetlands and coastal habitats (e.g., mangroves, seagrasses), for volume and depth of carbon storage on vast areas of land, grasslands have enormous potential:
Grasses themselves are C sinks, especially considering their mass of roots and rhizomes underground. . . . [Numerous soil investigations] indicate that past estimates of organic C in tropical grassland soils have been low, in part because soils were not analyzed to sufficient depths. . . . Grassland and woodland soils may have comparable amounts of organic C in the surface 15cm. Beyond that depth, organic C values drop off dramatically in woodland soils but remain high in grassland soils to a meter or more. The fine structure and fertility of grassland soils is in large part due to this large C reservoir. (Retallack 2001:415)
Conventionally, it is estimated that approximately 40% of global land surface area is grasslands (52.5 million square kilometers, or ~5.25 billion hectares, or ~13 billion acres [Suttie 2005]), except for Greenland and Antarctica [White 2000:12; see Figure 1, below]. This is likely a significant underestimation of soil surface area and volume, since grasslands are not uniformly flat, with topographical variations adding carbon, water, etc. to areas that are typically calculated on the basis of a two-dimensional map projection [Blakemore 2016: fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regeneration, carbon capture and water storage than is conventionally assumed (see the section, “Do We Have More Soil for Carbon Storage than We Thought?”).
Natural grasslands are typically areas of low and seasonal rainfall. Unlike temperate environments with year-round rainfall, semi-arid and arid grasslands are dependent on grazing animals as a keystone species. The habits of grassland plants are as dependent on grazing animals as the animals are dependent on the plants that grasslands provide as food.
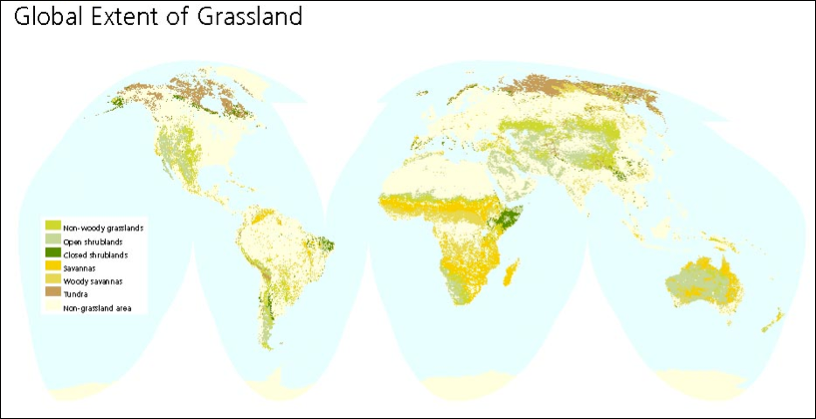
Figure 1: Global Extent of Grassland (White 2000:12).
Grazing animals are ruminants and as long as they are grazed in herds that move frequently, as they do in natural habitats in the company of predators, ruminant species are often interchangeable insofar as grassland health is concerned. Bison and antelope, for example, may be the wild ruminants that roamed the prairies and savannahs, but domestic cattle, when properly managed (human herders are the equivalent of wild predators), serve the same ecosystem functions.
Pioneering Work Of Allan Savory
Allan Savory, a wildlife biologist from Zimbabwe, began studying desertification in the 1950s. He pioneered an approach that he has termed Holistic Planned Grazing (HPG) for regenerative management of grasslands. He noted that there are essential differences between temperate grasslands, which he termed “non-brittle” environments, and arid and semi-arid grasslands, which he referred to as “brittle.” These distinctions are critical in understanding how different habitats require different management approaches.
Non-brittle environments, because of year-round rainfall, are relatively forgiving of mismanagement that destroys soil biota and exposes soil to sunlight, air and the elements. Recovery from soil degradation can be be rapid. Brittle environments, to the contrary, are fragile and easily desiccated, and when poorly managed, either from overgrazing or undergrazing, may take decades or centuries to recover or even ultimately turn to desert.
Brittle environments particularly need ruminant evolutionary partners. Their hooves are designed to open the soils to air and water, and their digestive systems deposit a feast for soil organisms. The ruminant gut is a moist refuge during the dry season for soil microbes which are essential to the health of the land. Grasses need to be bitten lest they shade out their own new growth.
What Savory discovered is that the same land may either flourish or die depending on how it is grazed. When ruminants are kept in check by predators they graze an area in tight herds for protection and then move to the next patch of fresh grasses and other plants, providing up to two years of recovery and regrowth time for the recently grazed pasture. On the other hand, when they are provided the safety of fencing and left to graze large areas at will, they return to their favorite plants and overgraze those areas, eventually compacting the soil, preventing water infiltration and proper aeration, killing the plants, and leading to desertification.
The difference in land health is dramatic.[19] The pictures below illustrate:


Mexico


Arizona


Zimbabwe
Fig. 1. These pictures are of neighboring properties in Mexico, Arizona and Zimbabwe. In each area they were taken on the same day, have similar soils, and the same precipitation. The pictures on the right are examples of properly managed livestock through Holistic Planned Grazing to restore grasslands. On the left we see examples of improperly managed livestock as well as exclusion from grazing (“resting the land”) [Savory Institute 2015:12]
Savory’s work, after decades of successful application on ranches in Africa, Asia, Australia and North and South America, garnered global attention (and controversy) after his TED Talk in 2013 [Savory 2013].
The primary point is that If well-managed, grassland soils can not only sequester annual greenhouse gas emissions but can also begin to draw down legacy atmospheric carbon as well. In addition, they provide human and other predator food, converting grasses inedible to non-ruminant mammals to meat.
Grasslands As Ecosystems
It is useful to understand how grasslands work as intact ecosystems, thereby providing a solid theoretical basis for observations of grasslands as vast carbon and water sinks. What appears above-ground is only a hint of grassland ecosystem dynamics. Soils are the planet’s most complex and least understood terrestrial ecosystem, yet soils are where most of the action takes place on grasslands as well. While all soils are built on the foundation of minerals provided by weathering of rock, the soils on grasslands are primarily biological soils. The kingdoms of life are the active agents in soil creation and it is the interactions among life forms that create the rich and productive grassland soils (see Soils section).
The basis for all life is the microbial kingdom. These smallest of cells, with their complex biochemistry, morphology and behavior, are active players in creating stable soil molecules, storing abundant carbon and water. In addition to microbes, soil ecosystems are built from exchanges among fungi, insects, worms, green plants, birds and small and large mammals. It is this set of rich interactions that creates the biodiverse, abundant and resilient environment of global grasslands.
For the sake of illustration, let us start the discussion with the ruminant gut during a dry season. Microbes survive in a warm, moist environment while constantly being cycled into the soil through digestion and elimination. The short-term hoof disturbance with minimal compaction while animals are constantly moving opens the soils to available moisture from precipitation, urination and condensation. During the rainy season the water is more effectively absorbed into opened soils, nourishing plants, raising the water table and eventually even leading to perennial streams and ponds. Even limited rainfall goes a long way in spongy soils that are covered with grasses and other plants to keep the ground cool and moist [Byck 2014: 8’38”]
Methane
Methane, a relatively short-lived but powerful greenhouse gas, is often raised as a serious concern with beef production. This is surely true when animals are left to roam freely and overgraze, and then moved to concentrated animal feeding operations with large manure lagoons. However, it is important to consider the whole of ecosystem functions in assessing methane emissions {Savory Institute 2015]. This includes the conversion of the methane molecule into carbon dioxide by bacteria (metanotrophs) that live in healthy soils and literally eat energy-rich methane, and the oxidation of methane by hydroxyl radicals present in the lower atmosphere. It may include other ecosystem processes, such as the effects of earthworms increasing methanotrophic bacterial activity in landfills and pastures [Héry 2008; Kernecker 2014]. The result is a virtuous cycle where plants can then take up that carbon dioxide through photosynthesis and send some of the carbon back underground through their root systems. Throughout their life cycle under conventional industrial management, cattle are rarely if ever exposed to such healthy, biodiverse soils.
Historical methane data indicates that in the United States, for example, pre-settlement wild ruminants generated approximately 86% of the methane of current farmed ruminants (Hristov 2012:1371). Yet even with vast numbers of ruminants on grasslands across the planet, atmospheric methane remained constant until the global dependence on widespread use of fossil fuels and its effects on agriculture and animal husbandry began to grow rapidly in the 18th century (Fig. 2).
