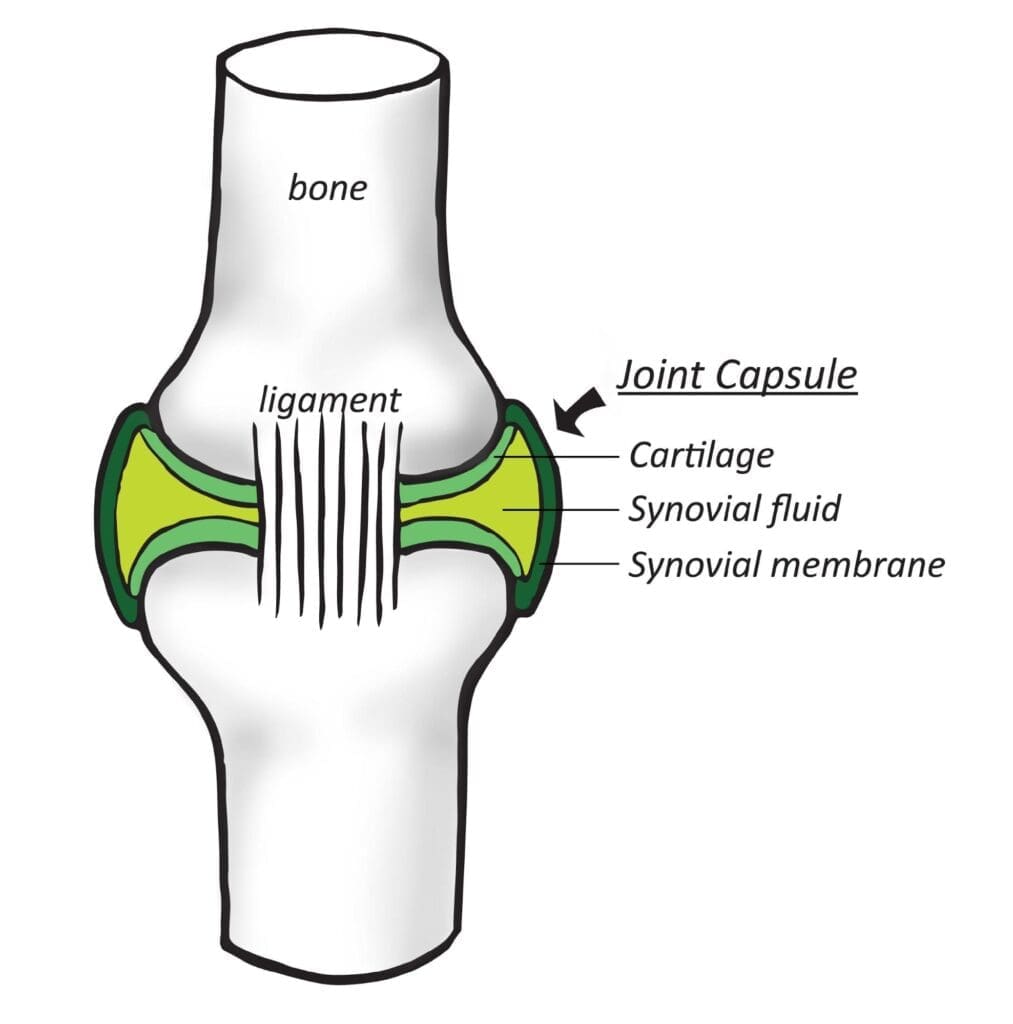
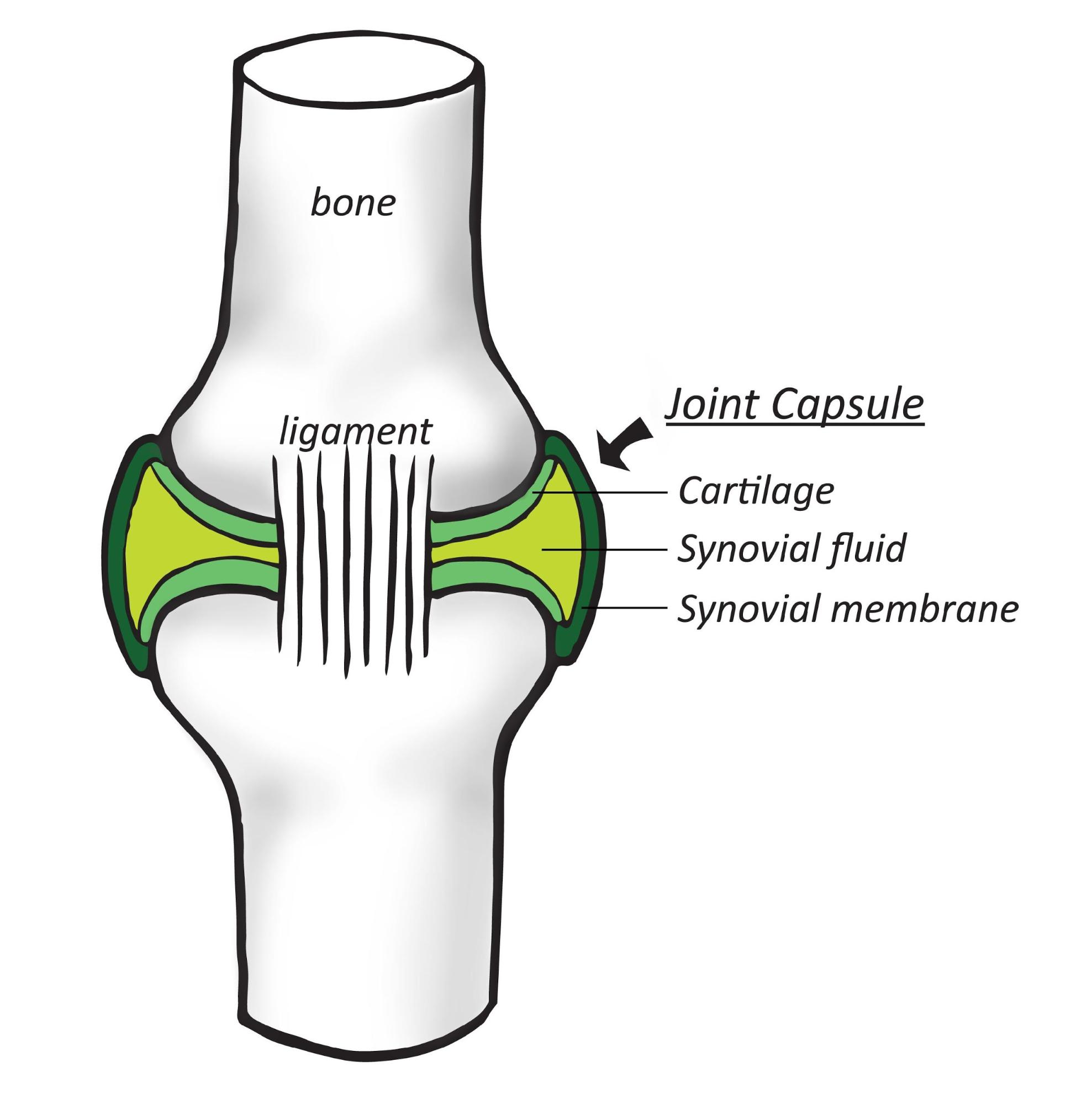
Whales have been viewed as a source of CO2 because they respire tons of CO2 annually. However, their feces could possibly offset this impact, as they may be a great contributor to carbon export (removal from the atmosphere) to the depths of the ocean. Iron-rich whale feces stimulate the growth of phytoplankton, which leads to more CO2 drawn into the ocean through photosynthesis.
Lavery et al. conducted this study to find out whether the 12,000 sperm whales in the Southern Ocean are acting as a carbon sink. The authors wondered whether the whales help the ocean absorb more carbon from the atmosphere than the whales themselves release through respiration. They note that these animals consume prey outside of but defecate within the photic zone (the layer nearest to the ocean surface), raising nutrient availability in the layer of ocean where photosynthesis is possible. Whale feces are also in liquid form, which disperses and persists within this area.
Using existing data on whale populations, consumption patterns, and average rates of iron retention compared to what isexpelled, the authors estimate that the South Ocean sperm whales contribute 36 tons of iron per year to the photic zone. After accounting for respiration rates, the authors conclude that whales do act as a net carbon sink by removing 2.4 X 105 metric tons of carbon from the atmosphere annually. Even under conservative scenarios (consumption of prey with lower iron concentrations), whales still help sequester more carbon than they respire.
These animals’ contribution to nutrient and carbon cycling in the ocean has previously been overlooked. Their feces not only enhance carbon sink in the ocean but also contribute to increasing numbers of prey. However, the reduction of sperm whales by commercial whaling has reduced krill populations and decreased allochthonous (originating externally) iron inputs to the Southern Ocean by 450 tons annually.
The reduction in sperm whale numbers owing to whaling has resulted in an extra 2 X 106 tonnes of carbon remaining in the atmosphere annually [Lavery 2010: 3].
In addition to sperm whales, there could be more organisms acting as carbon sinks in the ocean:
We have restricted our analysis to sperm whales; however, any organism that consumes prey outside the photic zone and defecates nutrient-rich waste that persists in the photic zone would stimulate new production and carbon export. Pygmy and dwarf sperm whales (Kogia spp.) and beaked whales (Family Ziphiidae) fulfill these criteria. The proportion of time baleen whales consume prey at depth is currently unknown, but fin whales (Balaenoptera physalus) dive to at least 470 m while feeding. Seals and sealions often consume prey at depth, but whether the[ir] waste is liquid (and buoyant) requires further investigation. [Lavery 2010: 4]
The ocean sequesters about 22% of global anthropogenic CO2emissions. Marine vertebrates contribute to the ocean’s carbon sink capacity in various ways, such as by fertilizing coastal vegetated habitats, and (through the work of marine predators) protecting this vegetation from overgrazing. Additionally, fish sequester carbon in the deep sea when they sink to the bottom after their natural death, whereas fishing releases the carbon embodied in fish back into the atmosphere when the catch is processed and consumed. Large fish (tuna, mackerel, shark, and billfish) that die in the ocean particularly contribute to “blue carbon” because these species are more likely to sink than be eaten near the surface. Unlike the CO2 released by terrestrial animals after death, the embodied carbon in marine corpses remains in the deep ocean.
This study estimates the extent to which fisheries have obstructed blue carbon sequestration. Mariani et al. report that fishing prevented 21.8 ± 4.4 Mt C (million metric tons of carbon) between 1950 and 2014 from being sequestered in the deep ocean. Industrial fisheries (as opposed to smaller, artisanal fisheries) are responsible for 85% of this extraction.
The amount of blue carbon extracted from the ocean through the harvest of large fish increased by almost one order of magnitude in 65 years (from 0.13 Mt C in 1950 to 1.09 Mt C in 2015). Combining CO2 emissions from fishing fleet transport and that of the fish removal itself amounts to 20.4 MtCO2 emitted in 2014, which is equivalent to the annual emission of 4.5 million cars.
Moreover, the authors found that government subsidies are encouraging overfishing. Almost half of the blue carbon extracted from the world’s oceans comes from areas that would be economically unprofitable without subsidies.
Our findings thus show that government subsidies, through supporting large-scale exploitation of large-bodied fish that are economically unviable, exacerbate the depletion of a natural carbon sink [Mariani 2010: 2].
Limiting and managing all fisheries on the unprofitable areas of the oceans could reduce CO2 emissions, rebuild fish stocks, and promote carbon sequestration by increasing the populations of large-bodied fish and the eventual deadfall of their carcasses to the depths.
Billions of animals, including insects, mammals, fish, and birds, migrate through the planet every year, which uniquely influences the environment and the ecological communities along migration routes.
“The frequency of migrations and the immense number of individuals involved often mean that migrant inputs constitute “resource pulses,” defined as occasional, intense, brief episodes of increased resource availability that can profoundly alter demographic rates and abundances of interacting populations” [Bauer & Hoye 2014: 6]
Effect on nutrients, energy, and toxicants:
Migrants transport nutrients, energy, and other substances from one ecosystem to another, creating a net inflow of energy and nutrients into the destination ecosystem. For example, salmon increased the nitrogen and phosphorus in their spawning habitat by 190% and 390% when migrating from the ocean back to their natal lakes and streams. At the same time, migrants may also introduce and accumulate toxicants, such as heavy metals, to receiving communities.
Effect on propagule dispersal:
Migrants play an important role in dispersing propagules, such as seeds, suckers, or spores across the resident communities.
In light of the importance of dispersal for population structure, adaptive capabilities, and evolutionary trajectories in theoretical studies, such long-distance dispersal events may be highly important for the (re)colonization of unoccupied habitats, the recovery of lost populations, maintenance of gene flow, and gene mixing in metapopulations, even if they are relatively rare events [Bauer & Hoye 2014: 2].
Moreover, migrants could also disperse propagules within resident communities. For example, long-nosed bats are responsible for up to 100% of columnar cacti pollination when they migrate to western Mexico. It is important to note that the timing of migration is very important; the migrants can only serve as major pollinators when visiting the communities during peak flowering.
Effect on parasite dispersal:
Migrants may increase parasite dynamics by facilitating the long-distance dispersal of parasites (including zoonotic pathogens like Ebola that also affect humans) to resident species. A few key mechanisms are involved in migration-facilitated parasite dispersal. For example, migrating animals are likely exposed to a greater range of parasites than are resident species. Some migrant animals may have suppressed immune responses due to the high investment of energy into migration, increasing their susceptibility to infection. In addition, while migrating, animals tend to aggregate in larger groups, thus enhancing transmission rates, compared to other times of the year when they are stationary.
However, the role of migrants in transmitting parasites is complicated. Studies of monarch butterflies have shown that they have a shorter flying distance when infected with parasites, andinfected Bewick’s swans delay their departure and travel shorter distances. These findings suggest that migrants may reduce infection risk through infection-induced delays.
Effect of migratory herbivores (plant-eating species):
Migrants may alter the nutrient cycling, productivity, the biomass of edible plants, and ground cover of dead plant material. The grazing intensity of migrant herbivores is decoupled from the timing of plant growth so plants can grow when they are left, which substantially increases the primary productivity compared to an ecosystem with the equivalent number of resident herbivores.
The outcome of the interaction between migrants and residents differs depending on the food resources. During periods of plenty of food residents could share the excess resources with the migrants. However, during the dry season when food is scarcer, synergistic negative effects may be created.
Effects of migratory predators:
Migratory predators can positively influence the communities through prey population control. For example, birds and bats may control the insect population, which reduces damage to crops. Seasonal outmigration may also reduce pressure on prey in the places left behind by migrants, allowing those populations to regrow.
Effects of migratory prey:
Migratory prey could be an important resource for resident predators. Some predators even time their reproduction to coincide with migratory prey to increase their reproductive rate.
Migratory prey may also provide resident prey with a temporal refuge from predation. However, an abundant number of migrants may harm residents by boosting the abundance of resident predators, which then switch toresident prey after the migratory prey departs.
Many ecosystems have evolved to depend upon the activities of both resident and transitory migrating animals, and understanding these relationships is critical to preserving and restoring ecosystem complexity and resiliency.
Across the globe, migration is an increasingly threatened phenomenon as a consequence of habitat destruction, creation of barriers, over-exploitation, and climate change. The loss of migrants and migratory behavior also entails the loss of their ecosystem services—the manifold transport and trophic effects outlined above. Management strategies must therefore be designed to conserve not only migratory species but also their ecosystem functions. Yet, the conservation of migrants poses exceptional scientific and societal challenges, as events at each stage of the migratory cycle affect behavior and demographic rates and ecological interactions at other stages [Bauer & Hoye 2014: 9].
This paper presents an analysis of microclimatic temperature effects of termite mounds in Zimbabwe and South Africa that provide important climatic “refuges” for other local organisms. The research compared the vegetation growing on the mounds with that on control plots in the surrounding savannah with respect to temperature differences. They found that more tall woody vegetation grows on termite mounds, compared to surrounding areas, creating shade that cools the mounds.
The authors observed that: “tall trees, being more prevalent on mounds, provide increased leafy, large-volume canopy and subcanopy vegetation, which in turn furnish more shade relative to the savanna matrix” [Joseph 2016: 7]. They found a 2°C temperature difference on the termite mounds compared to the surrounding area when the surrounding temperature was 34°C; the difference rose to 4°C at 40°C. Thus, these mound microhabitats maintained an even greater ambient temperature difference the warmer the ambient environment became.
Data were collected on 44 large termite mounds, each paired with off-mound savannah plots, in October 2015 (which was one of the hottest months on record in these areas) during the dry season. The mounds were more than 2 meters tall or more than 10 meters in diameter, and they were compared with an equivalently sized circular plot in the surrounding habitat. For each termite mound and control plot, the variables measured included: temperature, humidity, number of trees taller than 4 meters, tree canopy size, and amount of shade.
The median mean shade on mounds was 21% compared to 3% on the control plots, while the median maximum shade was 70% on mounds and only 10% on the surrounding plots, while humidity did not differ significantly. Such microclimates are likely to be important refugia for wildlife as droughts, fire events and higher ambient temperatures become more prevalent due to climate change.
Many large herbivores may have important roles in dryland ecosystems. Equids such as donkeys and horses, as well as elephants, have been reported to dig wells of a maximum depth of two meters, enhancing water availability for a variety of animals and plants. Noting that this subject has received limited research attention, the authors carried out a study for three summers at the Sonoran Desert of North America to survey changes in groundwater-fed streams and “equid well” water, and the associated effects on the ecosystem.
Effect on animals
They found that the equid wells “provided up to 74% of surface water by accessing the water table” at one of the four groundwater-fed streams they studied [Lundgren 2014: 1]. The wells were especially important at the intermittent stream (unsteady stream that occurs at irregular intervals), providing 100% of available surface water when all other water was lost.
The wells reduced the distance between neighboring water features significantly, thus reducing the distance that animals needed to travel to reach water. The water resources created by the equids also prevented some species from resorting to eating extra plant foods simply to extract its water content, as they are observed to do in the absence of available surface water. Using camera traps, the researchers observed 59 vertebrate species (limited to organisms weighing over 100g and excluding equids) at equid wells, 57 of which they recorded drinking. “Daily species richness was 64 and 51% higher on average at equid wells and background waters [other surface water, such as the streams], respectively, than at dry controls” [Lundgren 2014: 1].
Effects on vegetation
The presence of equid wells enhanced the growth of pioneer trees. The survey showed that the seeding density was higher in equid wells, which function as germination nurseries, than in the riverbank zone. Riverbanks were usually covered by herbs, which reduced the density of trees. Equid wells, on the other hand, provide a non-competitive environment for the small-seeded pioneer trees.
The feral donkeys that dug the equid wells are not native to this dryland ecosystem study site, and yet they proved to mitigate the effects of water reduction and high temperature on biodiversity and ecosystem function. Thus, the ecological roles once played by large native mammals that have since become extinct, can in some cases be filled by non-native substitutes (which are typically viewed as threats to conservation).
“The majority of the world’s 350,000 species of flowering plants rely on animal pollinators for reproduction” [Tremlett 2019: 2]. Of the many vertebrates performing this function, including birds, rodents, and reptiles, bats are thought to be the primary pollinators for about 1,000 species of plants across the tropics.
The authors of this study conducted this research in the municipality of Techaluta de Montenegro, Jalisco, Mexico, where they held exclusion experiments (alternately excluding different pollinator species) on Stenocereus queretaroensis, a type of cactus with edible fruit, to determine the efficiency of different pollinators. The experimental treatments allowed the authors to distinguish between nocturnal and diurnal (active in the daytime) pollinators, and between invertebrate and vertebrate pollinators.
Pollination carried out by birds and diurnal insects resulted in low seed sets, significantly lighter fruit weights, and lower sucrose concentrations compared to pollination carried out by bats.
This was the first research study to assess the impact of bat pollination on not only the quality of a high socio-economically important crop but also the yield of the crop.
We found that in the absence of pollination by nectarivorous bats, yield and quality (i.e. fruit weight, as size determines market value) of S. queretaroensis decreased significantly by 35% and 46% respectively. Hence, nectarivorous bats contribute substantially to the economic welfare of the rural production region [Tremlett 2019: 6].
However, despite its economic value, the significance of pollination by bats is not valued and appreciated. It is important to recognize the ecosystem services provided by bats, which might be crucial to sustaining rural livelihoods and well-being.
After having been wiped out by the 1920s, wolves were reintroduced to Yellowstone National Park in 1995-1996. This study assessed the importance of large carnivores to wild ungulates’ behavior and density, with secondary effects on plant communities, rivers and channels, and beaver communities. Focusing on the West and East Forks of Blacktail Deer Creek, the authors summarized the population trends of wolves, elk, and beaver; sampled the heights, recruitment, and browsing intensity of Geyer willow (a common local tall willow); measured dimensions of the channel, and ascertained beaver dam heights.
After the reintroduction of wolves, the Rocky Mountain elk population decreased from 17,000 in 1994 to about 4,000 to 5,000 in recent years. Browsing intensity therefore greatly decreased, leading to taller riparian willow stems, which is an important food web support and physical habitat for both terrestrial and aquatic wildlife species. The willow canopy cover over the water surface has also increased rapidly over the last two decades, which holds a significant role in supporting the aquatic biota:
Canopy cover can reduce the amount of solar radiation reaching a stream, especially important during summertime periods when solar angles are high, day lengths are long, and flows are normally low, thereby mediating potential increases in water temperature. Furthermore, invertebrates in the canopies of near‐channel willows provide food for fish and seasonal leaf‐fall represents an important carbon base for aquatic invertebrates which, in turn, provide ‘reciprocal flows of invertebrate prey’ to adjacent terrestrial consumers [Beschta & Ripple 2020: 8].
Another benefit of protecting the riparian vegetation from herbivores is the improvement of streambank stability. During the period of wolf absence, intensive elk herbivory caused streambank erosion and channel incision (river cuts downward into its bed, deepening the active channel and may lead to dissected landscape), resulting in less frequent overbank flow. The channel incision lowered water tables and reduced subsurface moisture in flood plain vegetation during summer.
The return of wolves started the process of riparian vegetation restoration, which in turn supported stream-dependent species such as beavers. The reduction of elk herbivory increased food sources and materials for beavers to construct dams, while also fostering the narrower and shallower channels preferred by beavers. Thus, along with the recovery of vegetation and channels, beavers have returned in 2018, creating active dams to further rehabilitate the ecosystem.
If beaver populations continue to increase over time, the ecological effects of these ‘ecosystem engineers’ may well have a significant role in restoring riparian vegetation, floodplains, and channel dimensions for at least portions of northern range streams [Beschta & Ripple 2020: 9].
Before the 1900s, wolves and other predators, such as bears and mountain lions, helped control the populations of herbivores in Yellowstone. However, the federal government exterminated these predators in a coordinated campaign. After the last wolf pack was killed, the elk numbers started increasing uncountably. The US Park Service subsequently attempted to control the elk population by shooting the animals or moving them out of the park.
When the park stopped killing elk in 1968, numbers shot up again from about 5,000 to close to 20,000. For the next several decades, elk cycled through population booms and collapses along with climate fluctuations; hard winters left the ground littered with hundreds of the carcasses of elk that had starved to death [Peterson 2020].
Wildlife officials, therefore, reintroduced wolves back to Yellowstone 25 years ago, which brought the elk population under control and ended their extreme population fluctuations due to climate variability. To study how the wolves maintained the balance, the scientists tracked the wolf packs and recorded details of elk kills by the wolves.
They found that the wolves killed cow elk during the years with normal amounts of rain and snow. During the dry years, when there is less vegetation and therefore less elk food, the wolves targeted bulls. The undernourished elk are generally easier to catch, so the wolves target bulls given their larger size. Sparing elk cows allows the elk to reproduce.
The wolves improve elk herd resilience by eliminating the weak and sick animals. Scientists believed the elk herds are now better prepared for climate change impact, such as the frequent droughts.
The result of reintroducing wolves to Yellowstone showed that wolves stabilize the elk population better than humans can. Now wolves may be reintroduced to other states which are home to a large number of elk.
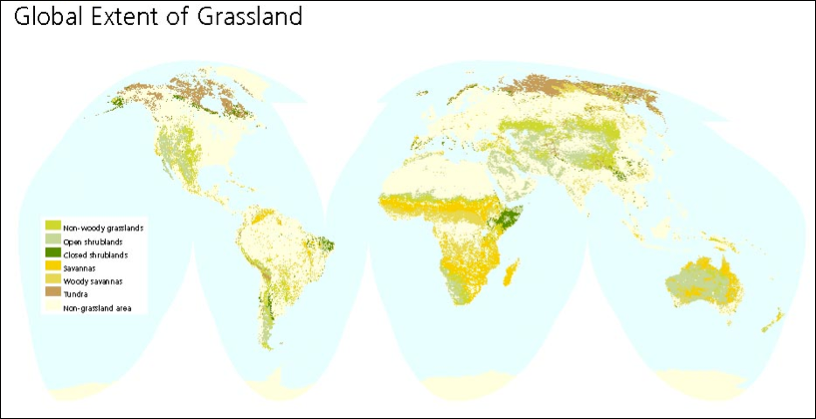
This article considers the overlooked role of grasslands and large herbivores in carbon storage. The principal question the authors pose is: what is the impact of large wild and domestic herbivores on the ability of ecosystems to absorb and store carbon over the long term? Their answer is that the activity of species like cattle, bison, boars, elephants, and rhinoceros, can significantly enhance ecosystem retention of carbon.
Questioning the assumption that fast-growing aboveground vegetation, especially trees, is the primary nature-based terrestrial sink for carbon, the authors argue for a whole-ecosystem carbon storage perspective. One problem with focusing on carbon storage in aboveground vegetation rather than that in the soil, they note, is that vegetation is more transient and vulnerable to disturbances, such as fire, while soil carbon tends to be stable, at least under natural and well-managed grasslands. Furthermore, the authors argue that the conventional focus on aboveground carbon storage has led to the “simplistic” generalization that large herbivores can be expected to damage vegetative ecosystems, and therefore have a negative impact on ecosystem carbon storage.
Yet this view misses the overall ecological impact of large herbivores, such as contributing to the soil through their wastes, and their bioturbation (churning of the soil by animals) activity. On the surface, large animals trample, forage, wallow and dig; just below the surface, the burrowing and digging of tunnels by soil-dwelling mammals like gophers, moles, voles, and shrews further loosens the soil; still deeper, there is the casting, burrowing and mining by macrofauna like insects, worms and dung beetles. Together, the multi-levelled bioturbation of these different species facilitates the vertical mixing of the organic material, putting it into contact with mineral soil particles for longer-term storage. Large grazing herbivores participate in vertical soil mixing (along with the smaller animals at lower levels in the soil), and therefore play an essential role in the long-term buildup of mineral-associated organic matter.
In addition to disturbing and mixing the soil, and enriching it through their body wastes, large herbivores clear pyrogenic (combustible) material on the ground and low shrubbery, thus increasing fire resistance. Their grazing also increases fine root growth and root exudation, which leads to increased microbial biomass. In turn, “microbial residues and plant exudates are effective substrates for persistent soil organic matter formation in the mineral-associated organic matter” [Kristensen 2021: 4].
In their conclusion, the authors emphasize the ecological value of natural grasslands, and the importance of preserving them:
Understanding the role that large herbivores may play in enhancing ecosystem carbon persistence, by reducing the flammability of aboveground carbon and shifting carbon storage from vulnerable pools towards more persistent soil pools at the biome scale, is crucial to balancing the ecosystem services provided by semi-open herbivore-rich systems against potential services from alternative land-uses, such as afforestation [Kristensen 2021: 9].
Can large herbivores enhance ecosystem carbon persistence? Kristensen et al. 2021
This article considers the overlooked role of grasslands and large herbivores in carbon storage. The principal question the authors pose is: what is the impact of large wild and domestic herbivores on the ability of ecosystems to absorb and store carbon over the long term? Their answer is that the activity of species like cattle, bison, boars, elephants, and rhinoceros, can significantly enhance ecosystem retention of carbon.
Questioning the assumption that fast-growing aboveground vegetation, especially trees, is the primary nature-based terrestrial sink for carbon, the authors argue for a whole-ecosystem carbon storage perspective. One problem with focusing on carbon storage in aboveground vegetation rather than that in the soil, they note, is that vegetation is more transient and vulnerable to disturbances, such as fire, while soil carbon tends to be stable, at least under natural and well-managed grasslands. Furthermore, the authors argue that the conventional focus on aboveground carbon storage has led to the “simplistic” generalization that large herbivores can be expected to damage vegetative ecosystems, and therefore have a negative impact on ecosystem carbon storage.
Yet this view misses the overall ecological impact of large herbivores, such as contributing to the soil through their wastes, and their bioturbation (churning of the soil by animals) activity. On the surface, large animals trample, forage, wallow and dig; just below the surface, the burrowing and digging of tunnels by soil-dwelling mammals like gophers, moles, voles, and shrews further loosens the soil; still deeper, there is the casting, burrowing and mining by macrofauna like insects, worms and dung beetles. Together, the multi-levelled bioturbation of these different species facilitates the vertical mixing of the organic material, putting it into contact with mineral soil particles for longer-term storage. Large grazing herbivores participate in vertical soil mixing (along with the smaller animals at lower levels in the soil), and therefore play an essential role in the long-term buildup of mineral-associated organic matter.