Thus, results are very different with animals grazed in a manner that mimics nature. Rowntree et al. describe the importance of accounting for the beneficial ecosystem services that well-managed grazing systems can provide.
. . . LCA’s [Life Cycle Assessments] often consider soil C to be in dynamic equilibrium. However, empirical data suggest otherwise (e.g. Machmuller et al., 2015; Teague et al., 2011). Recent studies such as Ripple et al. (2014) and Eshel et al. (2014) have reported the emissions from ruminants in food production without accounting for the beneficial ecosystem services that well-managed grazing systems can provide. In our study, we used 3 tC/ha/yr (1.2 tC/ac/yr) as a potential C sequestration figure, which is relatively high (Conant et al., 2001) but viable based on existing studies (Teague et al., 2011; Delgado et al., 2011; Machmuller et al., 2015; Teague et al., 2016). Importantly, the results presented here suggest that with appropriately managed grazing, a grass-finished beef model can not only contribute to food provisioning but also be ecologically regenerative as well. [Rowntree 2016:36]
This excerpt illustrates a paradigm shift in action, wherein investigators within the dominant paradigm are constrained from evaluating the possibilities offered by “beneficial ecosystem services” because it doesn’t occur to them to consider them. Such biological processes are invisible due to limiting assumptions of the paradigm.
Research into systems implications of holistically managed grass-finished beef is growing. It is only recently that mainstream researchers are beginning to understand that the biological function of animals in an ecosystem is as dependent on the ecosystem as it is on the biology of the animal.[20] Studying animals in isolation or as part of a synthetic system such as industrial agriculture often leads to incorrect conclusions.
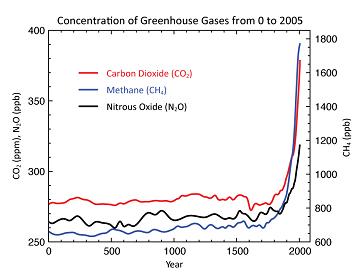
Fig. 2. Historical Concentrations of Greenhouse Gases [IPPC 2007]
Review of some studies of grasslands
Soil creation (pedogenesis) is conventionally defined as the weathering of rock; it may take 3,000 years or longer to create a foot of soil through geological processes. Soil created through biological activity, on the other hand, happens orders of magnitude faster, up to several inches per year.
Australian soil scientist Christine Jones notes that
The rates of soil formation provided in the scientific literature usually refer to the weathering of parent material and the differentiation of soil profiles. These are extremely slow processes, sometimes taking thousands of years. Topsoil formation is different and can occur rapidly under appropriate conditions. . . .
The late P.A. Yeomans, developer of the Keyline system of land management, recognised that the sustainability of the whole farm was dependent on living, vibrant topsoil. The formation of new topsoil using Keyline principles, at rates not previously considered possible, was due to the use of a tillage implement designed to increase soil oxygen and moisture levels, combined with a rest/recovery form of grazing and pasture slashing, to prune grass roots and feed soil biota, especially endemic earthworms. Yeomans was able to produce 10 cm of friable black soil within three years, on what was previously bare weathered red shale on his North Richmond farm (Hill 2002).
Bennett (1939) calculated a rate of topsoil formation of just over 11 t/ha/yr (4.4 t/ac/yr) for soils in which organic material was intermixed into surface layers. In situations where plant root mass is high, rates of topsoil formation of 15-20 t/ha/yr (6-8 t/ha/yr) have been indicated (Brady 1984). Healthy groundcover, high root biomass and high levels of associated microbial activity, are fundamental to the success of any technique for building new topsoil.
If the land management is appropriate, evidence of new topsoil formation can be seen within 12 months, with quite dramatic effects often observed within three years. Many people have built new topsoil in their vegetable or flower gardens. Some have started to build new topsoil on their farms. If you have not seen new soil being formed, make a point of doing so. (Jones 2003:19-20)
Healthy biodiverse grasslands with abundant animal populations provide favorable circumstances for biological soil accumulation and carbon sequestration, including opening soils to air and water, fertilizing soil life and stimulating growth of grasses.

Fig. 2. “Root Systems of Prairie Plants,” Heidi Natura, Conservation Research Institute, n.d., http://kmlandtrust.org/pdf/NPGpp5-6-11×17.pdf
Using current best land management practices, recent research has begun to confirm the importance of studying soil organic carbon accumulation on grasslands. In a 9-year study of bioenergy crops, investigators found that switchgrass and maize stored 50% of their soil organic carbon (SOC) below 30 cm (1 foot), up to 4 times more than used in models in use at that time (Follett 2012:866):
Most of the research on SOC in agricultural production systems focused on C in the 0 to 30 cm depth [22–27]. A few studies in which soil sampling has been conducted at greater depths indicate that production agriculture affects soil C deeper in the soil profile [28,29]. (Follett 2012:867)
Concerns about soil depth measurement are not new [Liebig 2008]; depth of soil measurement in estimating soil carbon storage potential is a significant issue. Conventional soil science, which largely addresses agricultural soils managed in industrial agricultural contexts, typically measures soil carbon down to around 30 – 40 cm (12 – 16 inches). Yet roots of native prairie plants may reach 5 times that depth (see Fig. 1), storing carbon in stable molecules for centuries and millennia as long as the soils are undisturbed and not exposed to light or desiccation. [See Soils section.]
Methodological issues for assessment of SOC have thus been problematic, and have likely led to serious overall underestimation of soil sequestration capacity. This is particularly relevant because these soils will not be able play their appropriate critical role in addressing climate until mainstream science and policy recognize and promote the potential of best practices in land management in all ecosystems. Harper & Tibbett found up to five times more soil carbon in Australian soils at depths greater than 1 meter (~3 feet) than is conventionally estimated:
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths (Table 1), it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5 m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm −2 or 21 % of the total store to bedrock. [Harper 2013: 645]
We discuss the dynamics of water cycling and forests elsewhere, but it is worth noting a recent paper suggesting that a more holistic view of ecosystem dynamics is in order. A paradigm shift prioritizing water over carbon as the driving climate force more accurately and effectively guides climate recovery strategies and offers more tactical and regenerative options [Ellison 2017; s.a. Schmidt 2017]. Water is more tangible to most people than carbon, and water recovery is more visible and rapid, offering hope and encouragement in a generally grim scenario. In addition there are numerous other benefits to people and landscapes with improved water management.
THE IMPORTANCE OF ANECDOTAL EVIDENCE
While anecdotal evidence is often disdained in academic science, in many scientific pursuits it forms the foundation of future study. This is especially true in such predominantly observational pursuits such as naturalist biology, ecology, rangeland science and agronomy, where study of the visible vagaries of the natural world inspires questions that may reach far beyond what a happenstance occurrence would imply. It is the weight of such observations that leads to formal protocols, hypotheses and theories to explore the details of a field. Isolating variables can be a very helpful tool; unfortunately, modern science practice has generally lost sight of systems contexts, and that system behavior can be very different from the behavior of any of its isolated parts.
Therefore, it is essential to embrace both analytical and holistic evidence in order to build a full understanding of how environmental systems work as wholes. Together these complementary approaches provide a more comprehensive picture of the systems in question, as well as much clearer guidance for how to proceed in current global ecosystem urgencies.
A small selection of a growing literature of informative anecdotal reports of grassland eco-restoration is included among formal studies below. They reflect the extensive experience of farmers, ranchers and other land managers, and demonstrate the potential positive effects of regenerative land management and eco-restoration on climate and the biosphere as a whole. See for example Stigge 2016, Oppenheimer 2015, Byck 2014, Brown n.d., Brown 2016.
Grassland Article Summaries
Byck 2014. This 12-minute video relates the experiences of three ranchers who manage cattle and land according to regenerative land management principles. They discuss their transition to Holistic Planned Grazing, where for two of them, in areas of ~15 inches of rainfall, their alternative had been bankruptcy. The video vividly illustrates the benefits of cover-cropping and organics, the improvement in lifestyle and economics, and the dramatic improvement in biodiversity and water management. [Byck 2014]
Follett 2012. A USDA study found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr). Ausmus reports that
. . . a 9-year project that evaluated the effects of nitrogen fertilizer and harvest treatments on soil organic carbon sequestration in switchgrass and no-till maize crops managed for biofeedstock production [found that] more than 50 percent of the soil carbon was found between 1 and 5 feet below the soil surface. The average annual increase of soil organic carbon throughout the first 5 feet of subsoil also exceeded 0.9 tons per acre per year [Ausmus 2014: 4-5].