In addition to disturbing and mixing the soil, and enriching it through their body wastes, large herbivores clear pyrogenic (combustible) material on the ground and low shrubbery, thus increasing fire resistance. Their grazing also increases fine root growth and root exudation, which leads to increased microbial biomass. In turn, “microbial residues and plant exudates are effective substrates for persistent soil organic matter formation in the mineral-associated organic matter” [Kristensen 2021: 4].
In their conclusion, the authors emphasize the ecological value of natural grasslands, and the importance of preserving them:
Understanding the role that large herbivores may play in enhancing ecosystem carbon persistence, by reducing the flammability of aboveground carbon and shifting carbon storage from vulnerable pools towards more persistent soil pools at the biome scale, is crucial to balancing the ecosystem services provided by semi-open herbivore-rich systems against potential services from alternative land-uses, such as afforestation [Kristensen 2021: 9].
25 years after returning to Yellowstone, wolves have helped stabilize the ecosystem, Peterson 2020
Before the 1900s, wolves and other predators, such as bears and mountain lions, helped control the populations of herbivores in Yellowstone. However, the federal government exterminated these predators in a coordinated campaign. After the last wolf pack was killed, the elk numbers started increasing uncountably. The US Park Service subsequently attempted to control the elk population by shooting the animals or moving them out of the park.
When the park stopped killing elk in 1968, numbers shot up again from about 5,000 to close to 20,000. For the next several decades, elk cycled through population booms and collapses along with climate fluctuations; hard winters left the ground littered with hundreds of the carcasses of elk that had starved to death [Peterson 2020].
Wildlife officials, therefore, reintroduced wolves back to Yellowstone 25 years ago, which brought the elk population under control and ended their extreme population fluctuations due to climate variability. To study how the wolves maintained the balance, the scientists tracked the wolf packs and recorded details of elk kills by the wolves.
They found that the wolves killed cow elk during the years with normal amounts of rain and snow. During the dry years, when there is less vegetation and therefore less elk food, the wolves targeted bulls. The undernourished elk are generally easier to catch, so the wolves target bulls given their larger size. Sparing elk cows allows the elk to reproduce.
The wolves improve elk herd resilience by eliminating the weak and sick animals. Scientists believed the elk herds are now better prepared for climate change impact, such as the frequent droughts.
The result of reintroducing wolves to Yellowstone showed that wolves stabilize the elk population better than humans can. Now wolves may be reintroduced to other states which are home to a large number of elk.
Can large carnivores change streams via a trophic cascade? Beschta & Ripple 2020
After having been wiped out by the 1920s, wolves were reintroduced to Yellowstone National Park in 1995-1996. This study assessed the importance of large carnivores to wild ungulates’ behavior and density, with secondary effects on plant communities, rivers and channels, and beaver communities. Focusing on the West and East Forks of Blacktail Deer Creek, the authors summarized the population trends of wolves, elk, and beaver; sampled the heights, recruitment, and browsing intensity of Geyer willow (a common local tall willow); measured dimensions of the channel, and ascertained beaver dam heights.
After the reintroduction of wolves, the Rocky Mountain elk population decreased from 17,000 in 1994 to about 4,000 to 5,000 in recent years. Browsing intensity therefore greatly decreased, leading to taller riparian willow stems, which is an important food web support and physical habitat for both terrestrial and aquatic wildlife species. The willow canopy cover over the water surface has also increased rapidly over the last two decades, which holds a significant role in supporting the aquatic biota:
Canopy cover can reduce the amount of solar radiation reaching a stream, especially important during summertime periods when solar angles are high, day lengths are long, and flows are normally low, thereby mediating potential increases in water temperature. Furthermore, invertebrates in the canopies of near‐channel willows provide food for fish and seasonal leaf‐fall represents an important carbon base for aquatic invertebrates which, in turn, provide ‘reciprocal flows of invertebrate prey’ to adjacent terrestrial consumers [Beschta & Ripple 2020: 8].
Another benefit of protecting the riparian vegetation from herbivores is the improvement of streambank stability. During the period of wolf absence, intensive elk herbivory caused streambank erosion and channel incision (river cuts downward into its bed, deepening the active channel and may lead to dissected landscape), resulting in less frequent overbank flow. The channel incision lowered water tables and reduced subsurface moisture in flood plain vegetation during summer.
The return of wolves started the process of riparian vegetation restoration, which in turn supported stream-dependent species such as beavers. The reduction of elk herbivory increased food sources and materials for beavers to construct dams, while also fostering the narrower and shallower channels preferred by beavers. Thus, along with the recovery of vegetation and channels, beavers have returned in 2018, creating active dams to further rehabilitate the ecosystem.
If beaver populations continue to increase over time, the ecological effects of these ‘ecosystem engineers’ may well have a significant role in restoring riparian vegetation, floodplains, and channel dimensions for at least portions of northern range streams [Beschta & Ripple 2020: 9].
Pollination by bats enhances both quality and yield of a major cash crop in Mexico, Tremlett et al. 2019
“The majority of the world’s 350,000 species of flowering plants rely on animal pollinators for reproduction” [Tremlett 2019: 2]. Of the many vertebrates performing this function, including birds, rodents, and reptiles, bats are thought to be the primary pollinators for about 1,000 species of plants across the tropics.
The authors of this study conducted this research in the municipality of Techaluta de Montenegro, Jalisco, Mexico, where they held exclusion experiments (alternately excluding different pollinator species) on Stenocereus queretaroensis, a type of cactus with edible fruit, to determine the efficiency of different pollinators. The experimental treatments allowed the authors to distinguish between nocturnal and diurnal (active in the daytime) pollinators, and between invertebrate and vertebrate pollinators.
Pollination carried out by birds and diurnal insects resulted in low seed sets, significantly lighter fruit weights, and lower sucrose concentrations compared to pollination carried out by bats.
This was the first research study to assess the impact of bat pollination on not only the quality of a high socio-economically important crop but also the yield of the crop.
We found that in the absence of pollination by nectarivorous bats, yield and quality (i.e. fruit weight, as size determines market value) of S. queretaroensis decreased significantly by 35% and 46% respectively. Hence, nectarivorous bats contribute substantially to the economic welfare of the rural production region [Tremlett 2019: 6].
However, despite its economic value, the significance of pollination by bats is not valued and appreciated. It is important to recognize the ecosystem services provided by bats, which might be crucial to sustaining rural livelihoods and well-being.
Equids engineer desert water availability, Lundgren et al. 2014
Many large herbivores may have important roles in dryland ecosystems. Equids such as donkeys and horses, as well as elephants, have been reported to dig wells of a maximum depth of two meters, enhancing water availability for a variety of animals and plants. Noting that this subject has received limited research attention, the authors carried out a study for three summers at the Sonoran Desert of North America to survey changes in groundwater-fed streams and “equid well” water, and the associated effects on the ecosystem.
Effect on animals
They found that the equid wells “provided up to 74% of surface water by accessing the water table” at one of the four groundwater-fed streams they studied [Lundgren 2014: 1]. The wells were especially important at the intermittent stream (unsteady stream that occurs at irregular intervals), providing 100% of available surface water when all other water was lost.
The wells reduced the distance between neighboring water features significantly, thus reducing the distance that animals needed to travel to reach water. The water resources created by the equids also prevented some species from resorting to eating extra plant foods simply to extract its water content, as they are observed to do in the absence of available surface water. Using camera traps, the researchers observed 59 vertebrate species (limited to organisms weighing over 100g and excluding equids) at equid wells, 57 of which they recorded drinking. “Daily species richness was 64 and 51% higher on average at equid wells and background waters [other surface water, such as the streams], respectively, than at dry controls” [Lundgren 2014: 1].
Effects on vegetation
The presence of equid wells enhanced the growth of pioneer trees. The survey showed that the seeding density was higher in equid wells, which function as germination nurseries, than in the riverbank zone. Riverbanks were usually covered by herbs, which reduced the density of trees. Equid wells, on the other hand, provide a non-competitive environment for the small-seeded pioneer trees.
The feral donkeys that dug the equid wells are not native to this dryland ecosystem study site, and yet they proved to mitigate the effects of water reduction and high temperature on biodiversity and ecosystem function. Thus, the ecological roles once played by large native mammals that have since become extinct, can in some cases be filled by non-native substitutes (which are typically viewed as threats to conservation).
Microclimates mitigate against hot temperatures in dryland ecosystems: termite mounds as an example, Joseph et al. 2016
This paper presents an analysis of microclimatic temperature effects of termite mounds in Zimbabwe and South Africa that provide important climatic “refuges” for other local organisms. The research compared the vegetation growing on the mounds with that on control plots in the surrounding savannah with respect to temperature differences. They found that more tall woody vegetation grows on termite mounds, compared to surrounding areas, creating shade that cools the mounds.
The authors observed that: “tall trees, being more prevalent on mounds, provide increased leafy, large-volume canopy and subcanopy vegetation, which in turn furnish more shade relative to the savanna matrix” [Joseph 2016: 7]. They found a 2°C temperature difference on the termite mounds compared to the surrounding area when the surrounding temperature was 34°C; the difference rose to 4°C at 40°C. Thus, these mound microhabitats maintained an even greater ambient temperature difference the warmer the ambient environment became.
Data were collected on 44 large termite mounds, each paired with off-mound savannah plots, in October 2015 (which was one of the hottest months on record in these areas) during the dry season. The mounds were more than 2 meters tall or more than 10 meters in diameter, and they were compared with an equivalently sized circular plot in the surrounding habitat. For each termite mound and control plot, the variables measured included: temperature, humidity, number of trees taller than 4 meters, tree canopy size, and amount of shade.
The median mean shade on mounds was 21% compared to 3% on the control plots, while the median maximum shade was 70% on mounds and only 10% on the surrounding plots, while humidity did not differ significantly. Such microclimates are likely to be important refugia for wildlife as droughts, fire events and higher ambient temperatures become more prevalent due to climate change.
Billions of animals, including insects, mammals, fish, and birds, migrate through the planet every year, which uniquely influences the environment and the ecological communities along migration routes.
“The frequency of migrations and the immense number of individuals involved often mean that migrant inputs constitute “resource pulses,” defined as occasional, intense, brief episodes of increased resource availability that can profoundly alter demographic rates and abundances of interacting populations” [Bauer & Hoye 2014: 6]
Effect on nutrients, energy, and toxicants:
Migrants transport nutrients, energy, and other substances from one ecosystem to another, creating a net inflow of energy and nutrients into the destination ecosystem. For example, salmon increased the nitrogen and phosphorus in their spawning habitat by 190% and 390% when migrating from the ocean back to their natal lakes and streams. At the same time, migrants may also introduce and accumulate toxicants, such as heavy metals, to receiving communities.
Effect on propagule dispersal:
Migrants play an important role in dispersing propagules, such as seeds, suckers, or spores across the resident communities.
In light of the importance of dispersal for population structure, adaptive capabilities, and evolutionary trajectories in theoretical studies, such long-distance dispersal events may be highly important for the (re)colonization of unoccupied habitats, the recovery of lost populations, maintenance of gene flow, and gene mixing in metapopulations, even if they are relatively rare events [Bauer & Hoye 2014: 2].
Moreover, migrants could also disperse propagules within resident communities. For example, long-nosed bats are responsible for up to 100% of columnar cacti pollination when they migrate to western Mexico. It is important to note that the timing of migration is very important; the migrants can only serve as major pollinators when visiting the communities during peak flowering.
Effect on parasite dispersal:
Migrants may increase parasite dynamics by facilitating the long-distance dispersal of parasites (including zoonotic pathogens like Ebola that also affect humans) to resident species. A few key mechanisms are involved in migration-facilitated parasite dispersal. For example, migrating animals are likely exposed to a greater range of parasites than are resident species. Some migrant animals may have suppressed immune responses due to the high investment of energy into migration, increasing their susceptibility to infection. In addition, while migrating, animals tend to aggregate in larger groups, thus enhancing transmission rates, compared to other times of the year when they are stationary.
However, the role of migrants in transmitting parasites is complicated. Studies of monarch butterflies have shown that they have a shorter flying distance when infected with parasites, andinfected Bewick’s swans delay their departure and travel shorter distances. These findings suggest that migrants may reduce infection risk through infection-induced delays.
Effect of migratory herbivores (plant-eating species):
Migrants may alter the nutrient cycling, productivity, the biomass of edible plants, and ground cover of dead plant material. The grazing intensity of migrant herbivores is decoupled from the timing of plant growth so plants can grow when they are left, which substantially increases the primary productivity compared to an ecosystem with the equivalent number of resident herbivores.
The outcome of the interaction between migrants and residents differs depending on the food resources. During periods of plenty of food residents could share the excess resources with the migrants. However, during the dry season when food is scarcer, synergistic negative effects may be created.
Effects of migratory predators:
Migratory predators can positively influence the communities through prey population control. For example, birds and bats may control the insect population, which reduces damage to crops. Seasonal outmigration may also reduce pressure on prey in the places left behind by migrants, allowing those populations to regrow.
Effects of migratory prey:
Migratory prey could be an important resource for resident predators. Some predators even time their reproduction to coincide with migratory prey to increase their reproductive rate.
Migratory prey may also provide resident prey with a temporal refuge from predation. However, an abundant number of migrants may harm residents by boosting the abundance of resident predators, which then switch toresident prey after the migratory prey departs.
Many ecosystems have evolved to depend upon the activities of both resident and transitory migrating animals, and understanding these relationships is critical to preserving and restoring ecosystem complexity and resiliency.
Across the globe, migration is an increasingly threatened phenomenon as a consequence of habitat destruction, creation of barriers, over-exploitation, and climate change. The loss of migrants and migratory behavior also entails the loss of their ecosystem services—the manifold transport and trophic effects outlined above. Management strategies must therefore be designed to conserve not only migratory species but also their ecosystem functions. Yet, the conservation of migrants poses exceptional scientific and societal challenges, as events at each stage of the migratory cycle affect behavior and demographic rates and ecological interactions at other stages [Bauer & Hoye 2014: 9].
Let more big fish sink: Fisheries prevent blue carbon sequestration—half in unprofitable areas, Mariani et. al 2010
The ocean sequesters about 22% of global anthropogenic CO2emissions. Marine vertebrates contribute to the ocean’s carbon sink capacity in various ways, such as by fertilizing coastal vegetated habitats, and (through the work of marine predators) protecting this vegetation from overgrazing. Additionally, fish sequester carbon in the deep sea when they sink to the bottom after their natural death, whereas fishing releases the carbon embodied in fish back into the atmosphere when the catch is processed and consumed. Large fish (tuna, mackerel, shark, and billfish) that die in the ocean particularly contribute to “blue carbon” because these species are more likely to sink than be eaten near the surface. Unlike the CO2 released by terrestrial animals after death, the embodied carbon in marine corpses remains in the deep ocean.
This study estimates the extent to which fisheries have obstructed blue carbon sequestration. Mariani et al. report that fishing prevented 21.8 ± 4.4 Mt C (million metric tons of carbon) between 1950 and 2014 from being sequestered in the deep ocean. Industrial fisheries (as opposed to smaller, artisanal fisheries) are responsible for 85% of this extraction.
The amount of blue carbon extracted from the ocean through the harvest of large fish increased by almost one order of magnitude in 65 years (from 0.13 Mt C in 1950 to 1.09 Mt C in 2015). Combining CO2 emissions from fishing fleet transport and that of the fish removal itself amounts to 20.4 MtCO2 emitted in 2014, which is equivalent to the annual emission of 4.5 million cars.
Moreover, the authors found that government subsidies are encouraging overfishing. Almost half of the blue carbon extracted from the world’s oceans comes from areas that would be economically unprofitable without subsidies.
Our findings thus show that government subsidies, through supporting large-scale exploitation of large-bodied fish that are economically unviable, exacerbate the depletion of a natural carbon sink [Mariani 2010: 2].
Limiting and managing all fisheries on the unprofitable areas of the oceans could reduce CO2 emissions, rebuild fish stocks, and promote carbon sequestration by increasing the populations of large-bodied fish and the eventual deadfall of their carcasses to the depths.
Iron defecation by sperm whales stimulates carbon export in the Southern Ocean, Lavery et. al 2010
Whales have been viewed as a source of CO2 because they respire tons of CO2 annually. However, their feces could possibly offset this impact, as they may be a great contributor to carbon export (removal from the atmosphere) to the depths of the ocean. Iron-rich whale feces stimulate the growth of phytoplankton, which leads to more CO2 drawn into the ocean through photosynthesis.
Lavery et al. conducted this study to find out whether the 12,000 sperm whales in the Southern Ocean are acting as a carbon sink. The authors wondered whether the whales help the ocean absorb more carbon from the atmosphere than the whales themselves release through respiration. They note that these animals consume prey outside of but defecate within the photic zone (the layer nearest to the ocean surface), raising nutrient availability in the layer of ocean where photosynthesis is possible. Whale feces are also in liquid form, which disperses and persists within this area.
Using existing data on whale populations, consumption patterns, and average rates of iron retention compared to what isexpelled, the authors estimate that the South Ocean sperm whales contribute 36 tons of iron per year to the photic zone. After accounting for respiration rates, the authors conclude that whales do act as a net carbon sink by removing 2.4 X 105 metric tons of carbon from the atmosphere annually. Even under conservative scenarios (consumption of prey with lower iron concentrations), whales still help sequester more carbon than they respire.
These animals’ contribution to nutrient and carbon cycling in the ocean has previously been overlooked. Their feces not only enhance carbon sink in the ocean but also contribute to increasing numbers of prey. However, the reduction of sperm whales by commercial whaling has reduced krill populations and decreased allochthonous (originating externally) iron inputs to the Southern Ocean by 450 tons annually.
The reduction in sperm whale numbers owing to whaling has resulted in an extra 2 X 106 tonnes of carbon remaining in the atmosphere annually [Lavery 2010: 3].
In addition to sperm whales, there could be more organisms acting as carbon sinks in the ocean:
We have restricted our analysis to sperm whales; however, any organism that consumes prey outside the photic zone and defecates nutrient-rich waste that persists in the photic zone would stimulate new production and carbon export. Pygmy and dwarf sperm whales (Kogia spp.) and beaked whales (Family Ziphiidae) fulfill these criteria. The proportion of time baleen whales consume prey at depth is currently unknown, but fin whales (Balaenoptera physalus) dive to at least 470 m while feeding. Seals and sealions often consume prey at depth, but whether the[ir] waste is liquid (and buoyant) requires further investigation. [Lavery 2010: 4]
Animals contribute vitally to Earth’s water, carbon, and nutrient cycles. Every ecosystem is supported by uncountable animal species, ranging from birds to insects and mammals to fish, as well as microscopic organisms. The devastating news is that the Earth is losing about 150 animal, plant and microbial species every day, mostly due to human activities.[8] Understanding the ecological value of animals could bring attention to and support for actions and policy to protect animals and the ecosystems they compose.
On land, large herbivores can enhance carbon storage and maintain a healthy ecosystem. Grazing is commonly viewed as damaging to the grasslands–and that is the case when herds are overpopulated or otherwise not leaving sufficient time for pastures to recover after grazing. But when herbivore populations are in balance with the ecosystem, grazing is beneficial and stimulates the growth of grasses. Grazing animals nourish soil with their waste and churn the soil to facilitate theincorporation of organic material through daily activity. Grazing also removes pyrogenic (combustible) carbon from the land surface and increases grasslands’ fire resistance.
Grazing becomes unbalanced when top predators are lost. Animals and plants have coevolved, each species relying on numerous other species both for food and population control. Just as wolves need elk for food, elk need wolves to keep the herd healthy by eliminating weak and sick elk and keeping local carrying capacity from being breached.
Plants need wolves too, as do all species that depend on those plants when, in the absence of predators, herbivore populations grow too large. In one of the most successful rewilding experiments, the reintroduction of wolves in Yellowstone National Park in the mid-1990s effectively stabilized the elk population, eliminating the need for the government to remove elk from the Northern Yellowstone herd due to overgrazing.
Wolf-mediated control of the herbivore population benefited plant communities, rivers, and streams. Beschta and Ripple [2020] explain that prior to wolf reintroduction, elk grazing along stream banks kept willow trees from growing tall and shading the stream. Overgrazing also eroded the bank and deepened the streambed, ultimately reducing the frequency of the natural overbank flow that had previously nourished the floodplain.
By 2017 (compared to the 1990s), however, these authors write, willow heights had nearly quadrupled, while:
Canopy cover over the stream, essentially absent in 1995, had increased to 43% and 93% along the West Fork and East Fork, respectively. These recent increases in tall willow heights, greater canopy cover, well‐vegetated streambanks, and the recent development of an inset floodplain all pointed towards a riparian/aquatic ecosystem beginning to recover [Beschta & Ripple 2020: 1].
Australia historically hosted an array of megafauna. Now that some species have gone extinct, there is a void left on the continent. Introduced wild donkeys fill that void by grazing, digging wells, and browsing vegetation. These behaviors improve plant and soil health by contributing to the nutrient cycle. Wild donkeys venture into areas where cows do not, meaning that these non-native species have found their place in Australia’s increasingly dry landscape.
Australia is not the only place from which large mammal populations have disappeared. Only one-third of 730 terrestrial ecoregions today have intact mammal assemblages, meaning that all of the species that were present on the landscape 500 years ago remain today. Noting that large mammals are critical ecosystem engineers, a 2022 study estimates that the reintroduction of 20 priority species (including bear, bison, beaver, cougar, deer, and gazelle, for example) “can trigger restoration of complete assemblages over 54% of the terrestrial realm,” thus improving overall ecosystem function [Vynne 2022: 1].
Creatures come in all shapes and sizes, and even the smallest ones play an ecological role. By building mounds that support denser, taller vegetation than surrounding land, termites create microclimates in hot arid environments that are up to 4°C cooler than elsewhere on the landscape. Cool, shady termite mounds thus become vital refuges for other species.
Ladybugs contribute directly to agricultural systems by keeping pest populations under control. Ladybugs prey on aphids, mealybugs, and other creatures that have an appetite for crops. Rather than investing in artificial pesticides made up of harsh chemicals that damage beneficial plants and harm wildlife, farmers can work with ladybugs to maintain the health of their crops.
We cannot reestablish the Earth’s balance without addressing the body of water that covers over 70% of the planet. One type of marine invertebrate, the sea sponges, support clean oceans by filtering water. Sea sponges also provide a home for other animals living inside or on their surface, and take part in natural underwater construction by helping corals anchor to substrate. Once corals find a secure place to grow, they build colorful reefs, and sea sponges are one reason these underwater cities continue to flourish.
Sperm whales stimulate carbon sequestration in the Southern Ocean. Lavery et al. [2010] demonstrate that South Ocean sperm whales’ iron-rich feces are a critical phytoplankton fertilizer, boosting photosynthesis and drawing in carbon, which ultimately sinks to the deep ocean. In this way, sperm whales are removing 2.4 X 105 metric tons of carbon from the atmosphere annually. However, commercial whaling greatly limits this carbon export activity:
The reduction in sperm whale numbers owing to whaling has resulted in an extra 2 X 106 tonnes of carbon remaining in the atmosphere annually. [Lavery 2010: 3]
Overfishing also reduces the ocean’s carbon storage potential. Mariani et al. [2010] found that between 1950 and 2014, the removal of tuna, mackerel, shark, and billfish, mainly by industrial fisheries, prevented 21.8 ± 4.4 Mt C (million metric tons of carbon) from being sequestered. Had the fish not been caught, but rather died a natural death, their bodies would have sunk to the bottom of the ocean and remained there. Unlike animal corpses decomposing on land, which release CO2, the embodied carbon in marine animals remains in the depths of the ocean after death.
Migratory animals – whether in the ocean, on land, or in the sky – interact in ecosystems at a macroscopic level. Bauer & Hoye [2014] showed that migratory animals influence the herbivory, predatory, and reproductive patterns of other species, redistribute nutrients, and disperse propagules (such as seeds, suckers, or spores), toxicants and parasites along their migration routes. Migrating animals may even enhance (re)colonization of unoccupied or lost habitat through propagule dispersal.
Recognizing and appreciating the importance of other species allows us to reconnect with nature and natural cycles. The mutually beneficial relationships between lands, waterways, and animals render animals inseparable from and indispensable to these ecosystems. Removing just one species from an ecosystem can lead to drastic effects on the entire ecosystem. When species are able fulfill their respective roles in ecosystems, those ecosystems maintain their function and balance.
To learn more about the fascinating and varied ways that wildly diverse creatures help to make our world tick, explore Biodiversity for a Livable Climate’s Featured Creature series athttps://bio4climate.org/featured-creature/.
Summaries of articles on the ecological roles of animals
Can large herbivores enhance ecosystem carbon persistence? Kristensen et al. 2021
This article considers the overlooked role of grasslands and large herbivores in carbon storage. The principal question the authors pose is: what is the impact of large wild and domestic herbivores on the ability of ecosystems to absorb and store carbon over the long term? Their answer is that the activity of species like cattle, bison, boars, elephants, and rhinoceros, can significantly enhance ecosystem retention of carbon.
Questioning the assumption that fast-growing aboveground vegetation, especially trees, is the primary nature-based terrestrial sink for carbon, the authors argue for a whole-ecosystem carbon storage perspective. One problem with focusing on carbon storage in aboveground vegetation rather than that in the soil, they note, is that vegetation is more transient and vulnerable to disturbances, such as fire, while soil carbon tends to be stable, at least under natural and well-managed grasslands. Furthermore, the authors argue that the conventional focus on aboveground carbon storage has led to the “simplistic” generalization that large herbivores can be expected to damage vegetative ecosystems, and therefore have a negative impact on ecosystem carbon storage.
Yet this view misses the overall ecological impact of large herbivores, such as contributing to the soil through their wastes, and their bioturbation (churning of the soil by animals) activity. On the surface, large animals trample, forage, wallow and dig; just below the surface, the burrowing and digging of tunnels by soil-dwelling mammals like gophers, moles, voles, and shrews further loosens the soil; still deeper, there is the casting, burrowing and mining by macrofauna like insects, worms and dung beetles. Together, the multi-levelled bioturbation of these different species facilitates the vertical mixing of the organic material, putting it into contact with mineral soil particles for longer-term storage. Large grazing herbivores participate in vertical soil mixing (along with the smaller animals at lower levels in the soil), and therefore play an essential role in the long-term buildup of mineral-associated organic matter.
In addition to disturbing and mixing the soil, and enriching it through their body wastes, large herbivores clear pyrogenic (combustible) material on the ground and low shrubbery, thus increasing fire resistance. Their grazing also increases fine root growth and root exudation, which leads to increased microbial biomass. In turn, “microbial residues and plant exudates are effective substrates for persistent soil organic matter formation in the mineral-associated organic matter” [Kristensen 2021: 4].
In their conclusion, the authors emphasize the ecological value of natural grasslands, and the importance of preserving them:
Understanding the role that large herbivores may play in enhancing ecosystem carbon persistence, by reducing the flammability of aboveground carbon and shifting carbon storage from vulnerable pools towards more persistent soil pools at the biome scale, is crucial to balancing the ecosystem services provided by semi-open herbivore-rich systems against potential services from alternative land-uses, such as afforestation [Kristensen 2021: 9].
25 years after returning to Yellowstone, wolves have helped stabilize the ecosystem, Peterson 2020
Before the 1900s, wolves and other predators, such as bears and mountain lions, helped control the populations of herbivores in Yellowstone. However, the federal government exterminated these predators in a coordinated campaign. After the last wolf pack was killed, the elk numbers started increasing uncountably. The US Park Service subsequently attempted to control the elk population by shooting the animals or moving them out of the park.
When the park stopped killing elk in 1968, numbers shot up again from about 5,000 to close to 20,000. For the next several decades, elk cycled through population booms and collapses along with climate fluctuations; hard winters left the ground littered with hundreds of the carcasses of elk that had starved to death [Peterson 2020].
Wildlife officials, therefore, reintroduced wolves back to Yellowstone 25 years ago, which brought the elk population under control and ended their extreme population fluctuations due to climate variability. To study how the wolves maintained the balance, the scientists tracked the wolf packs and recorded details of elk kills by the wolves.
They found that the wolves killed cow elk during the years with normal amounts of rain and snow. During the dry years, when there is less vegetation and therefore less elk food, the wolves targeted bulls. The undernourished elk are generally easier to catch, so the wolves target bulls given their larger size. Sparing elk cows allows the elk to reproduce.
The wolves improve elk herd resilience by eliminating the weak and sick animals. Scientists believed the elk herds are now better prepared for climate change impact, such as the frequent droughts.
The result of reintroducing wolves to Yellowstone showed that wolves stabilize the elk population better than humans can. Now wolves may be reintroduced to other states which are home to a large number of elk.
Can large carnivores change streams via a trophic cascade? Beschta & Ripple 2020
After having been wiped out by the 1920s, wolves were reintroduced to Yellowstone National Park in 1995-1996. This study assessed the importance of large carnivores to wild ungulates’ behavior and density, with secondary effects on plant communities, rivers and channels, and beaver communities. Focusing on the West and East Forks of Blacktail Deer Creek, the authors summarized the population trends of wolves, elk, and beaver; sampled the heights, recruitment, and browsing intensity of Geyer willow (a common local tall willow); measured dimensions of the channel, and ascertained beaver dam heights.