Of interest were the difficulties the authors faced in getting the study through peer review and published since their results were so unexpected, as reported in Ausmus 2014.[21] It originally appeared in Bioenergy Research in 2012. [Follett 2012].
Note too that the Follett study was performed on already degraded soils, “Perennial grasses could be used as bioenergy crops on about 20 million ha (ha = 10,000 m2 or 2.5 acres) of marginal or idle cropland in the USA alone [18]” (Follett 2012:867). [Emphasis added.] A well-supported inference is that healthy, biodiverse soils will yield even better results.
Degraded soils may be less effective carbon sinks than virgin soils even though they have lost most of their carbon because the soil life that creates long-lasting stable carbon molecules is damaged or destroyed by synthetic inputs, tilling and other forms of mismanagement. Nonetheless, results were dramatic:
In the first 9 years of a long-term C sequestration study in eastern Nebraska, USA, switchgrass and maize with best management practices had average annual increases in SOC per hectare that exceed 2 tC/yr for the 0 to 150 cm soil depth. For both switchgrass and maize, over 50 % of the increase in SOC was below the 30 cm depth. SOC sequestration by switchgrass was twofold to fourfold greater than that used in models to date which also assumed no SOC sequestration by maize. (Follett 2012:866) . . . .
Our results clearly show that significant amounts of C were sequestered deep in the soil profile by switchgrass grown and managed as a biomass energy crop and maize grown continuously in a no-tillage production system for the cultivar Trailblazer array of N fertility and harvest treatments for a 9-year period. For almost all other C sequestration reports used in bioenergy models, studies designed for other purposes were adapted to obtain soil C sequestration estimates, initial soil samples were not available, and control samples were from adjacent fields or non-treatment areas. Our results are supported by similar results reported by Liebig et al. [28] for four switchgrass fields managed with uniform N rates and harvest treatments for 5 years in the USA western Corn Belt. They are supported by the recent work of Varvel and Wilhelm [29] for maize in which significant increases in soil C occurred in soil layers up to 150 cm in depth in maize no-till plots as compared to tilled plots. The soil C that is sequestered deeper than 30 cm is expected to be more stable over time since it is below the tillage zone. Even in the top 30 cm of soil, sequestered C may be stable for extended periods in no-till production systems as reported previously by Follett et al. [33]. (Follett 2012:873)
Harper 2013. This study suggests that the standard sampling depth of 30cm vastly underestimates the global store of soil organic carbon, and thus, presumably, the potential of future soil carbon storage that could result from eco-restoration efforts. “Hypothesizing that SOC retained in soils below the top half metre (in highly weathered deep profiles) would account for the major proportion of SOC in the landscape” [Harper 2013: 642], this southwestern Australia study took deep soil samples in 38 spots across 5 locations. They found that 79% of total carbon store to bedrock occurred below a half meter depth, and 41% occurred below 5 meters depth. “There are two possible sources for the deep carbon; that produced in situ by roots or dissolved carbon that has moved downward from nearer the surface” [Harper 2013: 645]. The sample sites were recently reforested or were under agriculture, and previously had been “covered in a range of xerophytic plants, with root systems that extended to depths of 40 m, such as reported for a Eucalyptus marginata forest” [Harper 2013: 642]. The authors note that more research is needed to understand how deep SOC is affected by land-used changes and climate change.
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths, it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm−2 or 21 % of the total store to bedrock. If this is adjusted to 0.3 m depth, using an exponential function based on the samples in the surface metre, the value decreases to 5.6 kgCm−2. For the individual sites this ranged from 3.6 to 8.0 kgCm−2, or 14 –37 % of the total store. …the surface 5 m contained 16.3±1.38 kgCm−2 or 59 % of the total store to bedrock, with this proportion varying from 47 to 77% across the five sampling locations. The amount of carbon stored in the soils can also be contrasted with the biomass carbon storage of 11.0–16.0kg Cm−2 expected at equilibrium following reforestation for these sites (Harper et al. 2007) and likely previously removed from the sites by deforestation in advance of agriculture [p.645].
Liebig 2008. A study of switchgrass for bioenergy found rates of SOC (Soil Organic Carbon) increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft). “In this study, switchgrass significantly affected change in SOC. . . Across sites, SOC increased significantly at 0–30 cm (1 ft) and 0–120 cm (4 ft), with accrual rates of 1.1 and 2.9 tC/ha (0.44 and 1.16 tC/ac), respectively.” [Liebig 2008:215] This indicates the chronic underestimation of soil carbon capacity in the many studies which by convention only measure SOC down to 30 cm (1 ft).
Machmuller 2015. On intensively grazed former row-crop agricultural land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions. The study “sought to determine how fast and how much soil C accumulates following conversion of row crop agriculture to management-intensive grazed pastures in the southeastern United States. . . . The highest rates of belowground C accumulation occur when land is converted to grassland ecosystems” [Machmuller 2015: 2]. These intensively grazed managed systems led to an approximately 75% increase in soil carbon within six years,
[a] high C accumulation rate [that] stems from year round intensive forage/grazing management techniques on sandy soils with an initially low soil C content due to past conventional-till row crop agriculture. . . . These forage-management techniques are precisely those suggested to increase SOM in pasture systems and when they are applied to soils with degraded SOC content, such as soils in the southeastern United States, rapid C accumulation ensues. . . .
On the basis of a whole farm C-cycle analysis, C accumulation appears to offset methane emissions during the rapid soil C accumulation phase . . . As the C accumulation rate declines these farms will become net C-emitting—similar to all dairy production—because of ruminant methane emissions. However, the substantial soil-quality benefits of higher organic matter remain and will likely increase the sustainability of dairy production using management-intensive grazing [Machmuller 2015:3].
The eventual methane emissions may be markedly less than suggested, however, since the report did not consider methane breakdown from methanotrophic bacteria and atmospheric hydroxyl radical oxidation.
The authors conclude
that pasture-based intensively grazed dairy systems may provide a near-term solution for agricultural lands that have experienced soil-C loss from previous management practices. Emerging land uses, such as management-intensive grazing, offer profitable and sustainable solutions to our needs for pairing food production with soil restoration and C sequestration. [Machmuller 2015: 2-3]
McCosker 2000. A discussion of the introduction of what the author calls “cell grazing,” framed as a paradigm shift in Thomas Kuhn’s terms [Kuhn 1962] over the years 1990-1999. McCosker reviews the dichotomy between researchers and producers, and travels to see actual results in the U.S., Zimbabwe, Namibia, and South Africa. He states, “Only after seeing the outcomes time and again in all possible environments was I finally convinced that the principles could not be faulted.” This kind of paradigm shift has been experienced repeatedly as practitioners must develop the courage to challenge prevailing assumptions in order to take the requisite transitional steps.
Oppenheimer 2015. Farmers Paul and Elizabeth Kaiser own eight acres in Sebastopol in Northern California and they farm three of them, developing a model that other farmers are beginning to use.
[Kaiser] farms a mere eight acres, and harvests fewer than three of them. Nonetheless, his methods are at the forefront of a farming movement that is so new (at least in the U.S.), and so built for a climate-changed world of diminishing rains, that it opens up gargantuan possibilities. One might call this methodology sustainability on steroids, because it can generate substantial profits. Last year, Kaiser’s Sonoma County farm grossed more than $100,000 an acre, which is 10 times the average per-acre income of comparable California farms. This includes Sonoma’s legendary vineyards, which have been overtaking farmland for decades, largely because wine grapes have become much more lucrative these days than food, at least the way most farmers grow it.
Kaiser manages all of this without plowing an inch of his ground, without doing any weeding, and without using any sprays—either chemical or organic. And while most farmers, even on model organic farms, constantly tinker with various fertilizer cocktails, Kaiser concentrates on just one: a pile of rotten food and plants, commonly known as compost, and lots of it. Kaiser then adds this compost to a rare blend of farming practices, both old and new, all aimed at returning dirt to the richest, most fertile seedbed possible.
They use permaculture, agroforestry and other intensive techniques, have built deep, healthy soils by keeping the ground covered and spongy to capture water and carbon, all of which provide solid protection from droughts and floods.
Retallack 2001, 2013. “Grassland expansion initiated increased organic C storage in soils, soil water retention, speed of nutrient exploitation, surface albedo, and C burial in sediments eroded from their soils. These changes had many consequences, including long-term global cooling.” [Retallack 2001:422] and “This climatic zone is not only the most widespread, but also the most fertile region of our planet.” [Retallack 2013:78] The paleohistory of grasslands provides the basis for considering the potential of grasslands as huge biological carbon sinks that may be realized again with regenerative land management
Rodale Institute 2014.
Rodale reports that regenerative grazing practiced on a global scale could sequester 71% of annual emissions of 14 Gt C/yr. Combined results from regenerative grazing and agriculture techniques could, if practiced globally, lead to a net reduction of atmospheric carbon dioxide of 1.7 gigatons per year, or 0.85 ppm/year. This represents a potential sequestration of approx. 3t C/ha/yr (1.2t C/ac/yr) on grasslands and croplands. [Rodale Institute 2014:9] This does not include the considerable contributions of non-agricultural lands, nor recent developments in intensive regenerative practices such as permaculture and biochar.
Moreover, Rodale’s side-by-side trial after 30+ years showed that, following the three-year transition period, organic yields match conventional yields, except in drought years, when organic yields surpass conventional yields. Furthermore, energy input and greenhouse gas emissions were lower in organic systems, and profits were higher.
Rowntree 2016. Examining ruminant methane and net carbon sequestration for grassfed beef in a systems context, Rowntree found net sequestration rates of up to 2.11 tons/ha/yr (0.84 tC/ac/yr) for non-irrigated, lightly stocked grazing.
Therefore, including soil carbon sequestration (SCS) potential brings the differences in grazing environments into focus, and significantly changes the outcome. When comparing two well-managed grazing strategies, grass-finished (MOB) and conventional (IRG), each strategy could be an overall carbon sink, but the MOB grazing would only need sequester half as much carbon (1 tC/ha/yr) as IRG grazing (2 tC/ha/yr) for a net zero greenhouse gas footprint. Methane emissions were similar in both grazing environments, but MOB grazing offered significant benefits in increased carbon sequestration. [Rowntree 2016:36]
It is unlikely that such SCS would take place in the absence of a healthy biodiverse ecosystem, one that is supported through MOB grazing. The result is higher net methane emissions under conventional grazing practices. The opposite occurs with properly managed grazing practices, where grasslands as a system actively build more soil carbon for years, leading to a net increase in soil carbon despite enteric methane production by ruminants.[22]
The recent call for improved management of grazing systems as part of an international climate change mitigation strategy is critical, particularly in light of many existing beef LCAs [Life Cycle Assessments] that have concluded that beef cattle produced in grazing systems are a particularly large sources of GHG emissions. To identify the best opportunities to reduce GHG emissions from beef production, a systems approach that considers the potential to increase soil C and reduce ecosystem-level GHG emissions is essential… [W]e generated an LCA that indicates highly-managed grass-finished beef systems in the Upper Midwestern United States can mitigate GHG emissions through SCS while contributing to food provisioning at stocking rates as high as 2.5 Animal Units (AU) per hectare. From this data, we conclude that well-managed grazing and grass-finishing systems in environmentally appropriate settings can positively contribute to reducing the carbon footprint of beef cattle, while lowering overall atmospheric CO2 concentrations. [Rowntree 2016:36]
Schwartz 2013. Cows Save the Planet was a landmark book that helped launch the regenerative agriculture and eco-restoration climate movements. The author gathers stories from practitioners around the world and paints a picture of broad possibilities for addressing global warming, floods, droughts, desertification, malnutrition and many other seemingly unrelated problems that have a single common cause: human mismanagement of lands across the planet. An excellent introduction for a general readership as well as for scientists unfamiliar with the potentials that nature provides.
Shinn n.d. Ridge Shinn is a rancher in Central Massachusetts (Big Picture Beef) who introduced Allan Savory’s methods to the state. For over ten years he has been investigating the holistic context for bringing healthy meat through the local supply chain, and its relationship to human health and global warming. He states,
Since the 1990’s, science has discovered important connections between rotational grazing, soil health, and healthy food. Big Picture Beef’s methods for raising 100% grass-fed cattle offer huge benefits for the environment and for society. The long term goal of the program is Northeast beef for Northeast markets, carbon sequestration, soil fertility and biodiversity, energy savings, and a revitalized rural economy.
Teague et al. 2016. In a review of the literature, the authors conclude that regenerative conservation cropping and adaptive multi-paddock grazing can turn agricultural soils from a carbon source in conventional agriculture into a carbon sink at rate of ~3 tC/ha/yr (~1.2 tC/ac/yr). Key factors include the use of no-till, cover crops, managed grazing, organic soil amendments and biotic fertilizer formulations. These practices can result in elimination of soil erosion and loss, the greatest agricultural contribution to global warming (1 Gt C/yr). Benefits may include “increased water infiltration, improved water catchment, greater biodiversity, increased ecosystem stability and resilience, and improved C sequestration.” [Teague 2016:158]
Conclusion
These reports demonstrate the promising potential for storage of massive amounts of soil carbon to address both the atmospheric and eco-destruction aspects of climate, along with dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Forests
Note: As mentioned in the Release notes, we have a small staff, and therefore have had to postpone some important material for the next release, scheduled for January 2018. This is particularly true of forests and we will include a more thorough examination of their importance in addressing climate moving forward. Nonetheless, we felt that the investigations here were innovative and interesting, and we wanted to make them available to our readers sooner rather than later.
Forests cover nearly 31% of Earth’s total land area [FAO 2016], and remain one of the major terrestrial ecosystems on the planet. Forests play a significant role in the global ecosystem through cooling, evapotranspiration, covering/shading/sheltering, providing fuel and fiber, aiding cloud formation, and creating wind. Because global forests and wooded lands store an estimated 485 Gt of carbon, forest conservation and afforestation are recognized in the United Nations Framework Convention on Climate Change (UNFCCC) as key strategies for climate change mitigation [UNFCCC 2017].
Despite this acknowledgment, “for the world as a whole, carbon stocks in forest biomass decreased by an estimated 0.22 Gt annually during the period 2011–2015. This was mainly because of a reduction in the global forest area” [UNFCCC 2017]. Indeed, humanity has been in the business of clearing forests for thousands of years, and this continues today. However, rapid reductions in deforestation could abate further carbon emissions and thus extreme results of climate change. Moreover, reductions in deforestation and implementation of agroforestry practices together could restore biodiversity in damaged ecosystems, repair local and global water cycles, and, ultimately, help restore carbon levels to pre-industrial levels. Here we present several articles illustrating the impact of forests on global climate, as well as the potential for restorative afforestation and agroforestry practices to sequester large amounts of carbon.
Forest Article Summaries
Ellison 2017. This paper takes the innovative and paradigm-shifting position that carbon is not the primary consideration in climate; rather, it is water that should be a central focus in assessing climate processes and effects. It considers forests from a systems perspective.
Forest-driven water and energy cycles are poorly integrated into regional, national, continental and global decision-making on climate change adaptation, mitigation, land use and water management. This constrains humanity’s ability to protect our planet’s climate and life-sustaining functions. The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. The effects of tree cover on climate at local, regional and continental scales offer benefits that demand wider recognition. The forest- and tree-centered research insights we review and analyze provide a knowledge-base for improving plans, policies and actions. Our understanding of how trees and forests influence water, energy and carbon cycles has important implications, both for the structure of planning, management and governance institutions, as well as for how trees and forests might be used to improve sustainability, adaptation and mitigation efforts. [Ellison 2017: Abstract]
Ford 2017. Structural Complexity Enhancement (SCE) is part of a larger ecological concept: nature tends to complexity, providing its resiliency, flexibility and inventiveness. SCE in treatment of forests is a management approach that promotes development of late-successional structure, including elevated levels of coarse woody debris. It adds variety to tree ages (favoring older trees), and variations in available sunlight and habitat.