After the reintroduction of wolves, the Rocky Mountain elk population decreased from 17,000 in 1994 to about 4,000 to 5,000 in recent years. Browsing intensity therefore greatly decreased, leading to taller riparian willow stems, which is an important food web support and physical habitat for both terrestrial and aquatic wildlife species. The willow canopy cover over the water surface has also increased rapidly over the last two decades, which holds a significant role in supporting the aquatic biota:
Canopy cover can reduce the amount of solar radiation reaching a stream, especially important during summertime periods when solar angles are high, day lengths are long, and flows are normally low, thereby mediating potential increases in water temperature. Furthermore, invertebrates in the canopies of near‐channel willows provide food for fish and seasonal leaf‐fall represents an important carbon base for aquatic invertebrates which, in turn, provide ‘reciprocal flows of invertebrate prey’ to adjacent terrestrial consumers [Beschta & Ripple 2020: 8].
Another benefit of protecting the riparian vegetation from herbivores is the improvement of streambank stability. During the period of wolf absence, intensive elk herbivory caused streambank erosion and channel incision (river cuts downward into its bed, deepening the active channel and may lead to dissected landscape), resulting in less frequent overbank flow. The channel incision lowered water tables and reduced subsurface moisture in flood plain vegetation during summer.
The return of wolves started the process of riparian vegetation restoration, which in turn supported stream-dependent species such as beavers. The reduction of elk herbivory increased food sources and materials for beavers to construct dams, while also fostering the narrower and shallower channels preferred by beavers. Thus, along with the recovery of vegetation and channels, beavers have returned in 2018, creating active dams to further rehabilitate the ecosystem.
If beaver populations continue to increase over time, the ecological effects of these ‘ecosystem engineers’ may well have a significant role in restoring riparian vegetation, floodplains, and channel dimensions for at least portions of northern range streams [Beschta & Ripple 2020: 9].
Pollination by bats enhances both quality and yield of a major cash crop in Mexico, Tremlett et al. 2019
“The majority of the world’s 350,000 species of flowering plants rely on animal pollinators for reproduction” [Tremlett 2019: 2]. Of the many vertebrates performing this function, including birds, rodents, and reptiles, bats are thought to be the primary pollinators for about 1,000 species of plants across the tropics.
The authors of this study conducted this research in the municipality of Techaluta de Montenegro, Jalisco, Mexico, where they held exclusion experiments (alternately excluding different pollinator species) on Stenocereus queretaroensis, a type of cactus with edible fruit, to determine the efficiency of different pollinators. The experimental treatments allowed the authors to distinguish between nocturnal and diurnal (active in the daytime) pollinators, and between invertebrate and vertebrate pollinators.
Pollination carried out by birds and diurnal insects resulted in low seed sets, significantly lighter fruit weights, and lower sucrose concentrations compared to pollination carried out by bats.
This was the first research study to assess the impact of bat pollination on not only the quality of a high socio-economically important crop but also the yield of the crop.
We found that in the absence of pollination by nectarivorous bats, yield and quality (i.e. fruit weight, as size determines market value) of S. queretaroensis decreased significantly by 35% and 46% respectively. Hence, nectarivorous bats contribute substantially to the economic welfare of the rural production region [Tremlett 2019: 6].
However, despite its economic value, the significance of pollination by bats is not valued and appreciated. It is important to recognize the ecosystem services provided by bats, which might be crucial to sustaining rural livelihoods and well-being.
Equids engineer desert water availability, Lundgren et al. 2014
Many large herbivores may have important roles in dryland ecosystems. Equids such as donkeys and horses, as well as elephants, have been reported to dig wells of a maximum depth of two meters, enhancing water availability for a variety of animals and plants. Noting that this subject has received limited research attention, the authors carried out a study for three summers at the Sonoran Desert of North America to survey changes in groundwater-fed streams and “equid well” water, and the associated effects on the ecosystem.
Effect on animals
They found that the equid wells “provided up to 74% of surface water by accessing the water table” at one of the four groundwater-fed streams they studied [Lundgren 2014: 1]. The wells were especially important at the intermittent stream (unsteady stream that occurs at irregular intervals), providing 100% of available surface water when all other water was lost.
The wells reduced the distance between neighboring water features significantly, thus reducing the distance that animals needed to travel to reach water. The water resources created by the equids also prevented some species from resorting to eating extra plant foods simply to extract its water content, as they are observed to do in the absence of available surface water. Using camera traps, the researchers observed 59 vertebrate species (limited to organisms weighing over 100g and excluding equids) at equid wells, 57 of which they recorded drinking. “Daily species richness was 64 and 51% higher on average at equid wells and background waters [other surface water, such as the streams], respectively, than at dry controls” [Lundgren 2014: 1].
Effects on vegetation
The presence of equid wells enhanced the growth of pioneer trees. The survey showed that the seeding density was higher in equid wells, which function as germination nurseries, than in the riverbank zone. Riverbanks were usually covered by herbs, which reduced the density of trees. Equid wells, on the other hand, provide a non-competitive environment for the small-seeded pioneer trees.
The feral donkeys that dug the equid wells are not native to this dryland ecosystem study site, and yet they proved to mitigate the effects of water reduction and high temperature on biodiversity and ecosystem function. Thus, the ecological roles once played by large native mammals that have since become extinct, can in some cases be filled by non-native substitutes (which are typically viewed as threats to conservation).
Microclimates mitigate against hot temperatures in dryland ecosystems: termite mounds as an example, Joseph et al. 2016
This paper presents an analysis of microclimatic temperature effects of termite mounds in Zimbabwe and South Africa that provide important climatic “refuges” for other local organisms. The research compared the vegetation growing on the mounds with that on control plots in the surrounding savannah with respect to temperature differences. They found that more tall woody vegetation grows on termite mounds, compared to surrounding areas, creating shade that cools the mounds.
The authors observed that: “tall trees, being more prevalent on mounds, provide increased leafy, large-volume canopy and subcanopy vegetation, which in turn furnish more shade relative to the savanna matrix” [Joseph 2016: 7]. They found a 2°C temperature difference on the termite mounds compared to the surrounding area when the surrounding temperature was 34°C; the difference rose to 4°C at 40°C. Thus, these mound microhabitats maintained an even greater ambient temperature difference the warmer the ambient environment became.
Data were collected on 44 large termite mounds, each paired with off-mound savannah plots, in October 2015 (which was one of the hottest months on record in these areas) during the dry season. The mounds were more than 2 meters tall or more than 10 meters in diameter, and they were compared with an equivalently sized circular plot in the surrounding habitat. For each termite mound and control plot, the variables measured included: temperature, humidity, number of trees taller than 4 meters, tree canopy size, and amount of shade.
The median mean shade on mounds was 21% compared to 3% on the control plots, while the median maximum shade was 70% on mounds and only 10% on the surrounding plots, while humidity did not differ significantly. Such microclimates are likely to be important refugia for wildlife as droughts, fire events and higher ambient temperatures become more prevalent due to climate change.
Billions of animals, including insects, mammals, fish, and birds, migrate through the planet every year, which uniquely influences the environment and the ecological communities along migration routes.
“The frequency of migrations and the immense number of individuals involved often mean that migrant inputs constitute “resource pulses,” defined as occasional, intense, brief episodes of increased resource availability that can profoundly alter demographic rates and abundances of interacting populations” [Bauer & Hoye 2014: 6]
Effect on nutrients, energy, and toxicants:
Migrants transport nutrients, energy, and other substances from one ecosystem to another, creating a net inflow of energy and nutrients into the destination ecosystem. For example, salmon increased the nitrogen and phosphorus in their spawning habitat by 190% and 390% when migrating from the ocean back to their natal lakes and streams. At the same time, migrants may also introduce and accumulate toxicants, such as heavy metals, to receiving communities.
Effect on propagule dispersal:
Migrants play an important role in dispersing propagules, such as seeds, suckers, or spores across the resident communities.
In light of the importance of dispersal for population structure, adaptive capabilities, and evolutionary trajectories in theoretical studies, such long-distance dispersal events may be highly important for the (re)colonization of unoccupied habitats, the recovery of lost populations, maintenance of gene flow, and gene mixing in metapopulations, even if they are relatively rare events [Bauer & Hoye 2014: 2].
Moreover, migrants could also disperse propagules within resident communities. For example, long-nosed bats are responsible for up to 100% of columnar cacti pollination when they migrate to western Mexico. It is important to note that the timing of migration is very important; the migrants can only serve as major pollinators when visiting the communities during peak flowering.
Effect on parasite dispersal:
Migrants may increase parasite dynamics by facilitating the long-distance dispersal of parasites (including zoonotic pathogens like Ebola that also affect humans) to resident species. A few key mechanisms are involved in migration-facilitated parasite dispersal. For example, migrating animals are likely exposed to a greater range of parasites than are resident species. Some migrant animals may have suppressed immune responses due to the high investment of energy into migration, increasing their susceptibility to infection. In addition, while migrating, animals tend to aggregate in larger groups, thus enhancing transmission rates, compared to other times of the year when they are stationary.
However, the role of migrants in transmitting parasites is complicated. Studies of monarch butterflies have shown that they have a shorter flying distance when infected with parasites, andinfected Bewick’s swans delay their departure and travel shorter distances. These findings suggest that migrants may reduce infection risk through infection-induced delays.
Effect of migratory herbivores (plant-eating species):
Migrants may alter the nutrient cycling, productivity, the biomass of edible plants, and ground cover of dead plant material. The grazing intensity of migrant herbivores is decoupled from the timing of plant growth so plants can grow when they are left, which substantially increases the primary productivity compared to an ecosystem with the equivalent number of resident herbivores.
The outcome of the interaction between migrants and residents differs depending on the food resources. During periods of plenty of food residents could share the excess resources with the migrants. However, during the dry season when food is scarcer, synergistic negative effects may be created.
Effects of migratory predators:
Migratory predators can positively influence the communities through prey population control. For example, birds and bats may control the insect population, which reduces damage to crops. Seasonal outmigration may also reduce pressure on prey in the places left behind by migrants, allowing those populations to regrow.
Effects of migratory prey:
Migratory prey could be an important resource for resident predators. Some predators even time their reproduction to coincide with migratory prey to increase their reproductive rate.
Migratory prey may also provide resident prey with a temporal refuge from predation. However, an abundant number of migrants may harm residents by boosting the abundance of resident predators, which then switch toresident prey after the migratory prey departs.
Many ecosystems have evolved to depend upon the activities of both resident and transitory migrating animals, and understanding these relationships is critical to preserving and restoring ecosystem complexity and resiliency.
Across the globe, migration is an increasingly threatened phenomenon as a consequence of habitat destruction, creation of barriers, over-exploitation, and climate change. The loss of migrants and migratory behavior also entails the loss of their ecosystem services—the manifold transport and trophic effects outlined above. Management strategies must therefore be designed to conserve not only migratory species but also their ecosystem functions. Yet, the conservation of migrants poses exceptional scientific and societal challenges, as events at each stage of the migratory cycle affect behavior and demographic rates and ecological interactions at other stages [Bauer & Hoye 2014: 9].
Let more big fish sink: Fisheries prevent blue carbon sequestration—half in unprofitable areas, Mariani et. al 2010
The ocean sequesters about 22% of global anthropogenic CO2emissions. Marine vertebrates contribute to the ocean’s carbon sink capacity in various ways, such as by fertilizing coastal vegetated habitats, and (through the work of marine predators) protecting this vegetation from overgrazing. Additionally, fish sequester carbon in the deep sea when they sink to the bottom after their natural death, whereas fishing releases the carbon embodied in fish back into the atmosphere when the catch is processed and consumed. Large fish (tuna, mackerel, shark, and billfish) that die in the ocean particularly contribute to “blue carbon” because these species are more likely to sink than be eaten near the surface. Unlike the CO2 released by terrestrial animals after death, the embodied carbon in marine corpses remains in the deep ocean.
This study estimates the extent to which fisheries have obstructed blue carbon sequestration. Mariani et al. report that fishing prevented 21.8 ± 4.4 Mt C (million metric tons of carbon) between 1950 and 2014 from being sequestered in the deep ocean. Industrial fisheries (as opposed to smaller, artisanal fisheries) are responsible for 85% of this extraction.
The amount of blue carbon extracted from the ocean through the harvest of large fish increased by almost one order of magnitude in 65 years (from 0.13 Mt C in 1950 to 1.09 Mt C in 2015). Combining CO2 emissions from fishing fleet transport and that of the fish removal itself amounts to 20.4 MtCO2 emitted in 2014, which is equivalent to the annual emission of 4.5 million cars.
Moreover, the authors found that government subsidies are encouraging overfishing. Almost half of the blue carbon extracted from the world’s oceans comes from areas that would be economically unprofitable without subsidies.
Our findings thus show that government subsidies, through supporting large-scale exploitation of large-bodied fish that are economically unviable, exacerbate the depletion of a natural carbon sink [Mariani 2010: 2].
Limiting and managing all fisheries on the unprofitable areas of the oceans could reduce CO2 emissions, rebuild fish stocks, and promote carbon sequestration by increasing the populations of large-bodied fish and the eventual deadfall of their carcasses to the depths.
Iron defecation by sperm whales stimulates carbon export in the Southern Ocean, Lavery et. al 2010
Whales have been viewed as a source of CO2 because they respire tons of CO2 annually. However, their feces could possibly offset this impact, as they may be a great contributor to carbon export (removal from the atmosphere) to the depths of the ocean. Iron-rich whale feces stimulate the growth of phytoplankton, which leads to more CO2 drawn into the ocean through photosynthesis.
Lavery et al. conducted this study to find out whether the 12,000 sperm whales in the Southern Ocean are acting as a carbon sink. The authors wondered whether the whales help the ocean absorb more carbon from the atmosphere than the whales themselves release through respiration. They note that these animals consume prey outside of but defecate within the photic zone (the layer nearest to the ocean surface), raising nutrient availability in the layer of ocean where photosynthesis is possible. Whale feces are also in liquid form, which disperses and persists within this area.
Using existing data on whale populations, consumption patterns, and average rates of iron retention compared to what isexpelled, the authors estimate that the South Ocean sperm whales contribute 36 tons of iron per year to the photic zone. After accounting for respiration rates, the authors conclude that whales do act as a net carbon sink by removing 2.4 X 105 metric tons of carbon from the atmosphere annually. Even under conservative scenarios (consumption of prey with lower iron concentrations), whales still help sequester more carbon than they respire.
These animals’ contribution to nutrient and carbon cycling in the ocean has previously been overlooked. Their feces not only enhance carbon sink in the ocean but also contribute to increasing numbers of prey. However, the reduction of sperm whales by commercial whaling has reduced krill populations and decreased allochthonous (originating externally) iron inputs to the Southern Ocean by 450 tons annually.
The reduction in sperm whale numbers owing to whaling has resulted in an extra 2 X 106 tonnes of carbon remaining in the atmosphere annually [Lavery 2010: 3].
In addition to sperm whales, there could be more organisms acting as carbon sinks in the ocean:
We have restricted our analysis to sperm whales; however, any organism that consumes prey outside the photic zone and defecates nutrient-rich waste that persists in the photic zone would stimulate new production and carbon export. Pygmy and dwarf sperm whales (Kogia spp.) and beaked whales (Family Ziphiidae) fulfill these criteria. The proportion of time baleen whales consume prey at depth is currently unknown, but fin whales (Balaenoptera physalus) dive to at least 470 m while feeding. Seals and sealions often consume prey at depth, but whether the[ir] waste is liquid (and buoyant) requires further investigation. [Lavery 2010: 4]
This article (also highlighted in Compendium v2n1) reviews research on the benefits of tree cover in relation to water and energy cycles.
Forests help produce rain. Vegetation releases water vapor through transpiration, increasing atmospheric moisture that is then transported by wind. In fact, “over most of the tropics, air that passes over forests for ten days typically produces at least twice as much rain as air that passes over sparse vegetation” [Ellison 2017: 53]. Forests also release biological particles, such as spores, bacteria and pollen into the atmosphere. Water condenses around these particles, forming raindrops.
In addition to the atmospheric moisture produced by forests that is transported by prevailing winds to generate downwind rain, forests also help transport moisture from the coasts to the interior of continents. According to the biotic pump theory [Makarieva & Gorshkov 2007], evapotranspiration over coastal forests creates low pressure zones that draw in atmospheric moisture from the ocean. This oceanic moisture eventually comes down as rain over land. Deforestation of coastal forests thus reduces this influx of moisture to land, while deforestation anywhere can decrease the reliability of rainfall downwind.
Through shading and evapotranspiration, forests cool the Earth’s surface in tropical and temperate climates. Due to a lower albedo compared to other land cover types at high latitudes, boreal forests potentially contribute to local warming. However, forests also increase cloud cover and thus albedo, higher in the atmosphere. In the absence of vegetation, such as in cities, solar energy remains in the environment in the form of heat, rather than driving evapotranspiration.
Using the sun’s energy, individual trees can transpire hundreds of liters of water per day. This represents a cooling power equivalent to 70 kWh for every 100 L of water transpired (enough to power two average household central air-conditioning units per day) [Ellison 2017: 54].
High-elevation forests have a unique potential to intercept fog and cloud droplets, which boosts tree growth, evapotranspiration, groundwater infiltration, and ultimately contributes up to 75% of catchment runoff.Tree cover can improve water infiltration due to increased organic matter to hold water and the presence of tree roots, which loosen and shade the soil and channel water into the ground. In areas where infiltration rates are greater than transpiration rates, the presence of trees increases groundwater recharge.
All of the aforementioned mechanisms distribute water naturally, hence reducing floods.
Local temperatures are affected not only by global climatic factors, but also by radiative (albedo) and non-radiative (evapotranspiration and convection) mechanisms related to local vegetation cover. Through evapotranspiration, solar energy is converted to latent heat and released from the planet’s surface, while convection refers to the turbulent mixing of air that dissipates sensible heat. The authors state that while albedo (reflectivity of land surface, which is often lower on forested land) is increasingly accounted for alongside greenhouse gases in climate models, the non-radiative mechanisms are not. However, the evapotranspiration and convection facilitated by vegetation have an important cooling effect and should therefore be included in models to avoid the risk of “promoting land sector policies that may be counter to the aims of mitigation or adaptation” [Bright 2017: 296].
The authors demonstrate that “non-radiative mechanisms dominate the local response in most regions for eight of nine common LCMC perturbations” [Bright 2017: 296]. Land cover and land management changes (LCMC) considered in the study include converting cropland or grassland to deciduous or evergreen forests. The authors found that gains in forest cover increased annual cooling in all but the northernmost latitudes, where the lower albedo of forests compared to grasslands had a warming effect that was stronger than the cooling effect of non-radiative mechanisms. In many regions, including much of Europe, the US, and the tropics, non-radiative cooling dominated albedo effects. “Over annual timescales, forest cover gains result in net cooling for many of the densely populated regions of the planet” [Bright 2017: 298].
Bright et al. conclude that “benchmarking the locally driven LCMC effect to that driven by global forcers (such as CO2) can provide an additional perspective by which to support the valuation of vegetated ecosystems and the local climate regulation services that they provide” [Bright 2017: 301].
This paper analyzes how land use and land cover change (LULCC) affects temperature and humidity. The authors examined the differential effects of forest versus deforested land on temperature and humidity by comparing different land-cover models. One model simulated the total potential vegetation (“PotVeg”) that would cover Earth in the absence of human interference, while the other was based on historical data of land use changes that occurred over a recent five-decade period (“AllHist”).
They found that deforested lands in mid-latitudes (North America, Eurasia) in the AllHist model are warmer and drier compared to the same lands covered in forest in the PotVeg model. Specifically, “conversion of mid-latitude natural forests to cropland and pastures is accompanied by an increase in the occurrence of hot-dry summers from once-in-a-decade to every 2–3 years” [Findell 2017: 1]. “Based on these simulations, the conversion of forests to cropland is coincident with much of the upper central US and central Europe experiencing extreme hot, dry summers” [Findell 2017: 6].
Turning lemons into lemonade, Sabajo et al. have used the great expansion of oil palm plantations and other crops in Indonesia to examine how such land-use change affects land surface temperature (LST). The authors observed a warming trend in the Jambi province of Sumatra of 1.05℃ and 1.56℃ in the morning and afternoon, respectively, between 2000 and 2015. The average morning (10:30 am) temperature increased by 0.07℃ per year; the midday afternoon (1:30 pm) temperature increased by 0.13℃ per year.
During roughly the same period (2000-2010), forest area decreased in Jambi by 17%, while oil palm and rubber plantations greatly expanded. Given that LST within the province’s remaining forests increased only .04C per year at 10:30 am, which the authors attribute mainly to global warming, they concluded that the overall higher province-wide daytime temperature increase was caused by the observed land cover change.
The team also compared temperatures between different land uses: forest, oil palm and rubber plantations, urban areas, and bare land. Despite having a higher albedo (reflectivity) than the forest areas, all converted (non-forest) lands were nonetheless warmer than forests, “suggesting that the albedo was not the dominant variable explaining the LST” [Sabajo 2017: 4629]. Evapotranspiration (ET) played a greater role. Non-vegetated surfaces (urban and bare) were the warmest.
The authors conclude: “The warming effect after forest conversion results from the reduced evaporative cooling, which was identified as the main determinant of regulating the surface temperature” [Sabajo 2017: 4631].
Deforestation has contributed to warming in the northern mid-latitudes of North America and Eurasia not only through a large contribution to global CO2 emissions, but also through biogeophysical effects. The latter refers to land-surface effects such as albedo and evapotranspiration, which vary according to the type of land cover. This study uses models to demonstrate that deforestation in the northern mid-latitudes has increased the intensity of hot days by about a third since pre-industrial times. Factoring in deforestation’s contribution to greenhouse gas emissions further increases deforestation’s heating effect.
Our best estimate suggests that the present-day contribution of deforestation to the TXx [yearly maximum temperature, or “hot days”] increase over this region still equals at least 50% once the warming entailed by the LCC [land cover change]-induced carbon emissions is considered [LeJeune 2018: 4].
“Extensive deforestation took place early in the industrial period over the northern mid-latitudes,” and then slowed down in the 20th Century [LeJeune 2018: 4]. By 1920, modeled increases of temperature “through biogeophysical effects had already reached 0.3°C (~75% of their present-day values) over the most deforested areas of North America and Eurasia. On average before 1920, local deforestation was responsible for most of the TXx [yearly maximum temperature] warming over these regions” [LeJeune 2018: 4]. Warming caused by greenhouse gases became more important during the 20th Century, “leading to a total warming of 1.3°C over North America and 1°C over Eurasia by the present-day” [LeJeune 2018: 4].
The amount of Earth’s green cover (measured as Leaf Area Index[6]) has increased globally since 1980, especially in northern latitudes, where growing seasons have lengthened. This is due mainly to increasing CO2 concentration, but also to warmer temperatures and changing precipitation patterns, nitrogen deposition, and land-use change (such as afforestation in China). Higher ambient CO2 can stimulate photosynthesis and reduce water loss, but the extent of the CO2 fertilization depends on the availability of other key nutrients (nitrogen and phosphorus) and water. Warmer temperatures due to climate change have increased greening in northern latitudes by extending the growing season, but diminished greening in the tropics, where temperatures were already optimal.
Greater global green cover has observable feedbacks on climate and the carbon cycle. In addition to offsetting 28% of anthropogenic emissions since 1980, vegetation affects hydrological cycles and air-surface temperatures. Since the 1980s, increased global evapotranspiration (ET) is mainly attributable to increased global greening. Higher transpiration rates from vegetation can reduce or enhance groundwater storage locally, depending on how the atmospheric moisture generated through evapotranspiration is recycled into rain and where that rain falls. In the world’s great rainforests, vegetation preserves groundwater.
The enhanced precipitation over transpiring regions is particularly evident in moist forests like the Amazon or Congo, which are ‘closed’ atmospheric systems where 80% of the rainfall originates from upwind ET. Such an efficient atmospheric water recycling mitigates water loss from the soil, sustains inland vegetation and maintains mesic[7] and humid ecosystems [Piao 2019: 9].
Vegetation affects land-surface temperature by way of ET (cooling effect) and albedo (warming or cooling effect, depending on how dark or light the surface is). While the relative strength of ET versus albedo varies by latitude, the net global effect of increasing vegetation cover is one of cooling the land surface.
While reforestation has been widely heralded as a means of sequestering carbon into the soil, there is growing evidence that it also serves to directly cool the land surface. But forests’ impacts on air temperature (measured over forests rather than within them) have been difficult to assess because of the confounding impacts of forest canopies on wind and temperature profiles near the surface. This study was implemented to create a new method for assessing to what degree forests also cool the air.
Most studies measure surface temperature, which “represents the aggregated temperature of solid canopy and soil elements,” and is measured at a midway point between the ground and the top of the canopy. Air temperature, on the other hand, is measured above the vegetation canopy (whether grasslands or forest). The study site, located in the Piedmont region near Durham North Carolina, consists of an old-field grassland, a pine forest, and an unevenly aged oak hickory forest, all within close proximity to each other. The study assessed temperatures at various heights in and above the grasslands and the two forests.
The authors found that surface temperatures are much lower in forests than in grassland; this difference often exceeds 5°C at midday during the growing season. Furthermore, the air is cooler over forests than over grasslands, though to a lesser degree than the surface temperature difference. The annual average air temperature difference of forests compared to grassland is 0.5°C to 1°C, while the difference reaches 2°C to 3°C during daytime growing season periods.
“Making the connection between land cover, surface temperature, and air temperature is becoming necessary for obtaining a complete picture of the climate mitigation and adaptation potential of managed land cover changes, including reforestation,” the authors conclude. “This energy balance perspective on the climate mitigation and adaptation potential of reforestation is especially relevant right now” given a recent global surge of interest in reforestation to sequester carbon [Novik & Katul 2020: 13].
Reforestation and afforestation (R&A) are well-established climate mitigation strategies in the wet tropics due to high carbon sequestration rates of forests/trees. However, at high latitudes (boreal regions), the low albedo of trees–compared to snow and other lighter land surfaces–leads to the absorption of energy, thus creating a warming effect that has a greater impact on temperature than the carbon capture accomplished by the limited vegetation productivity in boreal regions. This study explores the balance between albedo and carbon sequestration of forests at mid-latitudes, which has been less clear.
The authors found that forested areas have greater cloud cover than other types of land cover at midlatitudes, resulting in a higher albedo at the top of the atmosphere–where the clouds are–and leading to greater cooling. Specifically, they found “an association of forested lands with increased cloudiness… As a result, forests reflect extra solar radiation and thus reduce the radiative impacts of the lower surface albedo. This in turn implies a cooling effect of R&A at midlatitudes” [Cerasoli 2021: 1-2]. The increase in cloudiness is due to earlier afternoon cloud formation over forests compared to other vegetation types in wet regions.
Our results provide substantial evidence of remarkable benefits of R&A [reforestation and afforestation] around the 30° to 45° latitudinal range, due to the combined benefits of biomass gain and promotion of cloud formation over forests [Cerasoli 2021: 4].
Cloud cooling effects of afforestation and reforestation at midlatitudes, Cerasoli, Jin & Porporato 2021
Reforestation and afforestation (R&A) are well-established climate mitigation strategies in the wet tropics due to high carbon sequestration rates of forests/trees. However, at high latitudes (boreal regions), the low albedo of trees–compared to snow and other lighter land surfaces–leads to the absorption of energy, thus creating a warming effect that has a greater impact on temperature than the carbon capture accomplished by the limited vegetation productivity in boreal regions. This study explores the balance between albedo and carbon sequestration of forests at mid-latitudes, which has been less clear.
The authors found that forested areas have greater cloud cover than other types of land cover at midlatitudes, resulting in a higher albedo at the top of the atmosphere–where the clouds are–and leading to greater cooling. Specifically, they found “an association of forested lands with increased cloudiness… As a result, forests reflect extra solar radiation and thus reduce the radiative impacts of the lower surface albedo. This in turn implies a cooling effect of R&A at midlatitudes” [Cerasoli 2021: 1-2]. The increase in cloudiness is due to earlier afternoon cloud formation over forests compared to other vegetation types in wet regions.
Our results provide substantial evidence of remarkable benefits of R&A [reforestation and afforestation] around the 30° to 45° latitudinal range, due to the combined benefits of biomass gain and promotion of cloud formation over forests [Cerasoli 2021: 4].
The duality of reforestation impacts on surface and air temperature, Novick & Katul 2020
While reforestation has been widely heralded as a means of sequestering carbon into the soil, there is growing evidence that it also serves to directly cool the land surface. But forests’ impacts on air temperature (measured over forests rather than within them) have been difficult to assess because of the confounding impacts of forest canopies on wind and temperature profiles near the surface. This study was implemented to create a new method for assessing to what degree forests also cool the air.
Most studies measure surface temperature, which “represents the aggregated temperature of solid canopy and soil elements,” and is measured at a midway point between the ground and the top of the canopy. Air temperature, on the other hand, is measured above the vegetation canopy (whether grasslands or forest). The study site, located in the Piedmont region near Durham North Carolina, consists of an old-field grassland, a pine forest, and an unevenly aged oak hickory forest, all within close proximity to each other. The study assessed temperatures at various heights in and above the grasslands and the two forests.
The authors found that surface temperatures are much lower in forests than in grassland; this difference often exceeds 5°C at midday during the growing season. Furthermore, the air is cooler over forests than over grasslands, though to a lesser degree than the surface temperature difference. The annual average air temperature difference of forests compared to grassland is 0.5°C to 1°C, while the difference reaches 2°C to 3°C during daytime growing season periods.
“Making the connection between land cover, surface temperature, and air temperature is becoming necessary for obtaining a complete picture of the climate mitigation and adaptation potential of managed land cover changes, including reforestation,” the authors conclude. “This energy balance perspective on the climate mitigation and adaptation potential of reforestation is especially relevant right now” given a recent global surge of interest in reforestation to sequester carbon [Novik & Katul 2020: 13].
Characteristics, drivers and feedbacks of global greening, Piao et al. 2019
The amount of Earth’s green cover (measured as Leaf Area Index[6]) has increased globally since 1980, especially in northern latitudes, where growing seasons have lengthened. This is due mainly to increasing CO2 concentration, but also to warmer temperatures and changing precipitation patterns, nitrogen deposition, and land-use change (such as afforestation in China). Higher ambient CO2 can stimulate photosynthesis and reduce water loss, but the extent of the CO2 fertilization depends on the availability of other key nutrients (nitrogen and phosphorus) and water. Warmer temperatures due to climate change have increased greening in northern latitudes by extending the growing season, but diminished greening in the tropics, where temperatures were already optimal.
Greater global green cover has observable feedbacks on climate and the carbon cycle. In addition to offsetting 28% of anthropogenic emissions since 1980, vegetation affects hydrological cycles and air-surface temperatures. Since the 1980s, increased global evapotranspiration (ET) is mainly attributable to increased global greening. Higher transpiration rates from vegetation can reduce or enhance groundwater storage locally, depending on how the atmospheric moisture generated through evapotranspiration is recycled into rain and where that rain falls. In the world’s great rainforests, vegetation preserves groundwater.
The enhanced precipitation over transpiring regions is particularly evident in moist forests like the Amazon or Congo, which are ‘closed’ atmospheric systems where 80% of the rainfall originates from upwind ET. Such an efficient atmospheric water recycling mitigates water loss from the soil, sustains inland vegetation and maintains mesic[7] and humid ecosystems [Piao 2019: 9].
Vegetation affects land-surface temperature by way of ET (cooling effect) and albedo (warming or cooling effect, depending on how dark or light the surface is). While the relative strength of ET versus albedo varies by latitude, the net global effect of increasing vegetation cover is one of cooling the land surface.
Historical deforestation locally increased the intensity of hot days in northern mid-latitudes, Lejeune 2018
Deforestation has contributed to warming in the northern mid-latitudes of North America and Eurasia not only through a large contribution to global CO2 emissions, but also through biogeophysical effects. The latter refers to land-surface effects such as albedo and evapotranspiration, which vary according to the type of land cover. This study uses models to demonstrate that deforestation in the northern mid-latitudes has increased the intensity of hot days by about a third since pre-industrial times. Factoring in deforestation’s contribution to greenhouse gas emissions further increases deforestation’s heating effect.
Our best estimate suggests that the present-day contribution of deforestation to the TXx [yearly maximum temperature, or “hot days”] increase over this region still equals at least 50% once the warming entailed by the LCC [land cover change]-induced carbon emissions is considered [LeJeune 2018: 4].
“Extensive deforestation took place early in the industrial period over the northern mid-latitudes,” and then slowed down in the 20th Century [LeJeune 2018: 4]. By 1920, modeled increases of temperature “through biogeophysical effects had already reached 0.3°C (~75% of their present-day values) over the most deforested areas of North America and Eurasia. On average before 1920, local deforestation was responsible for most of the TXx [yearly maximum temperature] warming over these regions” [LeJeune 2018: 4]. Warming caused by greenhouse gases became more important during the 20th Century, “leading to a total warming of 1.3°C over North America and 1°C over Eurasia by the present-day” [LeJeune 2018: 4].
Expansion of oil palm and other cash crops causes an increase of the land surface temperature in the Jambi province in Indonesia, Sabajo 2017
Turning lemons into lemonade, Sabajo et al. have used the great expansion of oil palm plantations and other crops in Indonesia to examine how such land-use change affects land surface temperature (LST). The authors observed a warming trend in the Jambi province of Sumatra of 1.05℃ and 1.56℃ in the morning and afternoon, respectively, between 2000 and 2015. The average morning (10:30 am) temperature increased by 0.07℃ per year; the midday afternoon (1:30 pm) temperature increased by 0.13℃ per year.
During roughly the same period (2000-2010), forest area decreased in Jambi by 17%, while oil palm and rubber plantations greatly expanded. Given that LST within the province’s remaining forests increased only .04C per year at 10:30 am, which the authors attribute mainly to global warming, they concluded that the overall higher province-wide daytime temperature increase was caused by the observed land cover change.
The team also compared temperatures between different land uses: forest, oil palm and rubber plantations, urban areas, and bare land. Despite having a higher albedo (reflectivity) than the forest areas, all converted (non-forest) lands were nonetheless warmer than forests, “suggesting that the albedo was not the dominant variable explaining the LST” [Sabajo 2017: 4629]. Evapotranspiration (ET) played a greater role. Non-vegetated surfaces (urban and bare) were the warmest.
The authors conclude: “The warming effect after forest conversion results from the reduced evaporative cooling, which was identified as the main determinant of regulating the surface temperature” [Sabajo 2017: 4631].
The impact of anthropogenic land use and land cover change on regional climate extremes, Findell et al. 2017
This paper analyzes how land use and land cover change (LULCC) affects temperature and humidity. The authors examined the differential effects of forest versus deforested land on temperature and humidity by comparing different land-cover models. One model simulated the total potential vegetation (“PotVeg”) that would cover Earth in the absence of human interference, while the other was based on historical data of land use changes that occurred over a recent five-decade period (“AllHist”).
They found that deforested lands in mid-latitudes (North America, Eurasia) in the AllHist model are warmer and drier compared to the same lands covered in forest in the PotVeg model. Specifically, “conversion of mid-latitude natural forests to cropland and pastures is accompanied by an increase in the occurrence of hot-dry summers from once-in-a-decade to every 2–3 years” [Findell 2017: 1]. “Based on these simulations, the conversion of forests to cropland is coincident with much of the upper central US and central Europe experiencing extreme hot, dry summers” [Findell 2017: 6].
Local temperature response to land cover and management change driven by non-radiative processes, Bright et al. 2017
Local temperatures are affected not only by global climatic factors, but also by radiative (albedo) and non-radiative (evapotranspiration and convection) mechanisms related to local vegetation cover. Through evapotranspiration, solar energy is converted to latent heat and released from the planet’s surface, while convection refers to the turbulent mixing of air that dissipates sensible heat. The authors state that while albedo (reflectivity of land surface, which is often lower on forested land) is increasingly accounted for alongside greenhouse gases in climate models, the non-radiative mechanisms are not. However, the evapotranspiration and convection facilitated by vegetation have an important cooling effect and should therefore be included in models to avoid the risk of “promoting land sector policies that may be counter to the aims of mitigation or adaptation” [Bright 2017: 296].
The authors demonstrate that “non-radiative mechanisms dominate the local response in most regions for eight of nine common LCMC perturbations” [Bright 2017: 296]. Land cover and land management changes (LCMC) considered in the study include converting cropland or grassland to deciduous or evergreen forests. The authors found that gains in forest cover increased annual cooling in all but the northernmost latitudes, where the lower albedo of forests compared to grasslands had a warming effect that was stronger than the cooling effect of non-radiative mechanisms. In many regions, including much of Europe, the US, and the tropics, non-radiative cooling dominated albedo effects. “Over annual timescales, forest cover gains result in net cooling for many of the densely populated regions of the planet” [Bright 2017: 298].
Bright et al. conclude that “benchmarking the locally driven LCMC effect to that driven by global forcers (such as CO2) can provide an additional perspective by which to support the valuation of vegetated ecosystems and the local climate regulation services that they provide” [Bright 2017: 301].
Trees, forests and water: Cool insights for a hot world, Ellison et al. 2017
This article (also highlighted in Compendium v2n1) reviews research on the benefits of tree cover in relation to water and energy cycles.
Forests help produce rain. Vegetation releases water vapor through transpiration, increasing atmospheric moisture that is then transported by wind. In fact, “over most of the tropics, air that passes over forests for ten days typically produces at least twice as much rain as air that passes over sparse vegetation” [Ellison 2017: 53]. Forests also release biological particles, such as spores, bacteria and pollen into the atmosphere. Water condenses around these particles, forming raindrops.
In addition to the atmospheric moisture produced by forests that is transported by prevailing winds to generate downwind rain, forests also help transport moisture from the coasts to the interior of continents. According to the biotic pump theory [Makarieva & Gorshkov 2007], evapotranspiration over coastal forests creates low pressure zones that draw in atmospheric moisture from the ocean. This oceanic moisture eventually comes down as rain over land. Deforestation of coastal forests thus reduces this influx of moisture to land, while deforestation anywhere can decrease the reliability of rainfall downwind.
Through shading and evapotranspiration, forests cool the Earth’s surface in tropical and temperate climates. Due to a lower albedo compared to other land cover types at high latitudes, boreal forests potentially contribute to local warming. However, forests also increase cloud cover and thus albedo, higher in the atmosphere. In the absence of vegetation, such as in cities, solar energy remains in the environment in the form of heat, rather than driving evapotranspiration.
Using the sun’s energy, individual trees can transpire hundreds of liters of water per day. This represents a cooling power equivalent to 70 kWh for every 100 L of water transpired (enough to power two average household central air-conditioning units per day) [Ellison 2017: 54].
High-elevation forests have a unique potential to intercept fog and cloud droplets, which boosts tree growth, evapotranspiration, groundwater infiltration, and ultimately contributes up to 75% of catchment runoff.Tree cover can improve water infiltration due to increased organic matter to hold water and the presence of tree roots, which loosen and shade the soil and channel water into the ground. In areas where infiltration rates are greater than transpiration rates, the presence of trees increases groundwater recharge.
All of the aforementioned mechanisms distribute water naturally, hence reducing floods.
Earth is heating up: “Global surface temperature was 1.09°C higher in 2011– 2020 than 1850–1900,” according to the Intergovernmental Panel on Climate Change (IPCC)’s 6th Assessment Report.[3] Yet the mercury is not rising uniformly around the world – the Arctic is warming faster than are the lower latitudes, and temperatures over land are higher than over the ocean. Local temperatures everywhere are also affected by the type of land cover, with paved areas being hotter than vegetated land.
In Phoenix, Arizona, the temperature topped 110°F during the summer of 2020 – not just once, but for 53 days.[4] In the same year, Los Angeles experienced its hottest day ever: 122°F. Paris’ hottest day record was broken in 2019, and Buenos Aires’ second and third hottest days ever were in January 2022 (its hottest was in 1957). While climate change is to blame for the world’s increasing frequency of heatwaves, cities additionally experience the “urban heat island” (UHI) effect, which can add as much as 20°F compared to their rural surroundings.[5]
To deal with heatwaves, the city of Phoenix established a new team to manage the public health effects of heat and find ways to reduce it. Part of the plan is doubling the city tree canopy to 25% coverage. Urban heat islands are created by a low ratio of vegetated surfaces relative to concrete, asphalt, and other unvegetated and impervious surfaces, which absorb sunlight as heat and radiate it back out. By contrast, plants use the sun’s energy to transpire water into the atmosphere, a process that cools rather than heats the surrounding air.
Many cities are planting trees to dampen scorching summer heat as public awareness of the relationship between vegetation and the UHI grows. Expanding public green space and tree canopy is a practical way for localities to adapt to the juggernaut of a changing climate because increased vegetation cover not only lowers local temperatures, but can also reduce the severity of flooding and drought by improving rainwater infiltration.
Just as increasing tree cover reduces temperatures in cities, reforestation has a cooling effect at regional and even global scales. The studies profiled below explore a variety of aspects of the relationship between vegetation types, especially forests, and temperature. All produced in the past ten years, these studies show that reforestation (establishing forest on previously forested land) and afforestation (foresting land that was not previously in forest) can lower temperatures, and that deforestation, conversely, has raised temperatures across vast areas.
For example, historical conversion of natural forests in the midlatitudes to cropland and pasture has more than tripled the occurrence of hot-dry summers in those areas [Findell 2017]. Clearing forests heats the land not only by releasing CO2, which contributes to the greenhouse effect, but also changes the albedo (reflectivity), surface roughness, and evapotranspiration (ET) rates of land surfaces. Land-cover changes affect the temperature through biophysical exchanges of water and energy between the land and the atmosphere.
By 1920, the biophysical effects of deforestation were the main cause of increasing temperatures in deforested parts of North America and Eurasia [LeJeune 2018]. Over the 20th Century, greenhouse gases started to play a bigger role in regional heating. Today, the combined effects of the biophysical changes and greenhouse gas emissions wrought by deforestation account for an estimated half of the temperature rise since pre-industrial times in deforested areas of the midlatitudes [LeJeune 2018].
Interestingly, the presence of forests affects surface temperatures differently depending on latitude, although globally, vegetation gain has a net cooling effect [Piao 2020]. In boreal regions, forestation is associated with a warming effect due to a lower albedo of forests compared to non-forest land. By contrast, deforestation warms the mid-latitudes and tropics – in spite of the lower surface albedo of forested land. The cooling effect of ET drives temperature regulation in the tropics, while both ET and cloud formation from forests cool the mid-latitudes.
The Jambi province of tropical Sumatra, Indonesia, warmed an average of 1.05°C between 2000 and 2015, a period that coincided with rapid deforestation [Sabajo 2017]. The forests that remained in Jambi during this period also warmed slightly, but much less so than the region as a whole. The authors suggest that the smaller temperature rise inside forests reflects global warming, while the greater regional warming is due to the combined effect of global warming and the biophysical effects of local deforestation.
Vegetation cover both affects and is affected by climate change. Piao et al. [2019] documented an increase in global greening since 1980, which they attribute in part to growth-inducing impacts of climate change. Warmer temperatures and the fertilization effects of higher CO2 concentration, but also nitrogen deposition and direct afforestation efforts, all have increased vegetation cover, though not evenly across the planet, and not necessarily where it would have the greatest cooling effect. For example, warmer temperatures increase vegetation in the northern latitudes by lengthening the growing season, but reduce vegetation in the tropics where historical temperatures are already optimal for plant growth.
In addition to modulating temperatures, forests also influence precipitation. Through ET – the very same mechanism responsible for cooling the land – forests recycle water back into the atmosphere. Globally, at least 40% of the rain that falls on land comes from evapotranspiration on land, while as much as 70% of rain originating over the Amazon is ET-fed [Ellison 2017]. In Europe, converting a limited proportion (not expected to impinge on food security) of agricultural lands would increase rainfall on the continent by an estimated 7.6%, an important augmentation given the growing frequency of drought there [Baker 2021].
The role that forests and other natural vegetation types play in mitigating global warming and softening the blow of climate extremes has not always been universally clear, but this is changing. The studies highlighted below, which illustrate the heat- and drought-limiting capacity of forests, are among a growing body of work pointing to the importance–and power–of natural climate solutions.
Credit: Peter Donavan (soilcarboncoalition.org)
Summaries of articles showing the cooling effect of vegetation
Cloud cooling effects of afforestation and reforestation at midlatitudes, Cerasoli, Jin & Porporato 2021
Reforestation and afforestation (R&A) are well-established climate mitigation strategies in the wet tropics due to high carbon sequestration rates of forests/trees. However, at high latitudes (boreal regions), the low albedo of trees–compared to snow and other lighter land surfaces–leads to the absorption of energy, thus creating a warming effect that has a greater impact on temperature than the carbon capture accomplished by the limited vegetation productivity in boreal regions. This study explores the balance between albedo and carbon sequestration of forests at mid-latitudes, which has been less clear.
The authors found that forested areas have greater cloud cover than other types of land cover at midlatitudes, resulting in a higher albedo at the top of the atmosphere–where the clouds are–and leading to greater cooling. Specifically, they found “an association of forested lands with increased cloudiness… As a result, forests reflect extra solar radiation and thus reduce the radiative impacts of the lower surface albedo. This in turn implies a cooling effect of R&A at midlatitudes” [Cerasoli 2021: 1-2]. The increase in cloudiness is due to earlier afternoon cloud formation over forests compared to other vegetation types in wet regions.
Our results provide substantial evidence of remarkable benefits of R&A [reforestation and afforestation] around the 30° to 45° latitudinal range, due to the combined benefits of biomass gain and promotion of cloud formation over forests [Cerasoli 2021: 4].
The duality of reforestation impacts on surface and air temperature, Novick & Katul 2020
While reforestation has been widely heralded as a means of sequestering carbon into the soil, there is growing evidence that it also serves to directly cool the land surface. But forests’ impacts on air temperature (measured over forests rather than within them) have been difficult to assess because of the confounding impacts of forest canopies on wind and temperature profiles near the surface. This study was implemented to create a new method for assessing to what degree forests also cool the air.
Most studies measure surface temperature, which “represents the aggregated temperature of solid canopy and soil elements,” and is measured at a midway point between the ground and the top of the canopy. Air temperature, on the other hand, is measured above the vegetation canopy (whether grasslands or forest). The study site, located in the Piedmont region near Durham North Carolina, consists of an old-field grassland, a pine forest, and an unevenly aged oak hickory forest, all within close proximity to each other. The study assessed temperatures at various heights in and above the grasslands and the two forests.
The authors found that surface temperatures are much lower in forests than in grassland; this difference often exceeds 5°C at midday during the growing season. Furthermore, the air is cooler over forests than over grasslands, though to a lesser degree than the surface temperature difference. The annual average air temperature difference of forests compared to grassland is 0.5°C to 1°C, while the difference reaches 2°C to 3°C during daytime growing season periods.
“Making the connection between land cover, surface temperature, and air temperature is becoming necessary for obtaining a complete picture of the climate mitigation and adaptation potential of managed land cover changes, including reforestation,” the authors conclude. “This energy balance perspective on the climate mitigation and adaptation potential of reforestation is especially relevant right now” given a recent global surge of interest in reforestation to sequester carbon [Novik & Katul 2020: 13].
Characteristics, drivers and feedbacks of global greening, Piao et al. 2019
The amount of Earth’s green cover (measured as Leaf Area Index[6]) has increased globally since 1980, especially in northern latitudes, where growing seasons have lengthened. This is due mainly to increasing CO2 concentration, but also to warmer temperatures and changing precipitation patterns, nitrogen deposition, and land-use change (such as afforestation in China). Higher ambient CO2 can stimulate photosynthesis and reduce water loss, but the extent of the CO2 fertilization depends on the availability of other key nutrients (nitrogen and phosphorus) and water. Warmer temperatures due to climate change have increased greening in northern latitudes by extending the growing season, but diminished greening in the tropics, where temperatures were already optimal.
Greater global green cover has observable feedbacks on climate and the carbon cycle. In addition to offsetting 28% of anthropogenic emissions since 1980, vegetation affects hydrological cycles and air-surface temperatures. Since the 1980s, increased global evapotranspiration (ET) is mainly attributable to increased global greening. Higher transpiration rates from vegetation can reduce or enhance groundwater storage locally, depending on how the atmospheric moisture generated through evapotranspiration is recycled into rain and where that rain falls. In the world’s great rainforests, vegetation preserves groundwater.
The enhanced precipitation over transpiring regions is particularly evident in moist forests like the Amazon or Congo, which are ‘closed’ atmospheric systems where 80% of the rainfall originates from upwind ET. Such an efficient atmospheric water recycling mitigates water loss from the soil, sustains inland vegetation and maintains mesic[7] and humid ecosystems [Piao 2019: 9].
Vegetation affects land-surface temperature by way of ET (cooling effect) and albedo (warming or cooling effect, depending on how dark or light the surface is). While the relative strength of ET versus albedo varies by latitude, the net global effect of increasing vegetation cover is one of cooling the land surface.
Historical deforestation locally increased the intensity of hot days in northern mid-latitudes, Lejeune 2018
Deforestation has contributed to warming in the northern mid-latitudes of North America and Eurasia not only through a large contribution to global CO2 emissions, but also through biogeophysical effects. The latter refers to land-surface effects such as albedo and evapotranspiration, which vary according to the type of land cover. This study uses models to demonstrate that deforestation in the northern mid-latitudes has increased the intensity of hot days by about a third since pre-industrial times. Factoring in deforestation’s contribution to greenhouse gas emissions further increases deforestation’s heating effect.
Our best estimate suggests that the present-day contribution of deforestation to the TXx [yearly maximum temperature, or “hot days”] increase over this region still equals at least 50% once the warming entailed by the LCC [land cover change]-induced carbon emissions is considered [LeJeune 2018: 4].
“Extensive deforestation took place early in the industrial period over the northern mid-latitudes,” and then slowed down in the 20th Century [LeJeune 2018: 4]. By 1920, modeled increases of temperature “through biogeophysical effects had already reached 0.3°C (~75% of their present-day values) over the most deforested areas of North America and Eurasia. On average before 1920, local deforestation was responsible for most of the TXx [yearly maximum temperature] warming over these regions” [LeJeune 2018: 4]. Warming caused by greenhouse gases became more important during the 20th Century, “leading to a total warming of 1.3°C over North America and 1°C over Eurasia by the present-day” [LeJeune 2018: 4].
Expansion of oil palm and other cash crops causes an increase of the land surface temperature in the Jambi province in Indonesia, Sabajo 2017
Turning lemons into lemonade, Sabajo et al. have used the great expansion of oil palm plantations and other crops in Indonesia to examine how such land-use change affects land surface temperature (LST). The authors observed a warming trend in the Jambi province of Sumatra of 1.05℃ and 1.56℃ in the morning and afternoon, respectively, between 2000 and 2015. The average morning (10:30 am) temperature increased by 0.07℃ per year; the midday afternoon (1:30 pm) temperature increased by 0.13℃ per year.
During roughly the same period (2000-2010), forest area decreased in Jambi by 17%, while oil palm and rubber plantations greatly expanded. Given that LST within the province’s remaining forests increased only .04C per year at 10:30 am, which the authors attribute mainly to global warming, they concluded that the overall higher province-wide daytime temperature increase was caused by the observed land cover change.
The team also compared temperatures between different land uses: forest, oil palm and rubber plantations, urban areas, and bare land. Despite having a higher albedo (reflectivity) than the forest areas, all converted (non-forest) lands were nonetheless warmer than forests, “suggesting that the albedo was not the dominant variable explaining the LST” [Sabajo 2017: 4629]. Evapotranspiration (ET) played a greater role. Non-vegetated surfaces (urban and bare) were the warmest.
The authors conclude: “The warming effect after forest conversion results from the reduced evaporative cooling, which was identified as the main determinant of regulating the surface temperature” [Sabajo 2017: 4631].
The impact of anthropogenic land use and land cover change on regional climate extremes, Findell et al. 2017
This paper analyzes how land use and land cover change (LULCC) affects temperature and humidity. The authors examined the differential effects of forest versus deforested land on temperature and humidity by comparing different land-cover models. One model simulated the total potential vegetation (“PotVeg”) that would cover Earth in the absence of human interference, while the other was based on historical data of land use changes that occurred over a recent five-decade period (“AllHist”).
They found that deforested lands in mid-latitudes (North America, Eurasia) in the AllHist model are warmer and drier compared to the same lands covered in forest in the PotVeg model. Specifically, “conversion of mid-latitude natural forests to cropland and pastures is accompanied by an increase in the occurrence of hot-dry summers from once-in-a-decade to every 2–3 years” [Findell 2017: 1]. “Based on these simulations, the conversion of forests to cropland is coincident with much of the upper central US and central Europe experiencing extreme hot, dry summers” [Findell 2017: 6].
Local temperature response to land cover and management change driven by non-radiative processes, Bright et al. 2017
Local temperatures are affected not only by global climatic factors, but also by radiative (albedo) and non-radiative (evapotranspiration and convection) mechanisms related to local vegetation cover. Through evapotranspiration, solar energy is converted to latent heat and released from the planet’s surface, while convection refers to the turbulent mixing of air that dissipates sensible heat. The authors state that while albedo (reflectivity of land surface, which is often lower on forested land) is increasingly accounted for alongside greenhouse gases in climate models, the non-radiative mechanisms are not. However, the evapotranspiration and convection facilitated by vegetation have an important cooling effect and should therefore be included in models to avoid the risk of “promoting land sector policies that may be counter to the aims of mitigation or adaptation” [Bright 2017: 296].
The authors demonstrate that “non-radiative mechanisms dominate the local response in most regions for eight of nine common LCMC perturbations” [Bright 2017: 296]. Land cover and land management changes (LCMC) considered in the study include converting cropland or grassland to deciduous or evergreen forests. The authors found that gains in forest cover increased annual cooling in all but the northernmost latitudes, where the lower albedo of forests compared to grasslands had a warming effect that was stronger than the cooling effect of non-radiative mechanisms. In many regions, including much of Europe, the US, and the tropics, non-radiative cooling dominated albedo effects. “Over annual timescales, forest cover gains result in net cooling for many of the densely populated regions of the planet” [Bright 2017: 298].
Bright et al. conclude that “benchmarking the locally driven LCMC effect to that driven by global forcers (such as CO2) can provide an additional perspective by which to support the valuation of vegetated ecosystems and the local climate regulation services that they provide” [Bright 2017: 301].
Trees, forests and water: Cool insights for a hot world, Ellison et al. 2017
This article (also highlighted in Compendium v2n1) reviews research on the benefits of tree cover in relation to water and energy cycles.
Forests help produce rain. Vegetation releases water vapor through transpiration, increasing atmospheric moisture that is then transported by wind. In fact, “over most of the tropics, air that passes over forests for ten days typically produces at least twice as much rain as air that passes over sparse vegetation” [Ellison 2017: 53]. Forests also release biological particles, such as spores, bacteria and pollen into the atmosphere. Water condenses around these particles, forming raindrops.
In addition to the atmospheric moisture produced by forests that is transported by prevailing winds to generate downwind rain, forests also help transport moisture from the coasts to the interior of continents. According to the biotic pump theory [Makarieva & Gorshkov 2007], evapotranspiration over coastal forests creates low pressure zones that draw in atmospheric moisture from the ocean. This oceanic moisture eventually comes down as rain over land. Deforestation of coastal forests thus reduces this influx of moisture to land, while deforestation anywhere can decrease the reliability of rainfall downwind.
Through shading and evapotranspiration, forests cool the Earth’s surface in tropical and temperate climates. Due to a lower albedo compared to other land cover types at high latitudes, boreal forests potentially contribute to local warming. However, forests also increase cloud cover and thus albedo, higher in the atmosphere. In the absence of vegetation, such as in cities, solar energy remains in the environment in the form of heat, rather than driving evapotranspiration.
Using the sun’s energy, individual trees can transpire hundreds of liters of water per day. This represents a cooling power equivalent to 70 kWh for every 100 L of water transpired (enough to power two average household central air-conditioning units per day) [Ellison 2017: 54].
High-elevation forests have a unique potential to intercept fog and cloud droplets, which boosts tree growth, evapotranspiration, groundwater infiltration, and ultimately contributes up to 75% of catchment runoff.Tree cover can improve water infiltration due to increased organic matter to hold water and the presence of tree roots, which loosen and shade the soil and channel water into the ground. In areas where infiltration rates are greater than transpiration rates, the presence of trees increases groundwater recharge.
All of the aforementioned mechanisms distribute water naturally, hence reducing floods.
Jeannette Christine Armstrong is a Canadian author, educator, artist, and activist, who wrote this article about the traditional decision-making process in Okanagan, called “enowkinwixw,” which demonstrates a great practice of biophilia.
Okanagan, the Penticton Indian reservation in Canada where the author was born and raised, has a very fragile ecosystem. However, the author discovered that this community treats the land differently compared to other communities. They treasure natural resources and believe that the land is the people. They bear in mind that everything they have came from the land and that their every decision has a possibility to destroy the land. Therefore, they not only pay attention to the relationship among human beings but also how human relationships affect the land.
When the author was searching to understand how the community developed this form of interaction, the traditional decision-making process enowkinwixw caught her attention. The model is accomplished with four criteria:
This word demands four things from us: 1) that we solicit the most opposing views; 2) that we seek to understand those views using non-adversarial protocols; 3) that we each agree to be willing to make adjustments in our own interests to accommodate diverse needs expressed; and 4) that we collaboratively commit to support the outcomes [Armstrong 2020: 166].
Through this process, the minority voice is valued as it could point out the most important things that have been ignored. It is how the Okanagan could bring the minority into balance with the majority, minimize the conflicts and bring everyone to work together.
If we begin to think about the minority, about why there is a minority, why there is poverty, then we should be able to find creative ways to meet the needs of the minorities [Armstrong 2020 : 167].
The author believes that this is why the Okanagan community could add preservation of land into the decision-making process. By adopting enowkinwixw, the community respects every opinion from every perspective. Therefore, they train people to speak for different components that make up their existence, such as the children, the elders, and the water; while the author herself was appointed as a land speaker, who thinks and speaks for the land. In this way, the people realize that material things are not as important as the power of the land, which is what sustains them.