Large trees, previously assumed to slow in both productivity and growth rate (Weiner and Thomas 2001, Meinzer et al. 2011), function as long-term carbon sinks (Carey et al. 2001). These findings further support the significance of structural retention as a co-benefit to forest carbon storage. Adaptive silvicultural practices promoting multiple co-benefits, for instance, by integrating carbon with production of harvestable commodities, can contribute to efforts to dampen the intensity of future climate change while maintaining resilient ecosystems (Millar et al. 2007). Prescriptions that enhance in situ forest biomass and thus carbon storage offer one such alternative (Ducey et al. 2013). U.S. forests currently offset approximately 16% of the nation’s anthropogenic CO2 emissions, but this has the potential to decline as a result of land-use conversion and lack of management (EPA 2012, Joyce et al. 2014). While passive or low-intensity management options have been found to yield the greatest carbon storage benefit, assuming no inclusion of substitution effects (Nunery and Keeton 2010) or elevated disturbance risks (Hurteau et al. 2016), we suggest the consideration of SCE to enhance carbon storage. Multiple studies have explored co-benefits provided by management for or retention of elements of stand structural complexity, including residual large living and dead trees, horizontal variability, and downed CWM (Angers et al. 2005, Schwartz et al. 2005, Dyer et al. 2010, Gronewold et al. 2012, Chen et al. 2015). Silvicultural treatments can effectively integrate both carbon and late-successional biodiversity objectives through SCE based on this study and previous research (e.g., Dove and Keeton 2015). Remaining cognizant of the potential for old-growth compositional and structural baselines to shift over time and space with global change—climate impacts on forest growth and disturbance regimes, altered species ranges, and the effects of invasive species—will be important for adaptive management for late-successional functions such as carbon storage. [Ford 2017: 16]
Healing Harvest Forest Foundation.
The spot compaction of animal feet is far less damaging to the forest soil and tree roots than the continuous track created by a wheel or track driven machine. Small sized tracts of timber can not be harvested with conventional methods that require higher capitalization and expensive moving cost. The economic pressure in conventional forest harvesting operations influences most loggers to feel that they must cut all the trees to make their work cost effective. This restricts the silvicultural prescriptions available for the management of the forest….Our method of selecting individual trees on a “worst first” basis and limiting removal to no more than 30% retains the forested condition and is indeed improvement forestry…. The holes created in the forest canopy are substantial enough for “shade intolerant” species to regenerate naturally from seedlings of the superior specimens that are left in a healthy “good growing” condition. We believe that basically the repair of the forest from previous “high grading” is best accomplished through several successive “low grading” harvests. [Healing Harvest 1999]
Makarieva 2007. The authors examine ecological and geophysical principles to explain how land far inland away from the ocean can remain moist, given that gravity continuously pulls surface and groundwater into the ocean over time.
All freshwater on land originates in the ocean from which it has evaporated, is carried on air flux, and precipitates over the land. Coastal regions benefit from this cycle by their proximity to the ocean, yet in the absence of natural forests in coastal regions precipitation weakens as distance from the ocean increases, leaving inland areas arid. The authors propose the concept of a biotic pump to explain how large continents can be sufficiently moist deep into the interior and abundant with rivers and lakes.
Air and moisture are pulled horizontally by evapotranspiration from coastal forests. When water vapor from plants condenses, it creates a partial vacuum which pulls water evaporating from the ocean into the continental interior where it rains in forest. By contrast, deserts are unable to pull ocean evaporation to them because they lack any evaporative force.
Such ongoing deforestation, and crucially coastal deforestation on a large scale, threatens to cut off rain to the interiors of Earth’s continents thereby creating new deserts. The Amazonian rainforest is the prime example. Deforestation of the eastern coast of South America has led to changes in the rainforest that is resulting in drying and desertification of the interior, with unprecedented fires and loss of rivers. Historically, Australia’s interior became a desert around the time the first humans arrived on the continent, and the authors speculate that early coastal deforestation was the cause. On the other hand, restoring natural coastal forests can also restore inland water cycles and reverse desertification.
This article illustrates the importance of biological relationships that are ecologically complex and poorly understood. It highlights the significance of the precautionary principle in assessing what we don’t know (and what we don’t know that we don’t know) when altering ecological processes, and taking preventive action in the face of uncertainty.
Asner, Gregory P., Jonathan M.O. Scurlock, Jeffrey A. Hicke, Global synthesis of leaf area index observations: implications for ecological and remote sensing studies, Global Ecology & Biogeography (2003) 12 , 191– 205, Blackwell Publishing Ltd., http://www2.geog.ucl.ac.uk/~mdisney/teaching/teachingNEW/GMES/LAI_GLOBAL_RS.pdf. [Do we have more soil . . . ]
Ausmus, Steven, A Surprising Supply of Deep Soil Carbon, Agricultural Research, USDA, February 2014, 4-6, https://agresearchmag.ars.usda.gov/ar/archive/2014/feb/february2014.pdf (s.a. Follett 2012). [Grasslands]
Balfour, Lady Eve 1977, http://journeytoforever.org/farm_library/balfour_sustag.html [Earthworms].
Balfour, Lady Eve, Milton 1975, The Living Soil and the Haughley Experiment, 2nd revised edition, Faber & Faber, London, http://organiccollege.com/dl/file.php/1/the-living-soil.pdf [Earthworms].
Blakemore, R. and Axel Hochkirch, 2017, Soil: Restore earthworms to rebuild topsoil, Nature 545, 30 (04 May 2017), http://www.nature.com/nature/journal/v545/n7652/full/545030b.html [Earthworms]
Blakemore, R.J. 2000, Ecology of earthworms under the ‘Haughley experiment’ of organic and conventional management regimes. Biological Agriculture and Horticulture, 18:141—159, www.annelida.net/earthworm/Haughley/Haughley.doc. [Soils, Grasslands]
Blakemore, R.J. 2012, Call for a Census of Soil Invertebrates (CoSI), Zoology in the Middle East, 58: Supplementum 4: 163-176,
https://vermecology.wordpress.com/2017/04/30/census-of-soil-invertebrated-cosi/. [Soils, Earthworms]
Blakemore, R.J. 2015, Eco-taxonomic profile of iconic vermicomposter – the ‘African Nightcrawler’ earthworm, Eudrilus eugeniae (Kinberg, 1867), African Invertebrates, 56(3): 527-548, http://africaninvertebrates.org/ojs/index.php/AI/article/view/395]. [Earthworms]
Blakemore, R.J. 2016a, Veni, Vidi, Vermi – I. On the contribution of Darwin’s ‘humble earthworm’ to soil health, pollution-free primary production, organic ‘waste’ management & atmospheric carbon capture for a safe and sustainable global climate, July, 2016, VermEcology Occasional Papers, 2(1): 1-34, https://veop.files.wordpress.com/2016/09/vvv-i.pdf. [Soils, Grasslands]
Blakemore, R.J. 2016b, Veni, Vidi, Vermi – II. Earthworms in organic fields restore SOM & H2O and fix CO2, July, 2016, VermEcology Occasional Papers, 2(2): 1-26, https://veop.files.wordpress.com/2016/09/vvv-ii.pdf. [Soils, Grasslands]