This mindset is meant to help the world achieve a harmonic relationship between people and nature.
As I read the first chapter, Wilson brought me far into the forests of the Amazon Basin to encounter canopy-dwelling birds and frogs found nowhere else on Earth; he showed me the life cycle of a tiny moth so specialized that the adult lives only in the fur of the 3-toed sloth, and held up fistfuls of forest soil teeming with tiny life to demonstrate that “the woods were a biological maelstrom of which only the surface could be scanned by the naked eye.”
With his astonishing eye for detail and his fluid prose, he fascinated me, he drew me in, he fed my curiosity…and then he reminded me: “Eliminate just one kind of tree out of hundreds in such a forest, and some of its pollinators, leaf eaters, and wood borers will disappear with it, then various of their parasites and key predators, and perhaps a species of bat or bird that depends on its fruit… and when will the reverberations end?”
Throughout this book, Wilson brought me close to the beauty of living systems across the world. He taught me enough about them to make me curious, and then to make me care; and then, when he showed me how easily that entire system could be disrupted, he had my undivided attention. This kind of understanding and connection, Wilson argues—this human bond with other species—is an essential ingredient in motivating us to change our behaviors in order to slow the rate at which “the wildernesses of the world [are shriveling] into timber leases and threatened nature reserves.”
One of the challenges with building these connections and then acting upon them, Wilson suggests, is that we are programmed to operate in physiological, not ecological, time; that our minds “travel back and forth across hours, days, or at most a hundred years. The forests may all be cut, radiation slowly rise, and winters grow steadily colder, but if the effects are unlikely to become decisive for a few generations, very few people will be stirred to revolt.” But in recent times, our impact on ecosystems has been compressing “ecological time” to align with “physiological time,” and the results are becoming visible in the span of single generations; the massive Florida manatee die-off due to the destruction of their seagrass pastures is but one starkly visible example of the reverberation felt by the loss of a single species from an ecosystem.
In the chapter “The Conservation Ethic”, near the end of the book, Wilson explores the relationship between the human drive to perpetually expand—and the related desire for personal freedom, at least in Western cultures—and the necessity for conscious stewardship of the environment to ensure our ultimate survival—not only our physical survival, but our spiritual survival. He writes “The only way to make a conservation ethic work is to ground it in ultimately selfish reasoning.” In other words, it is likely that wild species and places will be best understood and protected with respect to their perceived value, such as the vast array of plant compounds that have shown promise as anti-cancer compounds, or, as has been increasingly recognized in recent years, for the ability of wild places to improve the emotional and spiritual well-being of the people who spend time there.
This book is even more relevant now than it was when it was released in 1984, and thus has the potential to reach a broader audience today. More people may be ready to hear some of the truths held in this book because now, more than ever before, the human race is seeing, and feeling, the long-term effects of the way we have treated the planet.
(La Symbiose : Structures et Fonctions, Rôle Écologique et Évolutif)
Book review by Ehsan Kayal
What is symbiosis? How is it defined? What does it involve? And how did it come to be? These are some of the questions French Biologist Marc-André Sélosse explores in this book.
It is not simple to define “symbiosis,” which differs between the English and French languages. Sélosse focuses on the narrower French definition, which involves “long-term coexistence of two different organisms throughout their life with reciprocal benefits” [Sélosse 2000: 22]. In English, the term “symbiosis” refers not only to mutualism (reciprocal benefits), but also to parasitism or commensalism (one partner benefit without impacting the other partner). One important aspect of the study of mutualistic symbiosis is exploring reciprocal physiological exchanges that happen between symbionts (organisms involved in symbiosis). Such exchanges are facilitated by morphological changes in one, the other or both partners.
Sélosse offers readers a glimpse of the breadth of symbiotic relationships found in nature. For example, there are bacteria living inside worms around deep-sea vents in the ocean that use chemical energy seeping from the earth to create organic compounds that nourish their worm hosts. The author also highlights the case of lichen, which appears to be a single species, but is actually formed by a symbiosis between fungi and algae. Sélosse focuses especially on symbiosis involving mycorrhiza, which are the “symbiotic organs formed by a root and a fungus” [Sélosse 2000: 140], and on root nodules, the “symbiotic organs of legumes, often around roots, containing bacteria of the Rhizobiaceae family responsible for fixing nitrogen” [Sélosse 2000: 140].
Leguminous plant hosts provide their Rhizobia symbionts with carbon, while protecting these bacteria from harmful oxygen, in exchange for nitrogen. The efficiency of these symbiosis is such that the “rhizobiaceae-containing root nodules fix as much atmospheric nitrogen per year as the fertilizer industry” [Sélosse 2000: 54]. Similarly, plants with mycorrhizal associations supply their symbionts with carbon in exchange for water, phosphate and other nutrients furnished by the fungi. Studies have linked “an increasing diversity of mycorrhiza” with “increasing diversity of plants” [Sélosse 2000: 67].
Symbiosis can short-circuit the mineralization-immobilization cycle (conversion of inorganic compounds to organic compounds by micro-organisms or plants) by bringing together partners that thrive on each other’s by-products. This results in a “concentration of resources” [Sélosse 2000: 49] that allows organisms to conquer new ecological niches as exemplified by lichen and corals, but also playing a key role in the evolution of the soil that promotes vegetation successions [Sélosse 2000: 60]. For example, lichen is the first to colonize bare rock, where it produces citric acid and holds water, slowing altering the rock in the early stage of soil creation. Rhizobial symbioses then pitch in by fixing nitrogen, otherwise absent in a rocky substrate.
The book also flips some of the common understandings of organismal biology. For instance, many herbivores lack digestive enzymes to break down plant material. Rather, they feed on the population of microorganisms of their rumen, or the “pocket situated upstream of the stomach in ruminants that harbors a symbiotic microflora” [Sélosse 2000: 141]). The rumen is a large organ, whose volume can represent “8 to 15% of total body weight” [Sélosse 2000: 32]. That makes those microorganisms (protists, fungi and bacteria) the “true” herbivores, while the ruminants are secondary consumers.
The older traces of symbiosis predate the origin of the eukaryotic cell (cell with a nucleus enclosed within a nuclear envelope), where the mitochondrion (the energy-producing organelle of the cell) are descendants of a likely unique endosymbiotic event occurring some two billion years ago. In that event, an anaerobic archaean (third kingdom of life composed of unicellular organisms lacking a nucleus; the other two kingdoms are eukaryotes and bacteria) captured a facultative aerobic type of bacteria, which became mitochondria. Similarly, the acquisition by some eukaryotic cells of a chloroplast originating from cyanobacteria gave way to photosynthetic plants.
In some sense, “no organism lives alone, and each carries a symbiotic cortege without which one cannot understand neither the physiology nor the ecological success of the organism” [Sélosse 2000: 134].
Symbiosis: Structure and Functions, Ecological and Evolutionary Role, Sélosse 2000
(La Symbiose : Structures et Fonctions, Rôle Écologique et Évolutif)
Book review by Ehsan Kayal
What is symbiosis? How is it defined? What does it involve? And how did it come to be? These are some of the questions French Biologist Marc-André Sélosse explores in this book.
It is not simple to define “symbiosis,” which differs between the English and French languages. Sélosse focuses on the narrower French definition, which involves “long-term coexistence of two different organisms throughout their life with reciprocal benefits” [Sélosse 2000: 22]. In English, the term “symbiosis” refers not only to mutualism (reciprocal benefits), but also to parasitism or commensalism (one partner benefit without impacting the other partner). One important aspect of the study of mutualistic symbiosis is exploring reciprocal physiological exchanges that happen between symbionts (organisms involved in symbiosis). Such exchanges are facilitated by morphological changes in one, the other or both partners.
Sélosse offers readers a glimpse of the breadth of symbiotic relationships found in nature. For example, there are bacteria living inside worms around deep-sea vents in the ocean that use chemical energy seeping from the earth to create organic compounds that nourish their worm hosts. The author also highlights the case of lichen, which appears to be a single species, but is actually formed by a symbiosis between fungi and algae. Sélosse focuses especially on symbiosis involving mycorrhiza, which are the “symbiotic organs formed by a root and a fungus” [Sélosse 2000: 140], and on root nodules, the “symbiotic organs of legumes, often around roots, containing bacteria of the Rhizobiaceae family responsible for fixing nitrogen” [Sélosse 2000: 140].
Leguminous plant hosts provide their Rhizobia symbionts with carbon, while protecting these bacteria from harmful oxygen, in exchange for nitrogen. The efficiency of these symbiosis is such that the “rhizobiaceae-containing root nodules fix as much atmospheric nitrogen per year as the fertilizer industry” [Sélosse 2000: 54]. Similarly, plants with mycorrhizal associations supply their symbionts with carbon in exchange for water, phosphate and other nutrients furnished by the fungi. Studies have linked “an increasing diversity of mycorrhiza” with “increasing diversity of plants” [Sélosse 2000: 67].
Symbiosis can short-circuit the mineralization-immobilization cycle (conversion of inorganic compounds to organic compounds by micro-organisms or plants) by bringing together partners that thrive on each other’s by-products. This results in a “concentration of resources” [Sélosse 2000: 49] that allows organisms to conquer new ecological niches as exemplified by lichen and corals, but also playing a key role in the evolution of the soil that promotes vegetation successions [Sélosse 2000: 60]. For example, lichen is the first to colonize bare rock, where it produces citric acid and holds water, slowing altering the rock in the early stage of soil creation. Rhizobial symbioses then pitch in by fixing nitrogen, otherwise absent in a rocky substrate.
The book also flips some of the common understandings of organismal biology. For instance, many herbivores lack digestive enzymes to break down plant material. Rather, they feed on the population of microorganisms of their rumen, or the “pocket situated upstream of the stomach in ruminants that harbors a symbiotic microflora” [Sélosse 2000: 141]). The rumen is a large organ, whose volume can represent “8 to 15% of total body weight” [Sélosse 2000: 32]. That makes those microorganisms (protists, fungi and bacteria) the “true” herbivores, while the ruminants are secondary consumers.
The older traces of symbiosis predate the origin of the eukaryotic cell (cell with a nucleus enclosed within a nuclear envelope), where the mitochondrion (the energy-producing organelle of the cell) are descendants of a likely unique endosymbiotic event occurring some two billion years ago. In that event, an anaerobic archaean (third kingdom of life composed of unicellular organisms lacking a nucleus; the other two kingdoms are eukaryotes and bacteria) captured a facultative aerobic type of bacteria, which became mitochondria. Similarly, the acquisition by some eukaryotic cells of a chloroplast originating from cyanobacteria gave way to photosynthetic plants.
In some sense, “no organism lives alone, and each carries a symbiotic cortege without which one cannot understand neither the physiology nor the ecological success of the organism” [Sélosse 2000: 134].
Biophilia: the human bond with other species, Wilson 1984
A book review by Rachel West
As I read the first chapter, Wilson brought me far into the forests of the Amazon Basin to encounter canopy-dwelling birds and frogs found nowhere else on Earth; he showed me the life cycle of a tiny moth so specialized that the adult lives only in the fur of the 3-toed sloth, and held up fistfuls of forest soil teeming with tiny life to demonstrate that “the woods were a biological maelstrom of which only the surface could be scanned by the naked eye.”
With his astonishing eye for detail and his fluid prose, he fascinated me, he drew me in, he fed my curiosity…and then he reminded me: “Eliminate just one kind of tree out of hundreds in such a forest, and some of its pollinators, leaf eaters, and wood borers will disappear with it, then various of their parasites and key predators, and perhaps a species of bat or bird that depends on its fruit… and when will the reverberations end?”
Throughout this book, Wilson brought me close to the beauty of living systems across the world. He taught me enough about them to make me curious, and then to make me care; and then, when he showed me how easily that entire system could be disrupted, he had my undivided attention. This kind of understanding and connection, Wilson argues—this human bond with other species—is an essential ingredient in motivating us to change our behaviors in order to slow the rate at which “the wildernesses of the world [are shriveling] into timber leases and threatened nature reserves.”
One of the challenges with building these connections and then acting upon them, Wilson suggests, is that we are programmed to operate in physiological, not ecological, time; that our minds “travel back and forth across hours, days, or at most a hundred years. The forests may all be cut, radiation slowly rise, and winters grow steadily colder, but if the effects are unlikely to become decisive for a few generations, very few people will be stirred to revolt.” But in recent times, our impact on ecosystems has been compressing “ecological time” to align with “physiological time,” and the results are becoming visible in the span of single generations; the massive Florida manatee die-off due to the destruction of their seagrass pastures is but one starkly visible example of the reverberation felt by the loss of a single species from an ecosystem.
In the chapter “The Conservation Ethic”, near the end of the book, Wilson explores the relationship between the human drive to perpetually expand—and the related desire for personal freedom, at least in Western cultures—and the necessity for conscious stewardship of the environment to ensure our ultimate survival—not only our physical survival, but our spiritual survival. He writes “The only way to make a conservation ethic work is to ground it in ultimately selfish reasoning.” In other words, it is likely that wild species and places will be best understood and protected with respect to their perceived value, such as the vast array of plant compounds that have shown promise as anti-cancer compounds, or, as has been increasingly recognized in recent years, for the ability of wild places to improve the emotional and spiritual well-being of the people who spend time there.
This book is even more relevant now than it was when it was released in 1984, and thus has the potential to reach a broader audience today. More people may be ready to hear some of the truths held in this book because now, more than ever before, the human race is seeing, and feeling, the long-term effects of the way we have treated the planet.
An Okanagan Worldview of Society, Armstrong 2020
Jeannette Christine Armstrong is a Canadian author, educator, artist, and activist, who wrote this article about the traditional decision-making process in Okanagan, called “enowkinwixw,” which demonstrates a great practice of biophilia.
Okanagan, the Penticton Indian reservation in Canada where the author was born and raised, has a very fragile ecosystem. However, the author discovered that this community treats the land differently compared to other communities. They treasure natural resources and believe that the land is the people. They bear in mind that everything they have came from the land and that their every decision has a possibility to destroy the land. Therefore, they not only pay attention to the relationship among human beings but also how human relationships affect the land.
When the author was searching to understand how the community developed this form of interaction, the traditional decision-making process enowkinwixw caught her attention. The model is accomplished with four criteria:
This word demands four things from us: 1) that we solicit the most opposing views; 2) that we seek to understand those views using non-adversarial protocols; 3) that we each agree to be willing to make adjustments in our own interests to accommodate diverse needs expressed; and 4) that we collaboratively commit to support the outcomes [Armstrong 2020: 166].
Through this process, the minority voice is valued as it could point out the most important things that have been ignored. It is how the Okanagan could bring the minority into balance with the majority, minimize the conflicts and bring everyone to work together.
If we begin to think about the minority, about why there is a minority, why there is poverty, then we should be able to find creative ways to meet the needs of the minorities [Armstrong 2020 : 167].
The author believes that this is why the Okanagan community could add preservation of land into the decision-making process. By adopting enowkinwixw, the community respects every opinion from every perspective. Therefore, they train people to speak for different components that make up their existence, such as the children, the elders, and the water; while the author herself was appointed as a land speaker, who thinks and speaks for the land. In this way, the people realize that material things are not as important as the power of the land, which is what sustains them.
This mindset is meant to help the world achieve a harmonic relationship between people and nature.
Well-planned tree-planting projects are an important component of global efforts to improve ecological and human well-being. But tree planting becomes problematic when it is promoted as a simple, silver bullet solution and overshadows other actions that have greater potential for addressing the drivers of specific environmental problems, such as taking bold and rapid steps to reduce deforestation and greenhouse gas emissions [Holl 2020: 580].
Some of the pitfalls to avoid in tree planting initiatives, according to the authors, include:
Use of non-native species, which does not result in a true forest and can result in ground water depletion in arid environments.
Planting trees in historic grasslands and savannas, harming those native ecosystems and species.
Abandoning trees after they are planted, which can result in high mortality due to insufficient water for developing saplings, being shaded out by faster growing herbaceous plants, grazing, or being re-cleared.
Planting trees in agricultural land, which risks pushing crop production into native forest land, which is then deforested.
The authors insist that reforestation takes careful planning, stakeholder engagement, clear goal-setting, and long-term monitoring and adaptive management of planted tree stands to ensure their survival. Above all, existing mature, native forests should be preserved.
The authors insist that reforestation takes careful planning, stakeholder engagement, clear goal-setting, and long-term monitoring and adaptive management of planted tree stands to ensure their survival. Above all, existing mature, native forests should be preserved.
The first priority to increase the overall number of trees on the planet must be to reduce the current rapid rate of forest clearing and degradation in many areas of the world. The immediate response of the G7 nations to the 2019 Amazon fires was to offer funding to reforest these areas, rather than to address the core issues of enforcing laws, protecting lands of indigenous people, and providing incentives to landowners to maintain forest cover. The simplistic assumption that tree planting can immediately compensate for clearing intact forest is not uncommon. Nonetheless, a large body of literature shows that even the best-planned restoration projects rarely fully recover the biodiversity of intact forest, owing to a lack of sources of forest-dependent flora and fauna in deforested landscapes, as well as degraded abiotic conditions resulting from anthropogenic activities [Holl 2020: 581].
As one of China’s major cities, Shanghai’s natural sub-ecosystem[5] has suffered drastic damage due to human activities and urbanization. Although urban re-naturalization has gained attention from city leaders, urban tree planting has largely consisted of two methods with limited ecological potential. One favors fast-growing monocultures to produce timber products and other benefits, while the other approach is to plant non-native species for decorative purposes. The authors believe the restoration progress of the natural sub-ecosystem could be further improved by adopting the “near-natural” method based on the concepts of potential natural vegetation[6] and ecological succession.
The near-natural forest uses all native species and aims to create a complex structure with high biodiversity, high biomass and multiplayer canopies. It was adopted successfully in many countries, but the authors thought long-term studies of these forests were lacking. Therefore, they conducted a 10-year study at a near-natural forest established in 2000 in Pudong New Area of Shanghai to investigate the effectiveness of the forest in providing ecological benefits.
Results showed that the near-natural forest had higher sustainability value than artificial (“even-aged, managed”) forest in Shanghai based on its ecological and economic benefits. The high tree density and multiple vertical structures of the forest improved the air quality and soil fertility and decreased the concentrations of air bacteria and dust. It also had a much lower planting and maintenance cost than artificial traditional methods. Although the near-natural forest could not transcend the benefits of the natural forest, the study successfully proved its important role in urban re-naturalization by bridging the difference between the artificial and natural forests.
Results showed that the near-natural forest had higher sustainability value than artificial (“even-aged, managed”) forest in Shanghai based on its ecological and economic benefits
Over the course of the study, authors discovered a potential limitation of the approach, at least in its application in Shanghai. They observed high evergreen seedlings mortality, attributable to over-exposure to sunlight. Therefore, in subsequent plantings in 2003 and 2004, the authors modified the approach to optimize it to local conditions:
The key to the new method is to create a mixed deciduous–evergreen community by simultaneously planting shade-tolerant evergreen broad-leaved species and light-demanding deciduous broad-leaved species, but using smaller individuals for the former and larger individuals for the latter to form a multilayer vegetation structure. The shade-tolerant evergreen species benefit from the rapid growth of the light-demanding deciduous species, which offer shade and nutrients in the form of litter layer-based fertilizer, improving the soil for the evergreen species [Guo 2015: 5].
Overall, they suggest that the near-natural forest is a very sustainable method to be applied in Shanghai.
This book draws lessons from ecological succession theory to inform ecological restoration, stating that: “restoration is fundamentally the management of succession” [Walker 2007: vi]. The latter is the natural process by which plants first colonize “new” land (post landslide, glacial retreat or volcanic eruption, for example) or degraded land, and over time develop into mature ecosystems through a series of changing plant communities. Ecological restoration is a human-led initiative to restore functioning ecosystems, or at least vegetation, on land degraded through human activity. The ultimate goal of restoration is to “establish a self-sufficient ecosystem that requires minimal or no continuing human inputs in order to provide a continuing supply of goods and services” [Hobbs 2007: 177].
Effective ecosystem restoration requires ecological knowledge. Likewise, the outcomes of such projects demonstrate our comprehension, or lack thereof, of ecological concepts: “Restoration is the acid test of our ability to understand not only how ecosystems are assembled and held together but also how they change over time” [Walker 2007: vi]. The authors contend, however, that restoration projects are more often guided by engineering, horticulture, and agronomy than by ecology. Aiming to clarify the ways in which ecological succession theory can and should inform restoration, this book poses the question: “What is the minimum amount of biophysical and successional information needed to restore a specific landscape or area” [Walker 2007b: 2]?
Succession comprises many ecological processes that underpin all ecological restoration and ecological restoration is a manipulation of these processes to achieve its goals. This means it is essential to understand how succession operates, and when and how to manipulate it [Prach 2007: 121].
Restoration can explicitly embrace a hands-off approach, where land is simply left to repair itself through natural ecological succession. On the other hand, understanding the successional process allows manipulation of various stages to speed up the process. For example, in the first stage of primary succession “winds deposit dust, pollen, seeds, and insects crucial to reducing infertility” [del Moral 2007: 23], on bare, inhospitable ground. Tough pioneer plants are able to establish then create shade, trap sediment, and deposit organic matter when they die, creating slightly better conditions for the next wave of colonizing plants. To mimic this first stage of site “amelioration”, the site can be physically manipulated by reshaping the ground for improved drainage or adding organic matter, for example.
Biological manipulation involves sowing or planting local/native varieties of later successional species that may not be otherwise present in the area due to human transformation of the broader landscape. While earlier successional species tend to have small, easily transported seeds, the later successional species (such as large canopy trees) that are often the target of restoration efforts often have large, less mobile seeds. Thus, if those plants are not present in the immediate environment as seed stock, they may never establish in the restored site without human assistance.
Anthropogenic environmental change and global dispersal of a wide variety of species outside their native ranges has expanded the range of “cosmopolitan,” non-native species and shrunk the range of regional and endemic species. “This replacement of specific native forms by generalist non-natives in space and time has mixed the taxonomic composition of once disparate biotas, an occurrence termed ‘biotic homogenization’” [Olden 2004: 18].
The authors explore the effect of this “global erosion of regional distinctiveness” [Olden 2004: 18] at three levels: Genetic homogenization reduces genetic variability within species or among populations of species, while taxonomic homogenization reduces distinctiveness among communities. Functional homogenization refers to a reduction of functional traits within an ecosystem. The identity of species making up a community, along with their respective functional traits, determines “ecosystem functions (such as nutrient retention or energy flow)” [Olden 2004: 20], so that narrowing species compositions risks diminishing ecosystem function.
A decrease in functional diversity might reduce overall community and ecosystem functioning, stability and resistance to environmental change by simply narrowing the available range of species-specific responses. Consider a severe drought that strongly affects a subset of species in a community that has (or lacks) a particular suite of functional traits. Historical communities, with much greater breadth in functional space, should exhibit higher resistance or resilience when compared with homogenized communities [Olden 2004: 20].
Genetic homogenization occurs when two distinct locally adapted populations of the same species interbreed. It also occurs when a single variety (such as captive fish bred in a central location) are released in many places to replenish dwindling native stocks. While such mixing has the potential to increase species diversity, this outcome is not assured.
Intraspecific hybridization can homogenize the unique characteristics of geographically distinct populations, as well as compromise the fitness of individuals by disrupting local adaptations [Olden 2004: 19].
The ecological, economic, and social damage of human-mediated dispersal of species into new regions, where they possess the ability to naturalize (become self-sustaining their new homeland), is one of the defining features of the Anthropocene Epoch. Globally, human activity has led to the naturalization of nearly 13,168 plant species (equal in size to the native European flora). The results from this research provide a baseline for monitoring global changes in biodiversity while highlighting the immediate action that has to be taken to comprehend and determine the spread of alien species on an international scale.
The ecological, economic, and social damage of human-mediated dispersal of species into new regions, where they possess the ability to naturalize (become self-sustaining their new homeland), is one of the defining features of the Anthropocene Epoch.
At least 3.9% of all currently known vascular plant species have become naturalized outside their natural ranges as a result of human activity. With the continued practice of international traffic and trade and globalization, the likelihood of more and more species being introduced and getting naturalized outside their native range is high.
To assess the accumulation of naturalized species in each continent as well as which continents have been the major donors of alien naturalized plant species globally, the researchers used a novel database, Global Naturalized Alien Flora (GloNAF), in addition to the data on the origin of naturalized species and estimates of the number of native species per continent. They found that when not taking into account the differences in total area, North America has accumulated the highest number of naturalized species (n=5,958). However, when considering the difference in total area, Australasia (a region comprising Australia, New Zealand, and neighboring islands) was found to have more extra-continental species than North America.
One possible explanation is that Australia’s long biogeographical isolation and drying climate have resulted in a native flora that is phylogenetically distinct, but not well-adapted to exploit the novel habitats created by European settlers [van Kleunen 2015: 101].
The major donors of alien species are Europe and temperate Asia, while North America is also a significant donor.
The widespread distribution of plants outside of their native range due to human activity is a significant yet underrecognized cause of global insect decline, according to this article. To illuminate the issue, the authors: “examine the evidence for and against the hypothesis that long term changes in the species composition of plant assemblages have contributed to local and global declines in the abundance and diversity of the insect communities dependent upon those assemblages” [Tallamy 2020: 2].
To be sure, insect conservationists have long noted the importance of habitat containing appropriate native host plants, but the widespread replacement of native host plants with non-native species has yet to penetrate the growing literature on insect declines in any meaningful way [Tallamy 2020: 1].
It is not simply the absence of native plants harms plant-eating insects, however, but also the presence of non-natives. While some insects feed successfully on non-native plants, this is the minority. Most either avoid non-native plants, or do use them and are killed or malnourished by doing so. For example,
Swallowworts (Vincetoxicum spp.) are confamilials of milkweeds (Asclepias spp.) and have become invasive in parts of the northeastern United States. Similar phytochemistry between swallowworts and milkweeds can lead monarch butterflies (Danaus plexxipus) and milkweed beetles (Chrysochus auratus) to fatally mistake these chemically protected plants as hosts. The degree to which Vincetoxicum act as ecological traps for these taxa is likely to become more pronounced as the plants become dominant and displace milkweeds in the landscape [Tallamy 2020: 3].
Species that share a particular environment over hundreds or thousands of years evolve in relation to one another. For plant-eating insects, adapting to certain plants meant developing “traits to detect and tolerate plant defenses over time” [Tallamy 2020: 2]. Most herbivorous insects adapted to only a particular set of plants, specializing in feeding on those plant hosts.
The diet of most insects is constrained to a single plant family in any one habitat or location, with dietary specialization even narrower both in many temperate lineages and hyper-diverse tropical lineages. In fact, diet specialization increases with decreasing latitudes, concurrent with theories of increased plant and animal diversity in the tropics [Tallamy 2020: 2].
When native plants are displaced in the landscape by non-native species, phytophagous [plant-eating] insects typically do not recognize the novel host for feeding or oviposition [egg laying], or may be unable to overcome novel plant defenses. The concurrent loss of native plant hosts and dominance of non-native plants can lead to local extirpation of phytophagous insects and thus to changes in the composition and structure of local food webs [Tallamy 2020: 2].
The most likely successful substitute for a native plant is a non-native plant in the same genus or family.
Non-native congeners [members of the same genus] or confamilials [members of the same family] that are similar in foliar chemistry and nutrition, phenology, and morphology, may occasionally serve as novel hosts for herbivorous insects and support higher diversity and abundance than non-native, non-congeners. However, novel use of congeners may increase larval mortality, extend development or pupation time, reduce biomass, and reduce fitness compared to that of native hosts [Tallamy 2020: 3].
The narrower the native plant diet an insect species has, the less likely to tolerate novel, non-native food sources. However, there are more species of specialist insects than of generalists, meaning a larger proportion of susceptible species. Adaptability to exotic host plants also depends on an insects’ feeding habits.
Insects with chewing (mandibulate) mouthparts are typically more susceptible to defensive secondary metabolites contained in leaf vacuoles than are insects with sucking (haustelate) mouthparts that tap into poorly defended xylem or phloem fluids. Thus, sucking insects find novel non-native plants to be acceptable hosts more often than do chewing species [Tallamy 2020: 4].
Considering that there are more than 4.5 times as many mandibulate insect herbivores as haustelate species, there is reason for concern when non-native plants replace native hosts; the largest guild of insect herbivores is also the most vulnerable to non-native plants and the most valuable to insectivores [Tallamy 2020: 5].
“The dispersal and spread of invasive plants has been driven by global trade networks and colonialism” [Tallamy 2020: 6] and, more specifically, from agroforestry, forestry, agriculture, and horticulture.
Although plants have always distributed themselves around the globe, the increased temporal and spatial mobility of humans has resulted in an extraordinary increase in the rate of plant movements and most species’ introductions have happened in the last 200 years. Habitat is rapidly being converted from coevolved native ecosystems into novel assemblages of plants and animals, making the conversion of native plant communities into plant assemblages dominated by non-native species one of the most ubiquitous threats to biodiversity today. The introduction of non-native plants has completely transformed the composition of present-day plant communities in both natural and human-dominated ecosystems around the globe and the magnitude of introductions is staggering. An estimated 13,168 plant species (about 3.9% of global vascular flora) have been introduced and naturalized beyond their native ranges as a result of human activity [Tallamy 2020: 6].
This study examined whether non-native plants in residential Washington DC limited the presence of the Carolina chickadee, a local breeding insectivore.
We predicted that areas with more native plants would support more chickadees, and chickadees would forage more often in the most insect-producing native plants [Narango 2017: 43].
The authors had also considered the possibility that non-native plants could promote increases in other food items (e.g. non-native arthropods), keeping overall prey biomass similar between native and non-native plants. What they found, though, affirmed their prediction: native plants produce more caterpillars, which in turn support more chickadees. In fact, the birds avoided foraging in non-native plants, including non-native species of the same tree genera: the chickadees preferred maples native to the eastern US compared to European-origin maples.
Native plants produce more caterpillars, which in turn support more chickadees.
Native plants were more likely to host a higher biomass of caterpillars compared to non-native plants, and chickadees strongly preferred to forage in native plants that supported the most caterpillars. In addition, chickadees were less likely to breed in yards as the dominance of non-native plants increased [Narango 2017: 42].
Also unique to our study is that we measured the probability of caterpillar occurrence between congeneric species (e.g. native vs. non-native Acer [maple]). This is particularly important considering the popularity and invasive qualities of congeneric species in this region such as Acer platanoides and Quercus acutissima. Although non-native congeners support more caterpillars in comparison to plants unrelated to any native species, congeners had a 47% (CI: 34%–59%) lower probability of having caterpillars compared to native species [Narango 2017: 47].
The authors state that local insects are adapted to local plants, presumably due to their shared co-evolutionary history.
This occurs in part because herbivorous insects have adapted to circumvent the phytochemical defenses of particular plant lineages, resulting in a radiation of specialized plant-insect associations. During urban conversion, native plants are replaced by non-native species with novel chemical, physical, and phenological features for which native herbivorous arthropods have few physiological or behavioral adaptations [Narango 2017: 42].
In this study, the insect and bird populations of six pairs of suburban yards were measured. Each pair contained one conventionally landscaped yard containing native canopy trees and a mixture of native and non-native shrubs, grasses and understory trees; and one yard with native species only (canopy, understory, shrub and grasses). The level of plant diversity was comparable between each of the pair; only the proportion of native species differed. The authors found that:
Avian abundance, diversity, richness, and biomass (particularly bird species of conservation concern) were all greater on native properties. Native nesting birds that are mostly dependent on insect populations to feed their young were more abundant on native properties. Lepidoptera [butterfly and moth species] abundance and diversity were also higher on native properties, suggesting that food availability might account for the differences detected in the bird communities between native and conventionally landscaped sites [Burghardt 2008: 223].
These results support the authors’ hypothesis based on an understanding of the co-evolutionary roots of species interactions.
Theory backed by decades of empirical evidence predicts that up to 90% of all species of insect herbivores can successfully reproduce only on plant lineages with which they have shared an evolutionary history [Burghardt 2008: 220].
This research evaluates the impact of the invasion of non-native plants in the distribution of lepidopteran (butterfly, skipper, and moth) communities. The authors assert that although the introduction of non-native plants has not resulted in a “global extinction”, they have had a considerable impact on how ecosystems function—they often result in significant bottom-up reductions of energy available in local food webs.
The experiment established four gardens near mature woodlots containing most, if not all, of the native species planted within the treatment. The richness and abundance were then compared for lepidopteran communities found on native versus corresponding non-native congener[4] species of 13 woody plant genera. For example, the genus Acer (maple) was selected for this study because the native and non-native maples were widespread in that area. In separate plots, the researchers also compared native plants and unrelated (non-congeneric) non-native plants for lepidopteran richness and abundance.
The study found that lepidopterans suffer from the replacement of native plants by non-natives, especially when those non-natives are unrelated to any native plant species. The authors explain that “insect herbivores adapted to the chemical challenges [toxic plant defenses] of particular native hosts may be able to adopt a novel plant species as a host if its phytochemistry is sufficiently similar to the original hosts” [Burghardt 2010: 10]. Over the two-year study, lepidopteran abundance and richness were depressed both on congener and (unrelated) non-congener non-native plants, but especially on the latter.
The study found that lepidopterans suffer from the replacement of native plants by non-natives, especially when those non-natives are unrelated to any native plant species.
Specialist lepidopteran species, which require specific diet and habitat conditions to survive, fared worse on non-natives than did generalists, which can eat a variety of foods and survive in many different habitats. The authors note, for example, that “geographically novel congeners were acceptable hosts to less than half of the generalists and only one fourth of the specialists that we found on native congeners in 2009” [Burghardt 2010: 11]. Only 7% of specialist species used non-congener non-natives as hosts.
The authors argue that the loss of lepidopteran diversity and abundance due to the displacement of native plant species with non-natives can ripple up the food chain, reducing diversity at higher trophic levels. Reduced diversity leads to lower ecosystem productivity and stability, thus disrupting the whole system.
The authors argue that the loss of lepidopteran diversity and abundance due to the displacement of native plant species with non-natives can ripple up the food chain, reducing diversity at higher trophic levels. Reduced diversity leads to lower ecosystem productivity and stability, thus disrupting the whole system.
Because insect herbivores are near the hub of most terrestrial food webs, comprising essential food stuffs for an incredible diversity of insect predators and parasitoids, spiders, amphibians, lizards, rodents, bats, birds, and even higher predators such as foxes and bears, it is particularly important to understand changes wrought by non-native plants on this critical taxon [Burghardt 2010: 13].
This review explores the question of whether plant-plant interactions drive evolutionary changes. “If such evolution is common, plant communities are not random assemblages of species.” The topic is under-studied compared to plant interactions with other groups.
Research on plant–consumer, plant–pollinator and plant–disperser interactions has been central to understanding the complex mutualistic and co-dependent interactions among species that structure communities. However, with some notable exceptions, interactions among plants have not been emphasized as processes that contribute to selection and evolution [Thorpe 2011: 730].
“The simplest interactions among plants are direct interactions, such as facilitation, resource competition and allelopathy” [Thorpe 2011: 731]. Facilitation is when one plant protects an adjacent plant, such as from drought and heat by providing shade, for example, or from browsing by being thorny or toxic to herbivores and surrounding the facilitated plant. Allelopathy refers to plants’ release of toxic substances that suppress the growth of another organism, including other plants. In natural communities, any given plant may be interacting with several different plants at the same time.
In natural communities, any given plant may be interacting with several different plants at the same time.
Competition for sunlight, water, and nutrients drives niche differentiation, or the carving out by species of particular spaces or timing within an ecosystem to obtain a share of limited resources. “The exceptionally rich body of ecological literature on the niche is based in part on the idea that competition can drive the evolution of niche differentiation, thus allowing species to coexist” [Thorpe 2011: 732].
Thorpe et al. refer to an example from a 1976 article by Parrish & Bazzaz , who “found that resource partitioning, as estimated from spatial overlap among root systems, was higher in stable prairie communities with a long community history than in early successional old-field communities composed of species without a common history” [Thorpe 2011: 731]. In other words, plants with a long coexistence history more efficiently divvy up resources than do species lacking a common community history.
The primary hypothesis for positive diversity–ecosystem function relationships has been niche ‘complementarity’, the idea that different species or functional groups occupy niches different enough from each other to more fully utilize resources or space, increasing and stabilizing productivity, and making it more difficult for other species to enter the community [Thorpe 2011: 733].
The authors are somewhat inconclusive, however, about what drives niche complementarity (resource partitioning).
We do not yet know whether complementarity is produced by interactions causing evolutionary shifts in niche space (and thus coexistence and more complete resource use) or by sorting of the existing species pool [Thorpe 2011: 733].
Plants can also adapt to one another’s allelopathic substances over time, a fact that contributes to the argument that plant-plant interactions produce evolutionary changes. “Recent experiments raise the possibility that some invaders may exude allelochemicals that are relatively ineffective against neighbors in natural communities, but highly inhibitory to plants in invaded communities” [Thorpe 2011: 734].
This article emphasizes the importance of species interactions as drivers of ecosystem function.
The classic conservation approach is to set aside national parks or to target specific species for protection, based on their rarity or endangered status. However, these approaches can have trade-offs for non-target species, while also potentially failing to protect ecosystem function. The authors, therefore, suggest that species interactions based on their functional significance should be the main focus on conservation efforts.
We propose that a shift in focus from species to interaction networks is necessary to achieve pressing conservation management and restoration ecology goals of conserving biodiversity, ecosystem processes and ultimately landscape-scale delivery of ecosystem services [Harvey 2017: 371].
Species depend on many other species in their communities, either directly or indirectly. An example of indirect dependence is the Phengaris arion butterfly’s need for European rabbits. The butterfly uses ant nests made in the open areas supplied by rabbit grazing for development of its larvae. Thus, no rabbits means no ants, which means no Phengaris arion.
Focusing on species interactions is more meaningful even than measuring species richness (the number of different species), because interactions can disappear – even if both species are present – if either group’s abundance has significantly dropped. The authors offer the example of 59 regionally extinct lepidoptera (butterfly and moth) species of central Europe. Eight of these extinctions were associated with the loss of particular host plant species, which actually occurred after the lepidoptera went extinct.
Focusing on species interactions is more meaningful even than measuring species richness (the number of different species), because interactions can disappear – even if both species are present – if either group’s abundance has significantly dropped.
Thus, strong declines of host plants can have cascading extinction effects on higher trophic levels before the plants actually go extinct, illustrating that interactions can be lost before any actual decline in species richness (plants persisted at low abundance). This illustrates that preserving keystone interactions, rather than species, can be a proactive way to maintain ecosystem integrity in the face of global change instead of allocating resources to already endangered species [Harvey 2017: 372].
There is interdependence among species even between neighboring ecosystems. For example, a manta ray species in the Palmyra Atoll south of Hawaii depends on two species of native trees to maintain its ocean plankton diet. When these trees were replaced with cultivated coconut palms, marine-foraging birds no longer nested on that shore, depriving the coastal waters of the nitrogen runoff from their guano, which had been feeding the plankton population.
The authors recommend that “the main lever to restore or conserve ecological network structure and stability is the management of spatial configuration” [Harvey 2017: 377]. Reflecting on the Palmyra Atoll, for example, it’s clear that a marine conservation plan would be incomplete without considering the nutrient flow from the tree-bird interactions on land.
Ecological restoration aims to recreate lost or degraded ecological communities. However, “community” has been a difficult concept to define – should the definition stress dominant species, species interactions, or a subset of strongly interacting species? These authors propose defining community on the basis of co-evolutionary relationships among species.
We propose that an Evolutionary Community is conceptualized as a unique grouping of species, which occur in a given geographic area and are connected by interspecific and abiotic interactions that have evolved over time [Steen 2017: 1021].
By treating communities “as entities that have formed over evolutionary time; this [Evolutionary Community] concept allows for a philosophical platform to help us understand what many conservation and restoration efforts are trying to accomplish” [Steen 2017: 1031]. That is, it offers a way to conceptualize the end goal of a restoration project.A particular evolutionary community could be recreated by assembling the constituent species, resulting in the ecological interactions among the species resuming as before.
What processes cause a group of species to cohere into a community? We argue that the parts of Evolutionary Communities are bound together by interspecific interactions in a shared biotic and abiotic environment, which promote co-evolution and community structure and dynamics. For example, longleaf pine trees are conduits for lightning strikes that ignite a highly flammable understory, often including dropped longleaf pine needles. The resulting ground fires are necessary for reproduction of other species and maintain habitat suitable for others (e.g., gopher tortoises). Gopher tortoises, through the process of burrow creation, provide structure important to other species. The establishment of one or more of the species listed above facilitated the persistence of additional species [Steen 2017: 1025].
Likewise, the demise of one species will negatively affect, or even cause the demise of, other species that depend on it. Thus, the reason to preserve or recreate an integral community is to support the interdependent component species, each of which in turn support the community as a whole.
This study uses the concept of Potential Natural Vegetation (PNV), developed in the mid-1900s by German botanist Reinhold Tüxen. Described by the authors as “one of the most successful novelties in vegetation science over the last decades” [Liu 2009: 1313], PNV can be defined as a projection of the natural vegetation that would exist in a given area in the absence of human interference.
“By showing the relationships between environmental variables and vegetation types, maps of the PNV are an important instrument in the study and planning of the environment, and act as decision-support tools for the solutions to environmental issues” [Liu 2009: 1313]. Such maps are informed by studying remnant natural (old growth) vegetation in the area and site observations of the area to be mapped.
Computer modeling can be used to predict “the geographic distribution of vegetation composition across a landscape from mapped environmental variables, such as climate, soils, and geology. When a predictive vegetation modeling is calibrated using observation of vegetation composition taken from mature or ‘climax’ vegetation stands, then potential natural vegetation is portrayed in a predictive map” [Liu 209: 1314].
Focusing on northeastern China, the study identified 16 vegetation types in the region, along with the environmental factors influencing their distribution. Climatic factors included: mean annual temperature, mean temperature of the coldest month, relative humidity, and potential evapotranspiration rate. Topographical factors were elevation and slope.
“Generally, as the elevation increases, the change of temperature and moisture leads to the obvious differentiation phenomenon in vegetation vertical zones. Slope is related to the hydrology (overland and subsurface flow velocity and runoff rate) and potential soil moisture and soil development of a habitat” [Liu 2009: 1315].
They compared the map created by their model to existing vegetation maps of the region. “Visual comparison of the predicted PNV distributions with their actual equivalents indicates a good agreement” [Liu 2009: 1317]. Some modeled vegetation types did not agree with existing maps, however, meaning that “some more important environmental factors may have been missing in the model” [Liu 2009: 1318]. The authors also state that calibrating their model with additional field data on what is currently growing, collected from throughout the region, would improve the model’s accuracy.
The article concludes by stating that a ‘vegetation-environment’ model can help to determine PNV under not only current, but also predicted future environmental conditions.
A vegetation type, or plant community, is identifiable by its distinct appearance compared to other landscape types within a landscape. For example, a grassland and a wetland differ in appearance from each other and from a forest, while a wetland-forest is yet another visibly different vegetation type. Plant species are recognizable by their form, which is related to how the plant functions. For example, in dry environments, plant leaves are more compact with harder surfaces to limit water loss, while plants in wetter environments have larger, “softer” leaves that release water readily when pores open to take in CO2. Such leaves have more surface area for photosynthesis, resulting in faster growth.
This form-function relationship explains why vegetation types differ around the globe. Plant species are adapted to particular climatic conditions according to their proximity to the equator or a coastline, for example, or their elevation.
The geographic regularity of vegetation distribution arises, of course, from the geographic regularity of Earth’s main climatic regions, driven by the global circulation pattern of the Earth’s atmosphere [Box 2013: 466].
These authors define the concept of a plant community through discussion of its evolution. They start by defining the term ‘vegetation’ in a way that may surprise some readers because it excludes plants growing in certain situations. To be considered vegetation, plants need to emerge spontaneously.
Vegetation, the central object of study in vegetation ecology, can be loosely defined as a system of largely spontaneously growing plants. Not all growing plants form vegetation, for instance, a sown corn field or a flower bed in a garden do not. But the weeds surrounding such plants do form vegetation. A pine plantation will become vegetation after some years of spontaneous growth of the pine trees and the subsequent development of an understory [van der Maarel 2013: 1].
Two competing schools of thought regarding the nature of a stand of plants growing together geographically are represented by two early 20th Century botanists. H.A. Gleason observed “that species are ‘individualistically’ distributed along omnipresent environmental gradients and thus cannot form bounded communities” [van der Maarel 2013: 2]. By contrast, E. Clements compared plant community with an integral organism, where the whole was greater than the sum of its parts. During the same time period, the Braun-Blanquet approach was developed, which “paid much attention to the relations of plant communities with the environment and the interactions within communities, which is now incorporated in the concept of ecosystem” [van der Maarel 2013: 2].
The authors state that while individual plant species are distributed according to abiotic environmental conditions, the fact of being co-located with particular sets of other species in a particular environment results in interspecies interactions, which are in fact ecosystem processes (emergent properties).
In conclusion, a plant community is generally recognized as a relatively uniform piece of vegetation in a uniform environment, with a recognizable floristic composition and structure, that is relatively distinct from the surrounding vegetation. Even if the populations of the participating species are usually distributed individualistically in the landscape, they may well interact within the community and build up an integrated unit with emergent properties. At the same time, plant communities can be convenient units for conveying information about vegetation and its environment [van der Maarel 2013: 4].
This article provides an overview of types of plant communities and the process of succession in those communities.
In each type of habitat, certain species group together as a community. Fossil records indicate that some of these groups (or very closely related precursors) have lived together for thousands or even millions of years. During that time, it is possible that an intricate balance has been fashioned. Community members share incoming solar radiation, soil water, and nutrients to produce a constant biomass; they recycle nutrients from the soil to living tissue and back again; and they alternate with each other in time and space. Synecologists attempt to determine what is involved in this balance between all the species of a community and their environment [Barbour 1987: 155].
Community concepts and attributes
A plant community is an identifiable stand of plants growing together in a certain spot. Clusters of species, called associations, are often found growing together in several different places within a larger region. “An association is a particular type of community, which has been described sufficiently and repeatedly in several locations such that we can conclude that it has: (a) a relatively consistent floristic composition, (b) a uniform physiognomy [appearance], and (c) a distribution that is characteristic of a particular habitat” [Barbour 1987: 156].
There are opposing views about why particular plant species are often found growing together in a plant community. Thecontinuum viewposits that species distribution is driven individualistically by each species’ particular tolerance to various environmental conditions. By contrast, the association view suggests that a plant community is an integrated whole, whose component species are interdependent.
Whatever the reasons that particular species tend to grow together in stands, however, such stands “exhibit collective or emergent attributes beyond those of the individual populations” [Barbour 1987: 159]. Examples of such community attributes include its vertical structure, canopy cover, species composition and diversity, biomass, productivity, stability, and nutrient cycling, for example.
Succession
Ecological succession is an important concept that helps explain the particular assemblage of plants growing in a given location.
“Plant succession is a directional, cumulative change in the species that occupy a given area through time” [Barbour 1987: 230]. This does not refer to cyclical changes that occur over seasons, nor to changes occurring in response to climate shifts over extremely long time spans like thousands or millions of years. Rather, succession is when the composition of plants at a particular site changes over a period of decades to centuries.
Succession begins when pioneer species colonize bare ground. These first arrivals tend to be opportunists that grow fast, reproduce quickly, and do not live long. The early successional plants start to improve the habitat conditions for other, more competitive plants to then take over, displacing the pioneers. “One of the driving forces behind succession is the effect plants may have on their habitat. Plants cast shade, add to the litter, dampen temperature oscillations, and increase the humidity, and their roots change the soil structure and chemistry. … Both the environment and the community change, and this metamorphosis is due to the activities of the organisms themselves.” [Barbour 1987: 233]
Overtime, slower-growing, larger, longer-living plant species outcompete the earlier successional species, eventually forming a climax community, which is not subsequently replaced by any other community. “Succession often leads to communities with greater and greater complexity and biomass and to habitats that are progressively more and more mesic (moist)” [Barbour 1987: 233]. Such changes result in climax communities tending to be self-sustaining due to efficient nutrient cycling and internal moderation of external fluctuations in temperature and humidity.
The particular plant composition of a climax community depends on the regional climate, as well as local soil conditions and topography, meaning that several climax communities can exist in a given landscape.
Typically, many plant communities coexist in a complex mosaic pattern. That is, one climax community does not cover an entire region. … In [some] cases, the mosaic reflects topographic differences, such as south-facing versus north-facing slopes, basins with poor drainage and fine-textured soil versus upland slopes with good drainage and coarser soil, or different distances from a stress such as salt spray. In such cases, the communities within the mosaic do not bear a successional relationship to one another; they constitute a toposequence. Each community in a toposequence may, in fact, be a climax community [Barbour 1987: 238].
Understanding ecological succession can help us to predict the future vegetation of a site by observing its current vegetation. “It is often possible to estimate a community’s future composition by extrapolation from changes measured in a short time, by comparing other communities that have plants of different ages, or by noting differences between overstory plants and understory seedlings” [Barbour 1987: 231] In some cases, the understory seedlings will later become the canopy, provided the localized conditions support this succession.
Produced by the Native Plant Society of Oregon, this article argues that, while the use of native species is an accepted tenet of conservation, the term “native” is not necessarily well understood; they attempt to clarify the term.
“Any definition of a native species, native ecosystem, or native landscape requires an historical benchmark” [Wilson 1991: 16]. Over the past 20,000 years, “vegetation in the Willamette Valley has changed dramatically with changing climate. Vegetation in a single place has probably varied from boreal parkland, to conifer forest, to oak savanna, to prairie. Each climatic phase supported a different flora” [Wilson 1991: 16]. Each of these vegetation types was native to a particular place, according to particular climatic conditions that changed overtime. The vegetation that developed in the past 10,000 years – the current Holocene period of climate stability – is thus the relevant reference.
“For the Pacific Northwest, the period that ended with Euro-American settlement is a natural historical benchmark. This period lasted long enough to have a significant impact on the vegetation of the region. The climates of much earlier times were different enough to limit their usefulness in defining today’s ecosystems” [Wilson 1991: 16]. Thus, “any species that had occurred in a particular ecological habitat [of the Pacific Northwest] before Euro-American settlement is a species native to that habitat” [Wilson 1991: 17].
A native ecosystem, then, is one dominated by native plants, animals and microorganisms that occurred together before the time of Euro-American settlement. Key species – for example, the dominant photosynthesizing plants, the top carnivores, the important decomposers, the nitrogen-fixers – must be present for a native ecosystem to persist and function on its own. To artificially maintain a conserved or restored ecosystem without all of its crucial components is both difficult and expensive. The species of native ecosystem must also occur together in nature. For example, landscaping with an artificial mixture of native species like vine maple, blue bunch wheatgrass, and Jeffrey pine does not produce a native ecosystem. These species are native to different areas within Oregon, but they would not naturally grow together in the same ecosystem. Restoration of native ecosystems must also account for proper structure and appearance. For example, a red fescue lawn does not have the structural complexity and species diversity exhibited by native bunchgrass prairies [Wilson 1991: 17].
Key species – for example, the dominant photosynthesizing plants, the top carnivores, the important decomposers, the nitrogen-fixers – must be present for a native ecosystem to persist and function on its own. To artificially maintain a conserved or restored ecosystem without all of its crucial components is both difficult and expensive [Wilson 1991: 17].
The following articles lay out a few key ecological concepts and terms that may be helpful to become familiar with for the growing number of biodiversity-conscious people and organizations that are beginning to plant more native plants on their land.
Native plants, native ecosystems, and native landscapes: an ecological definition of “native” will promote effective conservation and restoration, Wilson, Hibbs & Alverson 1991
Produced by the Native Plant Society of Oregon, this article argues that, while the use of native species is an accepted tenet of conservation, the term “native” is not necessarily well understood; they attempt to clarify the term.
“Any definition of a native species, native ecosystem, or native landscape requires an historical benchmark” [Wilson 1991: 16]. Over the past 20,000 years, “vegetation in the Willamette Valley has changed dramatically with changing climate. Vegetation in a single place has probably varied from boreal parkland, to conifer forest, to oak savanna, to prairie. Each climatic phase supported a different flora” [Wilson 1991: 16]. Each of these vegetation types was native to a particular place, according to particular climatic conditions that changed overtime. The vegetation that developed in the past 10,000 years – the current Holocene period of climate stability – is thus the relevant reference.
“For the Pacific Northwest, the period that ended with Euro-American settlement is a natural historical benchmark. This period lasted long enough to have a significant impact on the vegetation of the region. The climates of much earlier times were different enough to limit their usefulness in defining today’s ecosystems” [Wilson 1991: 16]. Thus, “any species that had occurred in a particular ecological habitat [of the Pacific Northwest] before Euro-American settlement is a species native to that habitat” [Wilson 1991: 17].
A native ecosystem, then, is one dominated by native plants, animals and microorganisms that occurred together before the time of Euro-American settlement. Key species – for example, the dominant photosynthesizing plants, the top carnivores, the important decomposers, the nitrogen-fixers – must be present for a native ecosystem to persist and function on its own. To artificially maintain a conserved or restored ecosystem without all of its crucial components is both difficult and expensive. The species of native ecosystem must also occur together in nature. For example, landscaping with an artificial mixture of native species like vine maple, blue bunch wheatgrass, and Jeffrey pine does not produce a native ecosystem. These species are native to different areas within Oregon, but they would not naturally grow together in the same ecosystem. Restoration of native ecosystems must also account for proper structure and appearance. For example, a red fescue lawn does not have the structural complexity and species diversity exhibited by native bunchgrass prairies [Wilson 1991: 17].
Key species – for example, the dominant photosynthesizing plants, the top carnivores, the important decomposers, the nitrogen-fixers – must be present for a native ecosystem to persist and function on its own. To artificially maintain a conserved or restored ecosystem without all of its crucial components is both difficult and expensive [Wilson 1991: 17].
The community as an ecological unit, Barbour, Burk & Pitts 1987
This article provides an overview of types of plant communities and the process of succession in those communities.
In each type of habitat, certain species group together as a community. Fossil records indicate that some of these groups (or very closely related precursors) have lived together for thousands or even millions of years. During that time, it is possible that an intricate balance has been fashioned. Community members share incoming solar radiation, soil water, and nutrients to produce a constant biomass; they recycle nutrients from the soil to living tissue and back again; and they alternate with each other in time and space. Synecologists attempt to determine what is involved in this balance between all the species of a community and their environment [Barbour 1987: 155].
Community concepts and attributes
A plant community is an identifiable stand of plants growing together in a certain spot. Clusters of species, called associations, are often found growing together in several different places within a larger region. “An association is a particular type of community, which has been described sufficiently and repeatedly in several locations such that we can conclude that it has: (a) a relatively consistent floristic composition, (b) a uniform physiognomy [appearance], and (c) a distribution that is characteristic of a particular habitat” [Barbour 1987: 156].
There are opposing views about why particular plant species are often found growing together in a plant community. Thecontinuum viewposits that species distribution is driven individualistically by each species’ particular tolerance to various environmental conditions. By contrast, the association view suggests that a plant community is an integrated whole, whose component species are interdependent.
Whatever the reasons that particular species tend to grow together in stands, however, such stands “exhibit collective or emergent attributes beyond those of the individual populations” [Barbour 1987: 159]. Examples of such community attributes include its vertical structure, canopy cover, species composition and diversity, biomass, productivity, stability, and nutrient cycling, for example.
Succession
Ecological succession is an important concept that helps explain the particular assemblage of plants growing in a given location.
“Plant succession is a directional, cumulative change in the species that occupy a given area through time” [Barbour 1987: 230]. This does not refer to cyclical changes that occur over seasons, nor to changes occurring in response to climate shifts over extremely long time spans like thousands or millions of years. Rather, succession is when the composition of plants at a particular site changes over a period of decades to centuries.
Succession begins when pioneer species colonize bare ground. These first arrivals tend to be opportunists that grow fast, reproduce quickly, and do not live long. The early successional plants start to improve the habitat conditions for other, more competitive plants to then take over, displacing the pioneers. “One of the driving forces behind succession is the effect plants may have on their habitat. Plants cast shade, add to the litter, dampen temperature oscillations, and increase the humidity, and their roots change the soil structure and chemistry. … Both the environment and the community change, and this metamorphosis is due to the activities of the organisms themselves.” [Barbour 1987: 233]
Overtime, slower-growing, larger, longer-living plant species outcompete the earlier successional species, eventually forming a climax community, which is not subsequently replaced by any other community. “Succession often leads to communities with greater and greater complexity and biomass and to habitats that are progressively more and more mesic (moist)” [Barbour 1987: 233]. Such changes result in climax communities tending to be self-sustaining due to efficient nutrient cycling and internal moderation of external fluctuations in temperature and humidity.
The particular plant composition of a climax community depends on the regional climate, as well as local soil conditions and topography, meaning that several climax communities can exist in a given landscape.
Typically, many plant communities coexist in a complex mosaic pattern. That is, one climax community does not cover an entire region. … In [some] cases, the mosaic reflects topographic differences, such as south-facing versus north-facing slopes, basins with poor drainage and fine-textured soil versus upland slopes with good drainage and coarser soil, or different distances from a stress such as salt spray. In such cases, the communities within the mosaic do not bear a successional relationship to one another; they constitute a toposequence. Each community in a toposequence may, in fact, be a climax community [Barbour 1987: 238].
Understanding ecological succession can help us to predict the future vegetation of a site by observing its current vegetation. “It is often possible to estimate a community’s future composition by extrapolation from changes measured in a short time, by comparing other communities that have plants of different ages, or by noting differences between overstory plants and understory seedlings” [Barbour 1987: 231] In some cases, the understory seedlings will later become the canopy, provided the localized conditions support this succession.
Vegetation Ecology: Historical Notes and Outline, van der Maarel & Franklin 2013
These authors define the concept of a plant community through discussion of its evolution. They start by defining the term ‘vegetation’ in a way that may surprise some readers because it excludes plants growing in certain situations. To be considered vegetation, plants need to emerge spontaneously.