Blakemore, R.J. 2016c, Cosmopolitan Earthworms - An Eco-Taxonomic Guide to the Peregrine Species of the World, 6th ed., VermEcology, Yokohama 2016 https://vermecology.wordpress.com/2016/07/20/cosmopolitan-earthworms-6th-edition-2016-is-now-available/. [Soils]
Blakemore, Robert J. 2017a, Food for Thought (Part II) – Like sands of the hourglass, so are the days of our soils (with apologies to Socrates), February 22, 2017, https://vermecology.wordpress.com/2017/02/22/food-for-thought-ii/. [Soils]
Brady, Nyle C., Raymond Weil 2008, The Nature and Properties of Soils, 14th Edition, https://www.pearson.com/us/higher-education/product/Brady-Nature-and-Properties-of-Soils-The-14th-Edition/9780132279383.html [Soils]
Bryce, Emma 2013, Do the UN's new numbers for livestock emissions kill the argument for vegetarianism?, The Guardian, 27 September 2013, https://www.theguardian.com/environment/world-on-a-plate/2013/sep/27/environment-food-ipcc-emissions-greenhouse-gas-livestock-vegetarian-meat [Earthworms]
Byck, Peter, 2014, Soil Carbon Cowboys, https://vimeo.com/80518559. [Grasslands]
Comis, Don 2002, Glomalin: Hiding Place for a Third of the World’s Stored Soil Carbon, Agricultural Research, 4-7, September 2002, https://agresearchmag.ars.usda.gov/AR/archive/2002/Sep/soil0902.pdf [Soils]
Crowther, Thomas, et al., 2016, Quantifying global soil carbon losses in response to warming. Nature (Letter), 540, 104–108, https://www.nature.com/nature/journal/v540/n7631/full/nature20150.html [Soils]
Darwin, Charles, R. 1881, The Formation of Vegetable Mould through the Action of Worms with Observations on their Habits, London: John Murray, http://charles-darwin.classic-literature.co.uk/formation-of-vegetable-mould/ebook-page-49.asp
[Soils, Earthworms]
DuPont, S. et al., 2010. No-tillage conversion of harvested perennial grassland to annual cropland reduces root biomass, decreases active carbon stocks, and impacts soil biota, Agriculture, Ecosystems and Environment 137, 25-32. [Croplands]
Duarte, Carlos, Marianne Holmer, Yngvar Olsen 2009, Will the Oceans Help Feed Humanity? Bioscience, December 2009, Vol. 59 No. 11, pp 967-976, https://academic.oup.com/bioscience/article/59/11/967/251334/Will-the-Oceans-Help-Feed-Humanity [Soils]
Eagle, Alison, et al. 2012. Greenhouse Gas Mitigation Potential of Agricultural Land Management in the United States A Synthesis of the Literature, Nicholas Institute for Environmental Policy Solutions Report NI R 10-04 3rd Edition. [Croplands]
Ellison, David, et al., Trees, forests and water: Cool insights for a hot world, Global Environmental Change 43, January 2017, 51–61, http://www.sciencedirect.com/science/article/pii/S0959378017300134. [Grasslands, Forests, Water]
FAO 2011, The state of the world’s land and water resources for food and agriculture: managing systems at risk, http://www.fao.org/docrep/017/i1688e/i1688e.pdf. [Croplands]
FAO 2016, Global forest resources assessment 2015: How are the world’s forests changing? Food and Agriculture Organization of the United Nations, 2nd ed., http://www.fao.org/3/a-i4793e.pdf. [Forests]
FAO 2016, Livestock’s Long Shadow http://www.fao.org/newsroom/en/news/2006/1000448/index.html [Earthworms]
FAO 2017a, Soil Biodiversity, Soil Conservation and Agriculture http://www.fao.org/soils-portal/soil-biodiversity/soil-conservation-and-agriculture/en/. [Soils, Earthworms]
FAO 2017b, Soil Organic Carbon: the hidden potential, Food and Agriculture Organization of the United Nations, Rome, Italy, http://www.fao.org/documents/card/en/c/ed16dbf7-b777-4d07-8790-798604fd490a/. [Croplands]
Foley, Jonathan A. et al., 2011, Solutions for a cultivated planet, Nature 478, 337–342, http://tempag.net/documents/solutions-for-a-cultivated-planet-foley-et-al-2011-nature/. [Croplands]
Follett, Ronald F., Kenneth P. Vogel et al. 2012, Soil Carbon Sequestration by Switchgrass and No-Till Maize Grown for Bioenergy, Bioenerg. Res. May 4, 2012, 5:866–875, http://link.springer.com/article/10.1007/s12155-012-9198-y (s.a. Ausmus 2014). [Grasslands, Croplands]
Ford, S., W. Keeton, 2017, Enhanced carbon storage through management for old-growth characteristics in northern hardwood-conifer forests, Ecosphere, Vol. 8: 4, http://onlinelibrary.wiley.com/doi/10.1002/ecs2.1721/full. [Forests]
Gerber, P.J., Hemming Steinfeld, Benjamin Henderson, et al. 2013, Tackling climate change through livestock - A global assessment of emissions and mitigation opportunities, Food and Agriculture Organization of the United Nations (FAO) 2013, http://www.fao.org/ag/againfo/resources/en/publications/tackling_climate_change/index.htm. [Earthworms]
Glover, Jerry, Cindy M. Cox, John P. Reganold, 2007, Future agriculture: a return to the roots? Scientific American, August 2007, https://www.scientificamerican.com/article/future-farming-a-return-to-roots/. [Croplands]
Harper R.J., M. Tibbett 2013, The hidden organic carbon in deep mineral soils, Plant and Soil 368 (1-2): 641-648, http://link.springer.com/article/10.1007%2Fs11104-013-1600-9. [Grasslands]
Healing Harvest Forest Foundation, 1999, Restorative forestry and modern horselogging.
http://www.healingharvestforestfoundation.org/restorative-forestry-and-modern-horselogging.html. [Forests]
Houghton, R. A. 2010, "How well do we know the flux of CO2 from land-use change?," Tellus (2010), 62B, 337–351, http://onlinelibrary.wiley.com/doi/10.1111/j.1600-0889.2010.00473.x/epdf
Howard, Albert 1945a, Sir Albert Howard on Earthworms: Introduction to The Formation of Vegetable Mould through the Action of Worms with Observations on their Habits by Charles Darwin, John Murray, London, 1881, Faber and Faber, London, 1945, http://journeytoforever.org/farm_library/oliver/howard_intro.html [Soils, Earthworms]
Howard, Albert 1945b, Miscellaneous papers presented online by Soil & Health, http://soilandhealth.org/wp-content/uploads/01aglibrary/010142howard.misc/010140.ogf.1945-47.htm [Soils]
Héry, Marina, Andrew C Singer, Deepak Kumaresan, et al. 2008, Effect of earthworms on the community structure of active methanotrophic bacteria in a landfill cover soil, International Society for Microbial Ecology 2, 92–104, https://www.ncbi.nlm.nih.gov/pubmed/18049457.
IPCC, 2013, Adoption and acceptance of the 2013 supplement to the 2006 guidelines: wetlands, http://www.ipcc.ch/meetings/session37/Doc_8b_Rev_2_Accepted_Report_Wetlands.pdf. [Soils]
Jackson, R.B., H.A. Mooney, E-D. Schulze 1997, A global budget for fine root biomass, surface area, and nutrient contents, Proc. Natl. Acad. Sci. USA, Vol. 94, pp. 7362–7366, July 1997 Ecology, https://jacksonlab.stanford.edu/sites/default/files/pnas97.pdf [Soils]
Jardine C.N., B. Boardman, A. Osman et al, 2009, Climate science of methane, in Jardine et al. (eds), Methane, Environmental Change Institute, University of Oxford, 14-23, http://www.eci.ox.ac.uk/research/energy/downloads/methaneuk/chapter02.pdf. [Wetlands]
Johnson, David 2017, Soils Beneath Our Feet: Can Regenerative Agriculture and Healthy Soils Help Combat Climate Change, https://www.youtube.com/watch?v=XlB4QSEMzdg. [Soils]
Kaiser, Paul and Elizabeth 2017, No-till Farmers Elizabeth and Paul Kaiser Keynote 2017 NOFA/Mass Winter Conference, https://www.youtube.com/watch?v=zAn5YxL1PbM. [Croplands]
Kallenbach, Cynthia, Serita D. Frey, A. Stuart Grandy 2016, Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls, Nature Communications 7:13630, https://www.nature.com/articles/ncomms13630 [Soils]
Kernecker, Maria, Joann K. Whalen, Robert L. Bradley 2014, Endogeic earthworms lower net methane production in saturated riparian soils, Biology and Fertility of Soils 50 (7), https://link.springer.com/article/10.1007/s00374-014-0965-0. [Grasslands]
Khan, S.A. et al. 2007, The Myth of Nitrogen Fertilization for Soil Carbon Sequestration, Journal of Environmental Quality, 36, https://www.ncbi.nlm.nih.gov/pubmed/17965385.
Khursheed S, Cheryl Simmons, Fouad Jaber 2016, “Glomalin” a Key to Locking Up Soil Carbon, Adv. Plants Agric. Res. 4(1): 00126, http://medcraveonline.com/APAR/APAR-04-00126.php [Soils]
Kravčík, Michal, Jan Pokorný, Juraj Kohutiar, et al., 2007, Water for the Recovery of the Climate: A New Water Paradigm, http://www.waterparadigm.org/download/Water_for_the_Recovery_of_the_Climate_A_New_Water_Paradigm.pdf [Water]
Kuhn, Thomas 1962, The Structure of Scientific Revolutions, U. Chicago Press, 1962; full text available (2d edition, 1970) at http://projektintegracija.pravo.hr/_download/repository/Kuhn_Structure_of_Scientific_Revolutions.pdf. [Introduction]
Lal R. 2008, Carbon sequestration, Philosophical Transactions of the Royal Society, 363: 815-830, http://rstb.royalsocietypublishing.org/content/royptb/363/1492/815.full.pdf. [Wetlands]
Lal, R., J. Kimble, E. Levine, B. Stewart 1995, Soils and Global Change, CRC Press. [Wetlands]
Lal, Rattan, 2016, Beyond COP21: Potential and challenges of the “4 per Thousand” initiative, Journal of Soil and Water Conservation 71(1), http://www.jswconline.org/content/71/1/20A.extract [Soils]
Lee, K.E., 1985, Earthworms, their ecology and relationships with soils and land use, Academic Press, New York, NY. [Soils, Earthworms]
Liebig, M.A., et al., Soil Carbon Storage by Switchgrass Grown for Bioenergy, Bioenerg. Res. (2008) 1:215–222, http://naldc.nal.usda.gov/download/28132/PDF [Grasslands, Croplands]
Machmuller, Megan B., et al., 2015, Emerging land use practices rapidly increase soil organic matter, Nature Communications, April 30, 2015, p. 1-5, http://www.nature.com/articles/ncomms7995. [Grasslands]
Magdoff, Fred, Harold Van Es 2009, Building Soils for Better Crops: sustainable soil management, Sustainable Agriculture Research and Education (SARE), http://www.sare.org/Learning-Center/Books/Building-Soils-for-Better-Crops-3rd-Edition [Soils]
Mangalassery, S., S. Sjogersten, D.L. Sparkes, S.J. Mooney 2015, Crops and soils review: Examining the potential for climate change mitigation from zero tillage, Journal of Agricultural Science, https://www.cambridge.org/core/journals/journal-of-agricultural-science/article/examining-the-potential-for-climate-change-mitigation-from-zero-tillage/7248675D2F3E3FFEBB3DFB2379790E6C. [Croplands]
McLaughlin, Hugh 2017, Biochar’s Role in the Landscape, Landscape Heros conference, January 31, 2017, University of Massachusetts, Amherst https://www.youtube.com/watch?v=Z0tTfJc1kNk&list=PLsWWRqCX9eSZBwrUokqGqz444f90ilK-X&index=8. [Biochar]
Minasny, Budiman et al. 2017. Soil carbon 4 per mille, Geoderma 292 (59-86), http://www.sciencedirect.com/science/article/pii/S0016706117300095. [Croplands] }
Mitra, S., R. Wassmann, P.L.G. Vlek 2005, An appraisal of global wetland area and its organic carbon stock, Current Science, 88:1, 10 January 2005, http://www.iisc.ernet.in/currsci/jan102005/25.pdf. [Wetlands]
Mitsch, W.J., B. Bernal, A.M. Nahlik, et al. 2012, Wetlands, carbon, and climate change, Landscape Ecology, 28:583, http://link.springer.com/article/10.1007/s10980-012-9758-8. [Wetlands]
Mitsch, W.J., J.G. Gosselink 2007, Wetlands, 4th ed., Wiley, Hoboken.
Mollison B.C. 1988, Permaculture: A Designers’ Manual, Tagari Publications, Tasmania. [Soils, Worms]
Montgomery, D. 2008, Dirt: The Erosion of Civilizations, UC Press, Berkeley, https://books.google.co.jp/books?id=D2im0qYGG2YC&pg=PA51&lpg=PA51&dq=aristotle+erosion+soil+athens+plato&source=bl&ots=txSi8MM2pb&sig=KlTF6TWmsGpKcT57D5mDckyGLkc&hl=en&sa=X&ved=0ahUKEwi_hJ2r0ZvUAhVDKZQKHanjDZwQ6AEIIjAA#v=onepage&q=aristotle%20erosion%20soi&f=false [Soils]
Mulvaney, R.L., S. A. Khan, T. R. Ellsworth 2009, Synthetic Nitrogen Fertilizers Deplete Soil Nitrogen: A Global Dilemma for Sustainable Cereal Production, Journal of Environmental Quality, Vol. 38 (November–December 2009), https://www.ncbi.nlm.nih.gov/pubmed/19875786.
NASA 2017, Satellite data confirm annual carbon dioxide minimum above 400 ppm, NASA Global Climate Change News, January 30, 2017, https://climate.nasa.gov/news/2535/satellite-data-confirm-annual-carbon-dioxide-minimum-above-400-ppm/ [Soils]
NASA 2017, Shuttle Radar Topography Mission, Jet Propulsion Laboratory, California Institute of Technology, https://www2.jpl.nasa.gov/srtm/coverage.html [Soils]
NASA Earth Observatory 2011, The Carbon Cycle, http://earthobservatory.nasa.gov/Features/CarbonCycle/. [Soils, Grasslands]
Nahlik, A.M., M.S. Fennessy 2016, Carbon storage in US wetlands, Nature Communications, December 13, 2016, https://www.nature.com/articles/ncomms13835. [Wetlands]
Nair, P. K. Ramachandran, B. Mohan Kumar, Vimala D. Nair 2009, Agroforestry as a strategy for carbon sequestration, Journal of Plant Nutrition Soil Science, 172, 10–23, http://onlinelibrary.wiley.com/doi/10.1002/jpln.200800030/abstract. [Croplands]
Odum, E. P., Gary W. Barrett 2005, Fundamentals of Ecology, 5th Ed., Thomson, http://tocs.ulb.tu-darmstadt.de/129640778.pdf [Earthworms]
Oppenheimer, Todd, The Drought Fighter (Paul Kaiser), Craftsmanship Quarterly, Jan 15, 2015, https://craftsmanship.net/drought-fighters/. See also Paul and Elizabeth Kaiser, Singing Frogs Farm website, http://www.singingfrogsfarm.com/our-farming-model.html. [Grasslands, Croplands ]
Paustian, Keith, Johannes Lehmann, et al. 2016, Climate-smart soils, Nature, 532: 49-57, http://www.css.cornell.edu/faculty/lehmann/publ/Nature%20532,%2049-57,%202016%20Paustian.pdf. [Biochar, Soils]
Paustian, Keith. et al. 1997, Agricultural soils as a sink to mitigate CO2 emissions, Soil Use and Management, 13 (230-244), http://onlinelibrary.wiley.com/doi/10.1111/j.1475-2743.1997.tb00594.x/abstract. [Croplands]
Pimentel, David, Michael Burgess 2013, Soil erosion threatens food production, Agriculture, 3(3): 443-463, http://www.mdpi.com/2077-0472/3/3/443 [Soils, Worms]
Pimentel, David. 2011. Food for Thought: A Review of the Role of Energy in Current and Evolving Agriculture, Critical Reviews in Plant Sciences, 30, Special Issue: Towards a More Sustainable Agriculture, http://www.tandfonline.com/doi/abs/10.1080/07352689.2011.554349?journalCode=bpts20.