Vegetation, the central object of study in vegetation ecology, can be loosely defined as a system of largely spontaneously growing plants. Not all growing plants form vegetation, for instance, a sown corn field or a flower bed in a garden do not. But the weeds surrounding such plants do form vegetation. A pine plantation will become vegetation after some years of spontaneous growth of the pine trees and the subsequent development of an understory [van der Maarel 2013: 1].
Two competing schools of thought regarding the nature of a stand of plants growing together geographically are represented by two early 20th Century botanists. H.A. Gleason observed “that species are ‘individualistically’ distributed along omnipresent environmental gradients and thus cannot form bounded communities” [van der Maarel 2013: 2]. By contrast, E. Clements compared plant community with an integral organism, where the whole was greater than the sum of its parts. During the same time period, the Braun-Blanquet approach was developed, which “paid much attention to the relations of plant communities with the environment and the interactions within communities, which is now incorporated in the concept of ecosystem” [van der Maarel 2013: 2].
The authors state that while individual plant species are distributed according to abiotic environmental conditions, the fact of being co-located with particular sets of other species in a particular environment results in interspecies interactions, which are in fact ecosystem processes (emergent properties).
In conclusion, a plant community is generally recognized as a relatively uniform piece of vegetation in a uniform environment, with a recognizable floristic composition and structure, that is relatively distinct from the surrounding vegetation. Even if the populations of the participating species are usually distributed individualistically in the landscape, they may well interact within the community and build up an integrated unit with emergent properties. At the same time, plant communities can be convenient units for conveying information about vegetation and its environment [van der Maarel 2013: 4].
Vegetation types and their broad-scale distribution, Box & Fujiwara 2013
A vegetation type, or plant community, is identifiable by its distinct appearance compared to other landscape types within a landscape. For example, a grassland and a wetland differ in appearance from each other and from a forest, while a wetland-forest is yet another visibly different vegetation type. Plant species are recognizable by their form, which is related to how the plant functions. For example, in dry environments, plant leaves are more compact with harder surfaces to limit water loss, while plants in wetter environments have larger, “softer” leaves that release water readily when pores open to take in CO2. Such leaves have more surface area for photosynthesis, resulting in faster growth.
This form-function relationship explains why vegetation types differ around the globe. Plant species are adapted to particular climatic conditions according to their proximity to the equator or a coastline, for example, or their elevation.
The geographic regularity of vegetation distribution arises, of course, from the geographic regularity of Earth’s main climatic regions, driven by the global circulation pattern of the Earth’s atmosphere [Box 2013: 466].
Predictive modeling of the potential natural vegetation pattern in northeastern China, Liu et al. 2009
This study uses the concept of Potential Natural Vegetation (PNV), developed in the mid-1900s by German botanist Reinhold Tüxen. Described by the authors as “one of the most successful novelties in vegetation science over the last decades” [Liu 2009: 1313], PNV can be defined as a projection of the natural vegetation that would exist in a given area in the absence of human interference.
“By showing the relationships between environmental variables and vegetation types, maps of the PNV are an important instrument in the study and planning of the environment, and act as decision-support tools for the solutions to environmental issues” [Liu 2009: 1313]. Such maps are informed by studying remnant natural (old growth) vegetation in the area and site observations of the area to be mapped.
Computer modeling can be used to predict “the geographic distribution of vegetation composition across a landscape from mapped environmental variables, such as climate, soils, and geology. When a predictive vegetation modeling is calibrated using observation of vegetation composition taken from mature or ‘climax’ vegetation stands, then potential natural vegetation is portrayed in a predictive map” [Liu 209: 1314].
Focusing on northeastern China, the study identified 16 vegetation types in the region, along with the environmental factors influencing their distribution. Climatic factors included: mean annual temperature, mean temperature of the coldest month, relative humidity, and potential evapotranspiration rate. Topographical factors were elevation and slope.
“Generally, as the elevation increases, the change of temperature and moisture leads to the obvious differentiation phenomenon in vegetation vertical zones. Slope is related to the hydrology (overland and subsurface flow velocity and runoff rate) and potential soil moisture and soil development of a habitat” [Liu 2009: 1315].
They compared the map created by their model to existing vegetation maps of the region. “Visual comparison of the predicted PNV distributions with their actual equivalents indicates a good agreement” [Liu 2009: 1317]. Some modeled vegetation types did not agree with existing maps, however, meaning that “some more important environmental factors may have been missing in the model” [Liu 2009: 1318]. The authors also state that calibrating their model with additional field data on what is currently growing, collected from throughout the region, would improve the model’s accuracy.
The article concludes by stating that a ‘vegetation-environment’ model can help to determine PNV under not only current, but also predicted future environmental conditions.
Conceptualizing communities as natural entities: a philosophical argument with basic and applied implications, Steen et al. 2017
Ecological restoration aims to recreate lost or degraded ecological communities. However, “community” has been a difficult concept to define – should the definition stress dominant species, species interactions, or a subset of strongly interacting species? These authors propose defining community on the basis of co-evolutionary relationships among species.
We propose that an Evolutionary Community is conceptualized as a unique grouping of species, which occur in a given geographic area and are connected by interspecific and abiotic interactions that have evolved over time [Steen 2017: 1021].
By treating communities “as entities that have formed over evolutionary time; this [Evolutionary Community] concept allows for a philosophical platform to help us understand what many conservation and restoration efforts are trying to accomplish” [Steen 2017: 1031]. That is, it offers a way to conceptualize the end goal of a restoration project.A particular evolutionary community could be recreated by assembling the constituent species, resulting in the ecological interactions among the species resuming as before.
What processes cause a group of species to cohere into a community? We argue that the parts of Evolutionary Communities are bound together by interspecific interactions in a shared biotic and abiotic environment, which promote co-evolution and community structure and dynamics. For example, longleaf pine trees are conduits for lightning strikes that ignite a highly flammable understory, often including dropped longleaf pine needles. The resulting ground fires are necessary for reproduction of other species and maintain habitat suitable for others (e.g., gopher tortoises). Gopher tortoises, through the process of burrow creation, provide structure important to other species. The establishment of one or more of the species listed above facilitated the persistence of additional species [Steen 2017: 1025].
Likewise, the demise of one species will negatively affect, or even cause the demise of, other species that depend on it. Thus, the reason to preserve or recreate an integral community is to support the interdependent component species, each of which in turn support the community as a whole.
Bridging ecology and conservation: from ecological networks to ecosystem function, Harvey et al. 2017
This article emphasizes the importance of species interactions as drivers of ecosystem function.
The classic conservation approach is to set aside national parks or to target specific species for protection, based on their rarity or endangered status. However, these approaches can have trade-offs for non-target species, while also potentially failing to protect ecosystem function. The authors, therefore, suggest that species interactions based on their functional significance should be the main focus on conservation efforts.
We propose that a shift in focus from species to interaction networks is necessary to achieve pressing conservation management and restoration ecology goals of conserving biodiversity, ecosystem processes and ultimately landscape-scale delivery of ecosystem services [Harvey 2017: 371].
Species depend on many other species in their communities, either directly or indirectly. An example of indirect dependence is the Phengaris arion butterfly’s need for European rabbits. The butterfly uses ant nests made in the open areas supplied by rabbit grazing for development of its larvae. Thus, no rabbits means no ants, which means no Phengaris arion.
Focusing on species interactions is more meaningful even than measuring species richness (the number of different species), because interactions can disappear – even if both species are present – if either group’s abundance has significantly dropped. The authors offer the example of 59 regionally extinct lepidoptera (butterfly and moth) species of central Europe. Eight of these extinctions were associated with the loss of particular host plant species, which actually occurred after the lepidoptera went extinct.
Focusing on species interactions is more meaningful even than measuring species richness (the number of different species), because interactions can disappear – even if both species are present – if either group’s abundance has significantly dropped.
Thus, strong declines of host plants can have cascading extinction effects on higher trophic levels before the plants actually go extinct, illustrating that interactions can be lost before any actual decline in species richness (plants persisted at low abundance). This illustrates that preserving keystone interactions, rather than species, can be a proactive way to maintain ecosystem integrity in the face of global change instead of allocating resources to already endangered species [Harvey 2017: 372].
There is interdependence among species even between neighboring ecosystems. For example, a manta ray species in the Palmyra Atoll south of Hawaii depends on two species of native trees to maintain its ocean plankton diet. When these trees were replaced with cultivated coconut palms, marine-foraging birds no longer nested on that shore, depriving the coastal waters of the nitrogen runoff from their guano, which had been feeding the plankton population.
The authors recommend that “the main lever to restore or conserve ecological network structure and stability is the management of spatial configuration” [Harvey 2017: 377]. Reflecting on the Palmyra Atoll, for example, it’s clear that a marine conservation plan would be incomplete without considering the nutrient flow from the tree-bird interactions on land.
Interactions among plants and evolution, Thorpe et al. 2011
This review explores the question of whether plant-plant interactions drive evolutionary changes. “If such evolution is common, plant communities are not random assemblages of species.” The topic is under-studied compared to plant interactions with other groups.
Research on plant–consumer, plant–pollinator and plant–disperser interactions has been central to understanding the complex mutualistic and co-dependent interactions among species that structure communities. However, with some notable exceptions, interactions among plants have not been emphasized as processes that contribute to selection and evolution [Thorpe 2011: 730].
“The simplest interactions among plants are direct interactions, such as facilitation, resource competition and allelopathy” [Thorpe 2011: 731]. Facilitation is when one plant protects an adjacent plant, such as from drought and heat by providing shade, for example, or from browsing by being thorny or toxic to herbivores and surrounding the facilitated plant. Allelopathy refers to plants’ release of toxic substances that suppress the growth of another organism, including other plants. In natural communities, any given plant may be interacting with several different plants at the same time.
In natural communities, any given plant may be interacting with several different plants at the same time.
Competition for sunlight, water, and nutrients drives niche differentiation, or the carving out by species of particular spaces or timing within an ecosystem to obtain a share of limited resources. “The exceptionally rich body of ecological literature on the niche is based in part on the idea that competition can drive the evolution of niche differentiation, thus allowing species to coexist” [Thorpe 2011: 732].
Thorpe et al. refer to an example from a 1976 article by Parrish & Bazzaz , who “found that resource partitioning, as estimated from spatial overlap among root systems, was higher in stable prairie communities with a long community history than in early successional old-field communities composed of species without a common history” [Thorpe 2011: 731]. In other words, plants with a long coexistence history more efficiently divvy up resources than do species lacking a common community history.
The primary hypothesis for positive diversity–ecosystem function relationships has been niche ‘complementarity’, the idea that different species or functional groups occupy niches different enough from each other to more fully utilize resources or space, increasing and stabilizing productivity, and making it more difficult for other species to enter the community [Thorpe 2011: 733].
The authors are somewhat inconclusive, however, about what drives niche complementarity (resource partitioning).
We do not yet know whether complementarity is produced by interactions causing evolutionary shifts in niche space (and thus coexistence and more complete resource use) or by sorting of the existing species pool [Thorpe 2011: 733].
Plants can also adapt to one another’s allelopathic substances over time, a fact that contributes to the argument that plant-plant interactions produce evolutionary changes. “Recent experiments raise the possibility that some invaders may exude allelochemicals that are relatively ineffective against neighbors in natural communities, but highly inhibitory to plants in invaded communities” [Thorpe 2011: 734].
Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities, Burghardt et al. 2010
This research evaluates the impact of the invasion of non-native plants in the distribution of lepidopteran (butterfly, skipper, and moth) communities. The authors assert that although the introduction of non-native plants has not resulted in a “global extinction”, they have had a considerable impact on how ecosystems function—they often result in significant bottom-up reductions of energy available in local food webs.
The experiment established four gardens near mature woodlots containing most, if not all, of the native species planted within the treatment. The richness and abundance were then compared for lepidopteran communities found on native versus corresponding non-native congener[4] species of 13 woody plant genera. For example, the genus Acer (maple) was selected for this study because the native and non-native maples were widespread in that area. In separate plots, the researchers also compared native plants and unrelated (non-congeneric) non-native plants for lepidopteran richness and abundance.
The study found that lepidopterans suffer from the replacement of native plants by non-natives, especially when those non-natives are unrelated to any native plant species. The authors explain that “insect herbivores adapted to the chemical challenges [toxic plant defenses] of particular native hosts may be able to adopt a novel plant species as a host if its phytochemistry is sufficiently similar to the original hosts” [Burghardt 2010: 10]. Over the two-year study, lepidopteran abundance and richness were depressed both on congener and (unrelated) non-congener non-native plants, but especially on the latter.
The study found that lepidopterans suffer from the replacement of native plants by non-natives, especially when those non-natives are unrelated to any native plant species.
Specialist lepidopteran species, which require specific diet and habitat conditions to survive, fared worse on non-natives than did generalists, which can eat a variety of foods and survive in many different habitats. The authors note, for example, that “geographically novel congeners were acceptable hosts to less than half of the generalists and only one fourth of the specialists that we found on native congeners in 2009” [Burghardt 2010: 11]. Only 7% of specialist species used non-congener non-natives as hosts.
The authors argue that the loss of lepidopteran diversity and abundance due to the displacement of native plant species with non-natives can ripple up the food chain, reducing diversity at higher trophic levels. Reduced diversity leads to lower ecosystem productivity and stability, thus disrupting the whole system.
The authors argue that the loss of lepidopteran diversity and abundance due to the displacement of native plant species with non-natives can ripple up the food chain, reducing diversity at higher trophic levels. Reduced diversity leads to lower ecosystem productivity and stability, thus disrupting the whole system.
Because insect herbivores are near the hub of most terrestrial food webs, comprising essential food stuffs for an incredible diversity of insect predators and parasitoids, spiders, amphibians, lizards, rodents, bats, birds, and even higher predators such as foxes and bears, it is particularly important to understand changes wrought by non-native plants on this critical taxon [Burghardt 2010: 13].
Impact of Native Plants on Bird and Butterfly Biodiversity in Suburban Landscapes, Burghardt, Tallamy & Shriver 2008
In this study, the insect and bird populations of six pairs of suburban yards were measured. Each pair contained one conventionally landscaped yard containing native canopy trees and a mixture of native and non-native shrubs, grasses and understory trees; and one yard with native species only (canopy, understory, shrub and grasses). The level of plant diversity was comparable between each of the pair; only the proportion of native species differed. The authors found that:
Avian abundance, diversity, richness, and biomass (particularly bird species of conservation concern) were all greater on native properties. Native nesting birds that are mostly dependent on insect populations to feed their young were more abundant on native properties. Lepidoptera [butterfly and moth species] abundance and diversity were also higher on native properties, suggesting that food availability might account for the differences detected in the bird communities between native and conventionally landscaped sites [Burghardt 2008: 223].
These results support the authors’ hypothesis based on an understanding of the co-evolutionary roots of species interactions.
Theory backed by decades of empirical evidence predicts that up to 90% of all species of insect herbivores can successfully reproduce only on plant lineages with which they have shared an evolutionary history [Burghardt 2008: 220].
Native plants improve breeding and foraging habitat for an insectivorous bird, Narango, Tallamy & Marra 2017
This study examined whether non-native plants in residential Washington DC limited the presence of the Carolina chickadee, a local breeding insectivore.
We predicted that areas with more native plants would support more chickadees, and chickadees would forage more often in the most insect-producing native plants [Narango 2017: 43].
The authors had also considered the possibility that non-native plants could promote increases in other food items (e.g. non-native arthropods), keeping overall prey biomass similar between native and non-native plants. What they found, though, affirmed their prediction: native plants produce more caterpillars, which in turn support more chickadees. In fact, the birds avoided foraging in non-native plants, including non-native species of the same tree genera: the chickadees preferred maples native to the eastern US compared to European-origin maples.
Native plants produce more caterpillars, which in turn support more chickadees.
Native plants were more likely to host a higher biomass of caterpillars compared to non-native plants, and chickadees strongly preferred to forage in native plants that supported the most caterpillars. In addition, chickadees were less likely to breed in yards as the dominance of non-native plants increased [Narango 2017: 42].
Also unique to our study is that we measured the probability of caterpillar occurrence between congeneric species (e.g. native vs. non-native Acer [maple]). This is particularly important considering the popularity and invasive qualities of congeneric species in this region such as Acer platanoides and Quercus acutissima. Although non-native congeners support more caterpillars in comparison to plants unrelated to any native species, congeners had a 47% (CI: 34%–59%) lower probability of having caterpillars compared to native species [Narango 2017: 47].
The authors state that local insects are adapted to local plants, presumably due to their shared co-evolutionary history.
This occurs in part because herbivorous insects have adapted to circumvent the phytochemical defenses of particular plant lineages, resulting in a radiation of specialized plant-insect associations. During urban conversion, native plants are replaced by non-native species with novel chemical, physical, and phenological features for which native herbivorous arthropods have few physiological or behavioral adaptations [Narango 2017: 42].
Do non-native plants contribute to insect declines? Tallamy, Narango & Mitchell 2020
The widespread distribution of plants outside of their native range due to human activity is a significant yet underrecognized cause of global insect decline, according to this article. To illuminate the issue, the authors: “examine the evidence for and against the hypothesis that long term changes in the species composition of plant assemblages have contributed to local and global declines in the abundance and diversity of the insect communities dependent upon those assemblages” [Tallamy 2020: 2].
To be sure, insect conservationists have long noted the importance of habitat containing appropriate native host plants, but the widespread replacement of native host plants with non-native species has yet to penetrate the growing literature on insect declines in any meaningful way [Tallamy 2020: 1].
It is not simply the absence of native plants harms plant-eating insects, however, but also the presence of non-natives. While some insects feed successfully on non-native plants, this is the minority. Most either avoid non-native plants, or do use them and are killed or malnourished by doing so. For example,
Swallowworts (Vincetoxicum spp.) are confamilials of milkweeds (Asclepias spp.) and have become invasive in parts of the northeastern United States. Similar phytochemistry between swallowworts and milkweeds can lead monarch butterflies (Danaus plexxipus) and milkweed beetles (Chrysochus auratus) to fatally mistake these chemically protected plants as hosts. The degree to which Vincetoxicum act as ecological traps for these taxa is likely to become more pronounced as the plants become dominant and displace milkweeds in the landscape [Tallamy 2020: 3].
Species that share a particular environment over hundreds or thousands of years evolve in relation to one another. For plant-eating insects, adapting to certain plants meant developing “traits to detect and tolerate plant defenses over time” [Tallamy 2020: 2]. Most herbivorous insects adapted to only a particular set of plants, specializing in feeding on those plant hosts.
The diet of most insects is constrained to a single plant family in any one habitat or location, with dietary specialization even narrower both in many temperate lineages and hyper-diverse tropical lineages. In fact, diet specialization increases with decreasing latitudes, concurrent with theories of increased plant and animal diversity in the tropics [Tallamy 2020: 2].
When native plants are displaced in the landscape by non-native species, phytophagous [plant-eating] insects typically do not recognize the novel host for feeding or oviposition [egg laying], or may be unable to overcome novel plant defenses. The concurrent loss of native plant hosts and dominance of non-native plants can lead to local extirpation of phytophagous insects and thus to changes in the composition and structure of local food webs [Tallamy 2020: 2].
The most likely successful substitute for a native plant is a non-native plant in the same genus or family.
Non-native congeners [members of the same genus] or confamilials [members of the same family] that are similar in foliar chemistry and nutrition, phenology, and morphology, may occasionally serve as novel hosts for herbivorous insects and support higher diversity and abundance than non-native, non-congeners. However, novel use of congeners may increase larval mortality, extend development or pupation time, reduce biomass, and reduce fitness compared to that of native hosts [Tallamy 2020: 3].
The narrower the native plant diet an insect species has, the less likely to tolerate novel, non-native food sources. However, there are more species of specialist insects than of generalists, meaning a larger proportion of susceptible species. Adaptability to exotic host plants also depends on an insects’ feeding habits.
Insects with chewing (mandibulate) mouthparts are typically more susceptible to defensive secondary metabolites contained in leaf vacuoles than are insects with sucking (haustelate) mouthparts that tap into poorly defended xylem or phloem fluids. Thus, sucking insects find novel non-native plants to be acceptable hosts more often than do chewing species [Tallamy 2020: 4].
Considering that there are more than 4.5 times as many mandibulate insect herbivores as haustelate species, there is reason for concern when non-native plants replace native hosts; the largest guild of insect herbivores is also the most vulnerable to non-native plants and the most valuable to insectivores [Tallamy 2020: 5].
“The dispersal and spread of invasive plants has been driven by global trade networks and colonialism” [Tallamy 2020: 6] and, more specifically, from agroforestry, forestry, agriculture, and horticulture.
Although plants have always distributed themselves around the globe, the increased temporal and spatial mobility of humans has resulted in an extraordinary increase in the rate of plant movements and most species’ introductions have happened in the last 200 years. Habitat is rapidly being converted from coevolved native ecosystems into novel assemblages of plants and animals, making the conversion of native plant communities into plant assemblages dominated by non-native species one of the most ubiquitous threats to biodiversity today. The introduction of non-native plants has completely transformed the composition of present-day plant communities in both natural and human-dominated ecosystems around the globe and the magnitude of introductions is staggering. An estimated 13,168 plant species (about 3.9% of global vascular flora) have been introduced and naturalized beyond their native ranges as a result of human activity [Tallamy 2020: 6].
Global exchange and accumulation of non-native species, van Kleunen et al. 2015
The ecological, economic, and social damage of human-mediated dispersal of species into new regions, where they possess the ability to naturalize (become self-sustaining their new homeland), is one of the defining features of the Anthropocene Epoch. Globally, human activity has led to the naturalization of nearly 13,168 plant species (equal in size to the native European flora). The results from this research provide a baseline for monitoring global changes in biodiversity while highlighting the immediate action that has to be taken to comprehend and determine the spread of alien species on an international scale.
The ecological, economic, and social damage of human-mediated dispersal of species into new regions, where they possess the ability to naturalize (become self-sustaining their new homeland), is one of the defining features of the Anthropocene Epoch.
At least 3.9% of all currently known vascular plant species have become naturalized outside their natural ranges as a result of human activity. With the continued practice of international traffic and trade and globalization, the likelihood of more and more species being introduced and getting naturalized outside their native range is high.
To assess the accumulation of naturalized species in each continent as well as which continents have been the major donors of alien naturalized plant species globally, the researchers used a novel database, Global Naturalized Alien Flora (GloNAF), in addition to the data on the origin of naturalized species and estimates of the number of native species per continent. They found that when not taking into account the differences in total area, North America has accumulated the highest number of naturalized species (n=5,958). However, when considering the difference in total area, Australasia (a region comprising Australia, New Zealand, and neighboring islands) was found to have more extra-continental species than North America.
One possible explanation is that Australia’s long biogeographical isolation and drying climate have resulted in a native flora that is phylogenetically distinct, but not well-adapted to exploit the novel habitats created by European settlers [van Kleunen 2015: 101].
The major donors of alien species are Europe and temperate Asia, while North America is also a significant donor.
Ecological and evolutionary consequences of biotic homogenization, Olden et al. 2004
Anthropogenic environmental change and global dispersal of a wide variety of species outside their native ranges has expanded the range of “cosmopolitan,” non-native species and shrunk the range of regional and endemic species. “This replacement of specific native forms by generalist non-natives in space and time has mixed the taxonomic composition of once disparate biotas, an occurrence termed ‘biotic homogenization’” [Olden 2004: 18].
The authors explore the effect of this “global erosion of regional distinctiveness” [Olden 2004: 18] at three levels: Genetic homogenization reduces genetic variability within species or among populations of species, while taxonomic homogenization reduces distinctiveness among communities. Functional homogenization refers to a reduction of functional traits within an ecosystem. The identity of species making up a community, along with their respective functional traits, determines “ecosystem functions (such as nutrient retention or energy flow)” [Olden 2004: 20], so that narrowing species compositions risks diminishing ecosystem function.
A decrease in functional diversity might reduce overall community and ecosystem functioning, stability and resistance to environmental change by simply narrowing the available range of species-specific responses. Consider a severe drought that strongly affects a subset of species in a community that has (or lacks) a particular suite of functional traits. Historical communities, with much greater breadth in functional space, should exhibit higher resistance or resilience when compared with homogenized communities [Olden 2004: 20].
Genetic homogenization occurs when two distinct locally adapted populations of the same species interbreed. It also occurs when a single variety (such as captive fish bred in a central location) are released in many places to replenish dwindling native stocks. While such mixing has the potential to increase species diversity, this outcome is not assured.
Intraspecific hybridization can homogenize the unique characteristics of geographically distinct populations, as well as compromise the fitness of individuals by disrupting local adaptations [Olden 2004: 19].
This book draws lessons from ecological succession theory to inform ecological restoration, stating that: “restoration is fundamentally the management of succession” [Walker 2007: vi]. The latter is the natural process by which plants first colonize “new” land (post landslide, glacial retreat or volcanic eruption, for example) or degraded land, and over time develop into mature ecosystems through a series of changing plant communities. Ecological restoration is a human-led initiative to restore functioning ecosystems, or at least vegetation, on land degraded through human activity. The ultimate goal of restoration is to “establish a self-sufficient ecosystem that requires minimal or no continuing human inputs in order to provide a continuing supply of goods and services” [Hobbs 2007: 177].
Effective ecosystem restoration requires ecological knowledge. Likewise, the outcomes of such projects demonstrate our comprehension, or lack thereof, of ecological concepts: “Restoration is the acid test of our ability to understand not only how ecosystems are assembled and held together but also how they change over time” [Walker 2007: vi]. The authors contend, however, that restoration projects are more often guided by engineering, horticulture, and agronomy than by ecology. Aiming to clarify the ways in which ecological succession theory can and should inform restoration, this book poses the question: “What is the minimum amount of biophysical and successional information needed to restore a specific landscape or area” [Walker 2007b: 2]?
Succession comprises many ecological processes that underpin all ecological restoration and ecological restoration is a manipulation of these processes to achieve its goals. This means it is essential to understand how succession operates, and when and how to manipulate it [Prach 2007: 121].
Restoration can explicitly embrace a hands-off approach, where land is simply left to repair itself through natural ecological succession. On the other hand, understanding the successional process allows manipulation of various stages to speed up the process. For example, in the first stage of primary succession “winds deposit dust, pollen, seeds, and insects crucial to reducing infertility” [del Moral 2007: 23], on bare, inhospitable ground. Tough pioneer plants are able to establish then create shade, trap sediment, and deposit organic matter when they die, creating slightly better conditions for the next wave of colonizing plants. To mimic this first stage of site “amelioration”, the site can be physically manipulated by reshaping the ground for improved drainage or adding organic matter, for example.
Biological manipulation involves sowing or planting local/native varieties of later successional species that may not be otherwise present in the area due to human transformation of the broader landscape. While earlier successional species tend to have small, easily transported seeds, the later successional species (such as large canopy trees) that are often the target of restoration efforts often have large, less mobile seeds. Thus, if those plants are not present in the immediate environment as seed stock, they may never establish in the restored site without human assistance.
Near-Natural Silviculture: Sustainable Approach for Urban Re-naturalization Assessment Based on 10 Years Recovering Dynamics and Eco-Benefits in Shanghai, Guo et. al 2015
As one of China’s major cities, Shanghai’s natural sub-ecosystem[5] has suffered drastic damage due to human activities and urbanization. Although urban re-naturalization has gained attention from city leaders, urban tree planting has largely consisted of two methods with limited ecological potential. One favors fast-growing monocultures to produce timber products and other benefits, while the other approach is to plant non-native species for decorative purposes. The authors believe the restoration progress of the natural sub-ecosystem could be further improved by adopting the “near-natural” method based on the concepts of potential natural vegetation[6] and ecological succession.
The near-natural forest uses all native species and aims to create a complex structure with high biodiversity, high biomass and multiplayer canopies. It was adopted successfully in many countries, but the authors thought long-term studies of these forests were lacking. Therefore, they conducted a 10-year study at a near-natural forest established in 2000 in Pudong New Area of Shanghai to investigate the effectiveness of the forest in providing ecological benefits.
Results showed that the near-natural forest had higher sustainability value than artificial (“even-aged, managed”) forest in Shanghai based on its ecological and economic benefits. The high tree density and multiple vertical structures of the forest improved the air quality and soil fertility and decreased the concentrations of air bacteria and dust. It also had a much lower planting and maintenance cost than artificial traditional methods. Although the near-natural forest could not transcend the benefits of the natural forest, the study successfully proved its important role in urban re-naturalization by bridging the difference between the artificial and natural forests.
Results showed that the near-natural forest had higher sustainability value than artificial (“even-aged, managed”) forest in Shanghai based on its ecological and economic benefits
Over the course of the study, authors discovered a potential limitation of the approach, at least in its application in Shanghai. They observed high evergreen seedlings mortality, attributable to over-exposure to sunlight. Therefore, in subsequent plantings in 2003 and 2004, the authors modified the approach to optimize it to local conditions:
The key to the new method is to create a mixed deciduous–evergreen community by simultaneously planting shade-tolerant evergreen broad-leaved species and light-demanding deciduous broad-leaved species, but using smaller individuals for the former and larger individuals for the latter to form a multilayer vegetation structure. The shade-tolerant evergreen species benefit from the rapid growth of the light-demanding deciduous species, which offer shade and nutrients in the form of litter layer-based fertilizer, improving the soil for the evergreen species [Guo 2015: 5].