[Croplands]
Poeplau C., Axel Don 2015, Carbon sequestration in agricultural soils via cultivation of cover crops – A meta-analysis, Agriculture, Ecosystems and Environment, 200, http://www.sciencedirect.com/science/article/pii/S0167880914004873. [Croplands]
Remediation Magazine 2017, Biochar’s long-term benefits to soil proven, May 30, 2017 http://www.remmag.com/2017/biochars-long-term-benefits-to-soil-proven/
Retallack, Gregory J. 2001, Cenozoic Expansion of Grasslands and Climatic Cooling, The Journal of Geology, 2001, 109 (407–426), http://www.journals.uchicago.edu/doi/abs/10.1086/320791?journalCode=jg. [Grasslands]
Retallack, Gregory J., 2013, Global Cooling by Grassland Soils of the Geological Past and Near Future, Annu. Rev. Earth Planet. Sci. 41:69–86, http://www.annualreviews.org/doi/abs/10.1146/annurev-earth-050212-124001. [Grasslands]
Rockström J., W. Steffen, H. Schellnhuber, R. Costanza, et al. 2009, A safe operating space for humanity, Nature, 461: 472–475, http://steadystate.org/wp-content/uploads/2009/12/Rockstrom_Nature_Boundaries.pdf. [Do we have more soils . . . ]
Rodale Institute White Paper 2014, Regenerative Organic Agriculture and Climate Change: A Down-to-Earth Solution to Global Warming, https://rodaleinstitute.org/regenerative-organic-agriculture-and-climate-change/ [Soils, Grasslands]
Rowntree, Jason, et al., Potential mitigation of midwest grass-finished beef production emissions with soil carbon sequestration in the United States of America, Future of Food: Journal on Food, Agriculture and Society 4 (3) Winter 2016, http://futureoffoodjournal.org/index.php/journal/article/view/232 [Grasslands]
Ryals, Rebecca, Whendee Silver 2013, Effects of organic matter amendments on net primary productivity and greenhouse gas emissions in annual grasslands, Ecological Applications, 23(1), http://www.c-agg.org/cm_vault/files/docs/38/ryals_and_silver_ecoapps2013.pdf
Savory Institute 2016, Restoring the Climate through Capture and Storage of Soil Carbon through Holistic Planned Grazing, http://savory.global/assets/docs/evidence-papers/RestoringClimateWhitePaper2015.pdf [Grasslands]
Savory Institute, An Exploration of Methane and Properly Managed Livestock through
Holistic Management, 2015, http://savory.global/assets/docs/evidence-papers/exploration-of-methane.pdf [Grasslands]
Savory, Allan, How to fight desertification and reverse climate change, TED Talk, February 2013, https://www.ted.com/talks/allan_savory_how_to_green_the_world_s_deserts_and_reverse_climate_change [Grasslands]
Schmidt, Michael W.I, Margaret Torn, Samuel Abiven, et al. 2011, Persistence of soil organic matter as an ecosystem property, Nature, October 6, 2011, 478: 49-56, http://www.nature.com/nature/journal/v478/n7367/full/nature10386.html [Grasslands, Soils]
Singh, Rajendra, 2007, Community Driven Approach for Artificial Recharge –TBS Experience, Tarun Bharat Sangh, Tarun Ashram, Bhikampura, District Alwar, Rajasthan, India, http://www.cgwb.gov.in/documents/papers/incidpapers/Paper%205%20-%20Rajendra%20Singh.pdf. See also, An ancient technology is helping India’s “water man” save thousands of parched villages, https://qz.com/367875/an-ancient-technology-is-helping-indias-water-man-save-thousands-of-parched-villages/; and Rajendra Singh - The water man of India wins 2015 Stockholm Water Prize, http://www.siwi.org/prizes/stockholmwaterprize/laureates/2015-2/. [Water]
Smith, Pete et al. 2007, Agriculture, in B. Metz, O.R. Davidson, P.R. Bosch, et al., eds., Climate Change 2007: Mitigation, Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, http://www.ipcc.ch/publications_and_data/ar4/wg3/en/ch8.html. [Croplands]
Smith, Steven, et al. 2015. Near-term acceleration in the rate of temperature change, Nature Climate Change 5, http://www.nature.com/nclimate/journal/v5/n4/full/nclimate2552.html. [Soils]
Suttie, J.M., S.G. Reynolds, C Batello, eds. 2005, Grasslands of the World. Food and Agriculture Organization of the United Nations, Ch. 1, http://www.fao.org/docrep/008/y8344e/y8344e05.htm - bm05. [Grasslands]
Taylor, Paul, ed. 2010, The Biochar Revolution: Transforming Agriculture and Environment, Global Publishing Group, http://globalpublishinggroup.com.au/authors/paul-taylor/the-biochar-revolution/. [Croplands]
Teague, W.R., Steve Apfelbaum, Rattan Lal et al. 2016, The role of ruminants in reducing agriculture’s carbon footprint in North America, J. Soil and Water Conservation, March/April 2016, 71:2, http://www.jswconline.org/content/71/2/156.abstract. [Grasslands, Croplands]
UNFCCC 2017 (United Nations Framework Convention on Climate Change) 2017, Land Use, Land-Use Change and Forestry (LULUCF), http://unfccc.int/land_use_and_climate_change/lulucf/items/1084.php, [Forests]
Udawatta, Ranjith. P., Jose, Shibu 2011, Carbon Sequestration Potential of Agroforestry Practices in Temperate North America, Carbon Sequestration Potential of Agroforestry Systems: Opportunities and Challenges, Advances in Agroforestry 8, http://www.springer.com/us/book/9789400716292. [Croplands]
Vepraskas, M.J., C.B. Craft 2016, Wetland Soils, 2nd ed., CRC Press. [Wetlands]
Vick, Elizabeth S.K., Paul C. Stoy, et al., The surface-atmosphere exchange of carbon dioxide, water, and sensible heat across a dryland wheat-fallow rotation,” Agriculture, Ecosystems and Environment 232 (2016) 129–140, http://www.sciencedirect.com/science/article/pii/S0167880916303863. [Grasslands, Croplands]
Wall, D. 2014, What Can Soil Creatures Say About Climate Change, American Association for the Advancement of Science, February 16, 2014, https://www.aaas.org/news/diana-wall-what-can-soil-creatures-say-about-climate-change [Soils]
Weng, Zhe (Han), Lukas Van Zwieten 2017, Biochar built soil carbon over a decade by stabilizing rhizodeposits, Nature Climate Change 7, 371–376, doi:10.1038/nclimate3276, https://www.nature.com/nclimate/journal/v7/n5/full/nclimate3276.html [Soils]
West, Tristram O., Wilfred M. Post 2002, Soil Organic Carbon Sequestration Rates by Tillage and Crop Rotation: A Global Data Analysis, Soil Science Society of America Journal 66:1930–1946, https://vtechworks.lib.vt.edu/handle/10919/69175. [Croplands]
White, R., S. Murray, M. Rohweder 2000, Pilot analysis of global ecosystems: Grassland ecosystems, World Resources Institute, Washington D.C., http://www.wri.org/publication/pilot-analysis-global-ecosystems-grassland-ecosystems.
[Croplands, Grasslands]
[5] Arbuscular mychorrhizal fungi form a relationship in which they penetrate the cortical roots of green plants for the purpose of mutual exchange of nutrients.
[6] Soils with organic horizon at least 10cm thick and, if less than 20 cm, then containing at least 12% carbon when mixed to a depth of 20cm [IPCC 2013].
[7] Cation exchange capacity represents the ability of the soil to hold nutrients.
[8] It is worth noting that at present oceans only contribute 2% of the world’s food supply, the remainder harvested from terrestrial ecosystems, primarily soils. [Duarte 2009]
[9] The Natural Resources Conservation Service, a division of the U.S. Department of Agriculture.
[10] Whereby an organism produces one or more biochemicals that influences the germination, growth, survival, and reproduction of other organisms.
[11] Earthworm scientist.
[12] The global surface area of earth is about 51 Gha. According to conventional calculations, about 71 percent (~36.2 Gha) is covered by water and 29 percent (14.8-15.1 Gha) is land [Space.com n.d.] with a mean estimate of ~15 Gha land. This land, excluding that in “extreme desert, rock, sand, ice, swamps, marshes, lakes, and streams” (19%), has an area available for photosynthetic activity estimated at 81% [Jackson 1997: table 2]. Therefore ~12 Gha represents land covered with soils, our primary concern. Coincidentally this corresponds to a flat area (~12 Gha) scanned by the SRTM (Shuttle Radar Topography Mission) satellite, excluding Antarctica and the Arctic [NASA 2017].
[14] Small-scale soil activity, such as earthworm burrows or casts, plays a significant role in carbon cycle dynamics [Darwin 1881]. Moreover, “The total fine-root C pool is 5% of the size of the atmospheric C pool, and the biomass of living fine roots is similar in magnitude to 33% of annual net primary productivity. The small diameter, relatively short lifespan, and low C:N ratio of fine roots result in their rapid turnover and quick decomposition compared with woody biomass, making them disproportionately important for estimates of annual net primary production, nutrient cycling, and carbon allocation” [Jackson 1997: 7365].
[15] Approximately 75 billion tons are lost from agricultural soils per year, with 32 million seconds per year, this equals approximately 2,378 t/sec - see https://vermecology.wordpress.com/2017/02/22/food-for-thought-ii/.
[16] Amundson 2015: 647
[17] Organic farming systems favor “greater diversity in the biological community, fewer annuals and more perennials, and greater reliance on internal rather than external resources”) [Rodale 2014: 7]
[18]C3 and C4 refer to two different photosynthetic pathways among flowering plants. C4 (warm-season) plants photosynthesize more efficiently and have higher water-use efficiency than do C3 (cool-season) plants; C4 plants are well-adapted to hotter, drier environments, while C3 plants thrive in cooler environments.
[19] For more examples of land comparisons under holistic management, see http://sheldonfrith.com/2016/02/06/holistic-management-comparison-pictures-infographic/.
[20] For another example of the dependency of parts of a system on the function of the system as a whole, see Schmidt 2011, Persistence of soil organic matter as an ecosystem property.
[21] See section on paradigms for discussion of the difficulties in shifting assumptions.
[22] In addition, as already noted, the powerful action of methane-decomposing bacteria (methanotrophs) found on natural pastures is often ignored, leading to an over-estimation of methane concentrations due to grazing on healthy, biodiverse grasslands.
[23] “IMRaD” stands for Introduction, Methods, Research and Discussion, the basic structure of most scientific