Overall, they suggest that the near-natural forest is a very sustainable method to be applied in Shanghai.
Tree planting is not a simple science, Holl & Brancalion 2020
Well-planned tree-planting projects are an important component of global efforts to improve ecological and human well-being. But tree planting becomes problematic when it is promoted as a simple, silver bullet solution and overshadows other actions that have greater potential for addressing the drivers of specific environmental problems, such as taking bold and rapid steps to reduce deforestation and greenhouse gas emissions [Holl 2020: 580].
Some of the pitfalls to avoid in tree planting initiatives, according to the authors, include:
Use of non-native species, which does not result in a true forest and can result in ground water depletion in arid environments.
Planting trees in historic grasslands and savannas, harming those native ecosystems and species.
Abandoning trees after they are planted, which can result in high mortality due to insufficient water for developing saplings, being shaded out by faster growing herbaceous plants, grazing, or being re-cleared.
Planting trees in agricultural land, which risks pushing crop production into native forest land, which is then deforested.
The authors insist that reforestation takes careful planning, stakeholder engagement, clear goal-setting, and long-term monitoring and adaptive management of planted tree stands to ensure their survival. Above all, existing mature, native forests should be preserved.
The authors insist that reforestation takes careful planning, stakeholder engagement, clear goal-setting, and long-term monitoring and adaptive management of planted tree stands to ensure their survival. Above all, existing mature, native forests should be preserved.
The first priority to increase the overall number of trees on the planet must be to reduce the current rapid rate of forest clearing and degradation in many areas of the world. The immediate response of the G7 nations to the 2019 Amazon fires was to offer funding to reforest these areas, rather than to address the core issues of enforcing laws, protecting lands of indigenous people, and providing incentives to landowners to maintain forest cover. The simplistic assumption that tree planting can immediately compensate for clearing intact forest is not uncommon. Nonetheless, a large body of literature shows that even the best-planned restoration projects rarely fully recover the biodiversity of intact forest, owing to a lack of sources of forest-dependent flora and fauna in deforested landscapes, as well as degraded abiotic conditions resulting from anthropogenic activities [Holl 2020: 581].
Bio4Climate has been studying the Miyawaki Method of reforestation over the past several months. This 50-year-old technique involves densely planting native forest species from shrub to canopy layer to create tiny, fast-growing urban ecosystems[3]. Members of our staff have joined local efforts to establish Miyawaki “mini-forests” in Cambridge, MA, in Los Angeles, CA, and one in France.
The attractiveness of the Miyawaki Method should be understood in the context of a growing global zest for tree planting, such as the World Economic Forum’s 1 Trillion Trees campaign and the Bonn Initiative. In fact, tree planting initiatives can have various goals – including timber production and carbon capture, both of which favor fast-growing, often non-native trees, or erosion control or improving ground permeability. When ecosystem restoration is the explicit goal, however, reforestation aims to reestablish the full complement of ecosystem functions and wildlife habitat inherent in native ecosystems.
Not only the Miyawaki Method, but many eco-restoration initiatives favor the use of native plants over exotic species. Indigenous plants are preferred because they are typically well-adapted to local environmental conditions. Additionally, native plants interact in important ways with other local species, such as by serving as food sources for herbivores, and thus transferring energy up the food chain. Many insects, for example, are specialists and can use only certain species in their diets. When these particular plant species are replaced by non-natives or otherwise lost locally, the insects that depend on them also disappear.
Several studies by Tallamy and his colleagues [Burghardt 2008, Burghardt 2010, Narango 2017, Tallamy 2020] working in eastern US illustrate the critical role of native plants for local moths and butterflies (also called “lepidopterans”), and thus also for the birds that depend on these insects. In 2010, Burghardt, et al. reported on a two-year experiment showing that lepidopteran richness (number of species) and abundance were lower on non-native compared to native plants.
The authors explain that insect herbivores are adapted to the self-defensive toxins produced by plants for only certain plant species, which makes them especially vulnerable when those species are replaced by non-natives. This is particularly true for specialists, which make up the majority of insect herbivore species: “Most species are specialists that use only a few of the myriad plant lineages available to them for growth and reproduction. … Specialists have followed an evolutionary path that has enabled them to circumvent particular plant defenses by developing highly specific behavioral and physiological adaptations that defuse such defenses” [Burghardt 2010: 2]. The authors add, however, that “even the most generalized insect herbivores use only a small fraction of the plants in their environments” [Burghardt 2010: 2].
As expected, specialist lepidopteran species fared worse on non-natives than did the generalists, which can eat a greater variety of foods and survive in wider habitat ranges, although both groups suffered. Non-native plants (Norway maple, for example) that were similar to native ones (red maple) were more commonly used by lepidopterans, than were non-natives with no related native counterpart. The authors explain that “insect herbivores adapted to the chemical challenges [toxic plant defenses] of particular native hosts may be able to adopt a novel plant species as a host if its phytochemistry is sufficiently similar to the original hosts” [Burghardt 2010: 10]. Only 7% of specialist species used non-natives with no similar native counterpart as hosts, while less than half of generalists did.
A Washington DC study [Narango 2017] showed that both lepidoptera and their bird predators prospered in the presence of native plants:
Native plants were more likely to host a higher biomass of caterpillars compared to non-native plants, and chickadees strongly preferred to forage in native plants that supported the most caterpillars. In addition, chickadees were less likely to breed in yards as the dominance of non-native plants increased [Narango 2017: 42].
A related study [Burghardt 2008] showed greater diversity and abundance of both lepidoptera and birds (including birds dependent on insects to feed their young) in suburban yards with native plants only, compared to yards with a mix of native and non-native plants. Interestingly, all yards in the study had similar levels of overall plant diversity, suggesting that it is native biodiversity rather than biodiversity per se that counts, at least for butterflies, moths and birds.
These studies illustrate the dependence of herbivorous insects on particular co-evolved plants, and how the food web is affected when that relationship is interrupted. A related line of inquiry is whether plants themselves depend on other plants with which they share a common history. The answer to this is less clear. Thorpe et al. [2011] point outthat the effects of plant-plant interactions on evolution have received less attention from researchers:
Research on plant-consumer, plant-pollinator and plant-disperser interactions has been central to understanding the complex mutualistic and co-dependent interactions among species that structure communities. However, with some notable exceptions, interactions among plants have not been emphasized as processes that contribute to selection and evolution [Thorpe 2011: 730].
Thorpe et al. [2011] highlight studies suggesting that plants, like insects, are adapted to the toxins produced by plants with which they co-evolved. Several North American experiments show that exotic European plants strongly inhibit the growth of native plants, while not negatively affecting European members of the same genus as the inhibited native plants. “Biogeographical differences in the effects of allelopathic chemicals circumstantially suggest they have roles as drivers of evolution” [Thorpe 2011: 735], state the authors. They add, however, that invasive species exert selection pressure fairly quickly on the communities they enter, resulting in adaptation that is passed on to the next generations.
In addition, plants are able to divvy up limited resources through “niche complementarity,” which is “the idea that different species or functional groups occupy niches different enough from each other to more fully utilize resources or space, increasing and stabilizing productivity, and making it more difficult for other species to enter the community” [Thorpe 2011: 733]. The authors cite a 1976 study that “found that resource partitioning, as estimated from spatial overlap among root systems, was higher in stable prairie communities with a long community history than in early successional old-field communities composed of species without a common history” [Thorpe 2011: 733].
The question of whether plant interactions drive evolution is related to an ongoing debate about the nature of a plant community. Two competing schools of thought regarding the nature of a stand of plants growing together geographically are represented by two early 20th Century botanists. H.A. Gleason observed “that species are ‘individualistically’ distributed along omnipresent environmental gradients and thus cannot form bounded communities” [van der Maarel 2013: 2]. In this view, species distribution is driven by each species’ particular tolerance to various environmental conditions, and “that the level of interactions and interdependence is relatively low, or at least nonspecific” [Barbour 1987: 158].
Barbour et al. explain that E. Clements, by contrast, compared a plant community with an integral organism, where the whole is greater than the sum of its parts, and whose component species are interdependent in the sense that they are bound together by multiple positive (and neutral) interactions. Both camps have observed that particular plant species grow together reliably in identifiable groups, forming a vegetation type (like an oak-beech forest, for example).
Fossil records indicate that some of these groups (or very closely related precursors) have lived together for thousands or even millions of years. During that time, it is possible that an intricate balance has been fashioned. Community members share incoming solar radiation, soil water, and nutrients to produce a constant biomass; they recycle nutrients from the soil to living tissue and back again; and they alternate with each other in time and space [Barbour 1987: 155].
Do plant species grow together in definable communities simply because they share the same environmental needs or are they also somewhat interdependent members of a specific community? The jury seems still to be out on this question. Regarding the larger ecological community (including animals), Steen et al. [2017] propose the concept of an “evolutionary community,” which emphasizes interdependence among specific groups of species.
What processes cause a group of species to cohere into a community? We argue that the parts of Evolutionary Communities are bound together by interspecific interactions in a shared biotic and abiotic environment, which promote co-evolution and community structure and dynamics. For example, longleaf pine trees are conduits for lightning strikes that ignite a highly flammable understory, often including dropped longleaf pine needles. The resulting ground fires are necessary for reproduction of other species and maintain habitat suitable for others (e.g., gopher tortoises). Gopher tortoises, through the process of burrow creation, provide structure important to other species. The establishment of one or more of the species listed above facilitated the persistence of additional species [Steen 2017: 1025].
These authors add that the Evolutionary Community concept can “help us understand what many conservation and restoration efforts are trying to accomplish” [Steen 2017: 1031],
stress that interspecies interdependence is a key concept for conservation, and call for the latter to focus less on species and more on interactions among species.
We propose that a shift in focus from species to interaction networks is necessary to achieve pressing conservation management and restoration ecology goals of conserving biodiversity, ecosystem processes and ultimately landscape-scale delivery of ecosystem services [Harvey 2017: 371].
All of these studies shed some light on the role of native species in the Miyawaki Method and other eco-restoration approaches. Is it possible, though, that non-native species could replace certain native species in a way that fulfills the ecological role of the replaced natives? In some contexts, it seems that communities of non-local species can thrive. Jones [2013] cites a perhaps not-terribly representational study of the vegetation on an island stranded between Africa and Brazil. This study documents “a highly functional cloud forest on Ascension Island in the south Atlantic made up of species with no history of coevolution, having been introduced from various continents. Thus, plants do not necessarily need to have co-evolved together either locally or nonlocally to display desirable community-level interactions” [Jones 2013: 1114].
By contrast, numerous studies document local ecological decline resulting from the establishment of non-native species. Jones [2013] supports the premise that local species are generally the best choice for eco-restoration, but he notes that in the context of environmental change, it may sometimes be possible that local species are no longer the best adapted.
Globally, 13,168 plant species (3.9% of known vascular plants) have become naturalized outside their native range due to human activity [van Kleunen 2015]. The anthropogenically expanding distribution of generalist species is shrinking the range of regional, endemic, often specialist species. This process, referred to as biotic homogenization, results in a “global erosion of regional distinctiveness” [Olden 2004: 18].
At the genetic level, homogenization threatens to diminish the local adaptations of native species through mixing of populations; at the ecosystem level, functional homogenization “might reduce overall community and ecosystem functioning, stability and resistance to environmental change by simply narrowing the available range of species-specific responses” [Olden 2004: 20]. Thus, there are local, regional, and global consequences of biotic homogenization.
With respect to the role of native species in eco-restoration, the Society for Ecological Restoration [SER 2019] distinguishes between restoration and “rehabilitation.”
Rehabilitation activities are well suited to a broad range of land and water management sectors where substantial native ecosystem recovery is not possible or desirable due to competing and legitimate human needs. … rehabilitation projects achieving some improvements in ecological conditions can later be targeted for ecological restoration. For instance, where revegetation of a degraded rangeland, or post-mine site, with a mix of native and nonnative plant species and native microsymbionts has resulted in improved soil function, restoration plans can be developed that include harvesting nonnative species and replacing them with native species as well as taking other actions to assist the system to recovering to the condition it would have been in if degradation had not occurred. In some cases where soil has been stabilized with nonnative species, native species can be added (or helped to recover spontaneously) and nonnative species removed to ultimately assist the recovery of a native ecosystem [SER 2019: 52].
In summary, it seems that while plant species can potentially adapt to growing with plants from outside their native range, this can sometimes take time, limiting growth of species or communities at least in the short term. By contrast, many native herbivore species are likely to become locally extinct rather than adapt to non-native plants when the latter displace natives. The loss of insects blocks the flow of energy from plants up the food chain, negatively affecting ecosystem function. While native plants are a more perfect ecosystem puzzle piece, non-native species certainly have important roles to play – at least in ecosystem “rehabilitation” as noted above and potentially to substitute for extinct natives of the same genus or functional group. Additional novel species groupings can be expected as climate change drives some species to migrate to more suitable latitudes.
Native plants article summaries
The following articles lay out a few key ecological concepts and terms that may be helpful to become familiar with for the growing number of biodiversity-conscious people and organizations that are beginning to plant more native plants on their land.
Native plants, native ecosystems, and native landscapes: an ecological definition of “native” will promote effective conservation and restoration, Wilson, Hibbs & Alverson 1991
Produced by the Native Plant Society of Oregon, this article argues that, while the use of native species is an accepted tenet of conservation, the term “native” is not necessarily well understood; they attempt to clarify the term.
“Any definition of a native species, native ecosystem, or native landscape requires an historical benchmark” [Wilson 1991: 16]. Over the past 20,000 years, “vegetation in the Willamette Valley has changed dramatically with changing climate. Vegetation in a single place has probably varied from boreal parkland, to conifer forest, to oak savanna, to prairie. Each climatic phase supported a different flora” [Wilson 1991: 16]. Each of these vegetation types was native to a particular place, according to particular climatic conditions that changed overtime. The vegetation that developed in the past 10,000 years – the current Holocene period of climate stability – is thus the relevant reference.
“For the Pacific Northwest, the period that ended with Euro-American settlement is a natural historical benchmark. This period lasted long enough to have a significant impact on the vegetation of the region. The climates of much earlier times were different enough to limit their usefulness in defining today’s ecosystems” [Wilson 1991: 16]. Thus, “any species that had occurred in a particular ecological habitat [of the Pacific Northwest] before Euro-American settlement is a species native to that habitat” [Wilson 1991: 17].
A native ecosystem, then, is one dominated by native plants, animals and microorganisms that occurred together before the time of Euro-American settlement. Key species – for example, the dominant photosynthesizing plants, the top carnivores, the important decomposers, the nitrogen-fixers – must be present for a native ecosystem to persist and function on its own. To artificially maintain a conserved or restored ecosystem without all of its crucial components is both difficult and expensive. The species of native ecosystem must also occur together in nature. For example, landscaping with an artificial mixture of native species like vine maple, blue bunch wheatgrass, and Jeffrey pine does not produce a native ecosystem. These species are native to different areas within Oregon, but they would not naturally grow together in the same ecosystem. Restoration of native ecosystems must also account for proper structure and appearance. For example, a red fescue lawn does not have the structural complexity and species diversity exhibited by native bunchgrass prairies [Wilson 1991: 17].
Key species – for example, the dominant photosynthesizing plants, the top carnivores, the important decomposers, the nitrogen-fixers – must be present for a native ecosystem to persist and function on its own. To artificially maintain a conserved or restored ecosystem without all of its crucial components is both difficult and expensive [Wilson 1991: 17].
The community as an ecological unit, Barbour, Burk & Pitts 1987
This article provides an overview of types of plant communities and the process of succession in those communities.
In each type of habitat, certain species group together as a community. Fossil records indicate that some of these groups (or very closely related precursors) have lived together for thousands or even millions of years. During that time, it is possible that an intricate balance has been fashioned. Community members share incoming solar radiation, soil water, and nutrients to produce a constant biomass; they recycle nutrients from the soil to living tissue and back again; and they alternate with each other in time and space. Synecologists attempt to determine what is involved in this balance between all the species of a community and their environment [Barbour 1987: 155].
Community concepts and attributes
A plant community is an identifiable stand of plants growing together in a certain spot. Clusters of species, called associations, are often found growing together in several different places within a larger region. “An association is a particular type of community, which has been described sufficiently and repeatedly in several locations such that we can conclude that it has: (a) a relatively consistent floristic composition, (b) a uniform physiognomy [appearance], and (c) a distribution that is characteristic of a particular habitat” [Barbour 1987: 156].
There are opposing views about why particular plant species are often found growing together in a plant community. Thecontinuum viewposits that species distribution is driven individualistically by each species’ particular tolerance to various environmental conditions. By contrast, the association view suggests that a plant community is an integrated whole, whose component species are interdependent.
Whatever the reasons that particular species tend to grow together in stands, however, such stands “exhibit collective or emergent attributes beyond those of the individual populations” [Barbour 1987: 159]. Examples of such community attributes include its vertical structure, canopy cover, species composition and diversity, biomass, productivity, stability, and nutrient cycling, for example.
Succession
Ecological succession is an important concept that helps explain the particular assemblage of plants growing in a given location.
“Plant succession is a directional, cumulative change in the species that occupy a given area through time” [Barbour 1987: 230]. This does not refer to cyclical changes that occur over seasons, nor to changes occurring in response to climate shifts over extremely long time spans like thousands or millions of years. Rather, succession is when the composition of plants at a particular site changes over a period of decades to centuries.
Succession begins when pioneer species colonize bare ground. These first arrivals tend to be opportunists that grow fast, reproduce quickly, and do not live long. The early successional plants start to improve the habitat conditions for other, more competitive plants to then take over, displacing the pioneers. “One of the driving forces behind succession is the effect plants may have on their habitat. Plants cast shade, add to the litter, dampen temperature oscillations, and increase the humidity, and their roots change the soil structure and chemistry. … Both the environment and the community change, and this metamorphosis is due to the activities of the organisms themselves.” [Barbour 1987: 233]
Overtime, slower-growing, larger, longer-living plant species outcompete the earlier successional species, eventually forming a climax community, which is not subsequently replaced by any other community. “Succession often leads to communities with greater and greater complexity and biomass and to habitats that are progressively more and more mesic (moist)” [Barbour 1987: 233]. Such changes result in climax communities tending to be self-sustaining due to efficient nutrient cycling and internal moderation of external fluctuations in temperature and humidity.
The particular plant composition of a climax community depends on the regional climate, as well as local soil conditions and topography, meaning that several climax communities can exist in a given landscape.
Typically, many plant communities coexist in a complex mosaic pattern. That is, one climax community does not cover an entire region. … In [some] cases, the mosaic reflects topographic differences, such as south-facing versus north-facing slopes, basins with poor drainage and fine-textured soil versus upland slopes with good drainage and coarser soil, or different distances from a stress such as salt spray. In such cases, the communities within the mosaic do not bear a successional relationship to one another; they constitute a toposequence. Each community in a toposequence may, in fact, be a climax community [Barbour 1987: 238].
Understanding ecological succession can help us to predict the future vegetation of a site by observing its current vegetation. “It is often possible to estimate a community’s future composition by extrapolation from changes measured in a short time, by comparing other communities that have plants of different ages, or by noting differences between overstory plants and understory seedlings” [Barbour 1987: 231] In some cases, the understory seedlings will later become the canopy, provided the localized conditions support this succession.
Vegetation Ecology: Historical Notes and Outline, van der Maarel & Franklin 2013
These authors define the concept of a plant community through discussion of its evolution. They start by defining the term ‘vegetation’ in a way that may surprise some readers because it excludes plants growing in certain situations. To be considered vegetation, plants need to emerge spontaneously.
Vegetation, the central object of study in vegetation ecology, can be loosely defined as a system of largely spontaneously growing plants. Not all growing plants form vegetation, for instance, a sown corn field or a flower bed in a garden do not. But the weeds surrounding such plants do form vegetation. A pine plantation will become vegetation after some years of spontaneous growth of the pine trees and the subsequent development of an understory [van der Maarel 2013: 1].
Two competing schools of thought regarding the nature of a stand of plants growing together geographically are represented by two early 20th Century botanists. H.A. Gleason observed “that species are ‘individualistically’ distributed along omnipresent environmental gradients and thus cannot form bounded communities” [van der Maarel 2013: 2]. By contrast, E. Clements compared plant community with an integral organism, where the whole was greater than the sum of its parts. During the same time period, the Braun-Blanquet approach was developed, which “paid much attention to the relations of plant communities with the environment and the interactions within communities, which is now incorporated in the concept of ecosystem” [van der Maarel 2013: 2].
The authors state that while individual plant species are distributed according to abiotic environmental conditions, the fact of being co-located with particular sets of other species in a particular environment results in interspecies interactions, which are in fact ecosystem processes (emergent properties).
In conclusion, a plant community is generally recognized as a relatively uniform piece of vegetation in a uniform environment, with a recognizable floristic composition and structure, that is relatively distinct from the surrounding vegetation. Even if the populations of the participating species are usually distributed individualistically in the landscape, they may well interact within the community and build up an integrated unit with emergent properties. At the same time, plant communities can be convenient units for conveying information about vegetation and its environment [van der Maarel 2013: 4].
Vegetation types and their broad-scale distribution, Box & Fujiwara 2013
A vegetation type, or plant community, is identifiable by its distinct appearance compared to other landscape types within a landscape. For example, a grassland and a wetland differ in appearance from each other and from a forest, while a wetland-forest is yet another visibly different vegetation type. Plant species are recognizable by their form, which is related to how the plant functions. For example, in dry environments, plant leaves are more compact with harder surfaces to limit water loss, while plants in wetter environments have larger, “softer” leaves that release water readily when pores open to take in CO2. Such leaves have more surface area for photosynthesis, resulting in faster growth.
This form-function relationship explains why vegetation types differ around the globe. Plant species are adapted to particular climatic conditions according to their proximity to the equator or a coastline, for example, or their elevation.
The geographic regularity of vegetation distribution arises, of course, from the geographic regularity of Earth’s main climatic regions, driven by the global circulation pattern of the Earth’s atmosphere [Box 2013: 466].
Predictive modeling of the potential natural vegetation pattern in northeastern China, Liu et al. 2009
This study uses the concept of Potential Natural Vegetation (PNV), developed in the mid-1900s by German botanist Reinhold Tüxen. Described by the authors as “one of the most successful novelties in vegetation science over the last decades” [Liu 2009: 1313], PNV can be defined as a projection of the natural vegetation that would exist in a given area in the absence of human interference.
“By showing the relationships between environmental variables and vegetation types, maps of the PNV are an important instrument in the study and planning of the environment, and act as decision-support tools for the solutions to environmental issues” [Liu 2009: 1313]. Such maps are informed by studying remnant natural (old growth) vegetation in the area and site observations of the area to be mapped.
Computer modeling can be used to predict “the geographic distribution of vegetation composition across a landscape from mapped environmental variables, such as climate, soils, and geology. When a predictive vegetation modeling is calibrated using observation of vegetation composition taken from mature or ‘climax’ vegetation stands, then potential natural vegetation is portrayed in a predictive map” [Liu 209: 1314].
Focusing on northeastern China, the study identified 16 vegetation types in the region, along with the environmental factors influencing their distribution. Climatic factors included: mean annual temperature, mean temperature of the coldest month, relative humidity, and potential evapotranspiration rate. Topographical factors were elevation and slope.
“Generally, as the elevation increases, the change of temperature and moisture leads to the obvious differentiation phenomenon in vegetation vertical zones. Slope is related to the hydrology (overland and subsurface flow velocity and runoff rate) and potential soil moisture and soil development of a habitat” [Liu 2009: 1315].
They compared the map created by their model to existing vegetation maps of the region. “Visual comparison of the predicted PNV distributions with their actual equivalents indicates a good agreement” [Liu 2009: 1317]. Some modeled vegetation types did not agree with existing maps, however, meaning that “some more important environmental factors may have been missing in the model” [Liu 2009: 1318]. The authors also state that calibrating their model with additional field data on what is currently growing, collected from throughout the region, would improve the model’s accuracy.
The article concludes by stating that a ‘vegetation-environment’ model can help to determine PNV under not only current, but also predicted future environmental conditions.
Conceptualizing communities as natural entities: a philosophical argument with basic and applied implications, Steen et al. 2017
Ecological restoration aims to recreate lost or degraded ecological communities. However, “community” has been a difficult concept to define – should the definition stress dominant species, species interactions, or a subset of strongly interacting species? These authors propose defining community on the basis of co-evolutionary relationships among species.
We propose that an Evolutionary Community is conceptualized as a unique grouping of species, which occur in a given geographic area and are connected by interspecific and abiotic interactions that have evolved over time [Steen 2017: 1021].
By treating communities “as entities that have formed over evolutionary time; this [Evolutionary Community] concept allows for a philosophical platform to help us understand what many conservation and restoration efforts are trying to accomplish” [Steen 2017: 1031]. That is, it offers a way to conceptualize the end goal of a restoration project.A particular evolutionary community could be recreated by assembling the constituent species, resulting in the ecological interactions among the species resuming as before.
What processes cause a group of species to cohere into a community? We argue that the parts of Evolutionary Communities are bound together by interspecific interactions in a shared biotic and abiotic environment, which promote co-evolution and community structure and dynamics. For example, longleaf pine trees are conduits for lightning strikes that ignite a highly flammable understory, often including dropped longleaf pine needles. The resulting ground fires are necessary for reproduction of other species and maintain habitat suitable for others (e.g., gopher tortoises). Gopher tortoises, through the process of burrow creation, provide structure important to other species. The establishment of one or more of the species listed above facilitated the persistence of additional species [Steen 2017: 1025].
Likewise, the demise of one species will negatively affect, or even cause the demise of, other species that depend on it. Thus, the reason to preserve or recreate an integral community is to support the interdependent component species, each of which in turn support the community as a whole.
Bridging ecology and conservation: from ecological networks to ecosystem function, Harvey et al. 2017
This article emphasizes the importance of species interactions as drivers of ecosystem function.
The classic conservation approach is to set aside national parks or to target specific species for protection, based on their rarity or endangered status. However, these approaches can have trade-offs for non-target species, while also potentially failing to protect ecosystem function. The authors, therefore, suggest that species interactions based on their functional significance should be the main focus on conservation efforts.
We propose that a shift in focus from species to interaction networks is necessary to achieve pressing conservation management and restoration ecology goals of conserving biodiversity, ecosystem processes and ultimately landscape-scale delivery of ecosystem services [Harvey 2017: 371].
Species depend on many other species in their communities, either directly or indirectly. An example of indirect dependence is the Phengaris arion butterfly’s need for European rabbits. The butterfly uses ant nests made in the open areas supplied by rabbit grazing for development of its larvae. Thus, no rabbits means no ants, which means no Phengaris arion.
Focusing on species interactions is more meaningful even than measuring species richness (the number of different species), because interactions can disappear – even if both species are present – if either group’s abundance has significantly dropped. The authors offer the example of 59 regionally extinct lepidoptera (butterfly and moth) species of central Europe. Eight of these extinctions were associated with the loss of particular host plant species, which actually occurred after the lepidoptera went extinct.
Focusing on species interactions is more meaningful even than measuring species richness (the number of different species), because interactions can disappear – even if both species are present – if either group’s abundance has significantly dropped.
Thus, strong declines of host plants can have cascading extinction effects on higher trophic levels before the plants actually go extinct, illustrating that interactions can be lost before any actual decline in species richness (plants persisted at low abundance). This illustrates that preserving keystone interactions, rather than species, can be a proactive way to maintain ecosystem integrity in the face of global change instead of allocating resources to already endangered species [Harvey 2017: 372].
There is interdependence among species even between neighboring ecosystems. For example, a manta ray species in the Palmyra Atoll south of Hawaii depends on two species of native trees to maintain its ocean plankton diet. When these trees were replaced with cultivated coconut palms, marine-foraging birds no longer nested on that shore, depriving the coastal waters of the nitrogen runoff from their guano, which had been feeding the plankton population.
The authors recommend that “the main lever to restore or conserve ecological network structure and stability is the management of spatial configuration” [Harvey 2017: 377]. Reflecting on the Palmyra Atoll, for example, it’s clear that a marine conservation plan would be incomplete without considering the nutrient flow from the tree-bird interactions on land.
Interactions among plants and evolution, Thorpe et al. 2011
This review explores the question of whether plant-plant interactions drive evolutionary changes. “If such evolution is common, plant communities are not random assemblages of species.” The topic is under-studied compared to plant interactions with other groups.
Research on plant–consumer, plant–pollinator and plant–disperser interactions has been central to understanding the complex mutualistic and co-dependent interactions among species that structure communities. However, with some notable exceptions, interactions among plants have not been emphasized as processes that contribute to selection and evolution [Thorpe 2011: 730].
“The simplest interactions among plants are direct interactions, such as facilitation, resource competition and allelopathy” [Thorpe 2011: 731]. Facilitation is when one plant protects an adjacent plant, such as from drought and heat by providing shade, for example, or from browsing by being thorny or toxic to herbivores and surrounding the facilitated plant. Allelopathy refers to plants’ release of toxic substances that suppress the growth of another organism, including other plants. In natural communities, any given plant may be interacting with several different plants at the same time.
In natural communities, any given plant may be interacting with several different plants at the same time.
Competition for sunlight, water, and nutrients drives niche differentiation, or the carving out by species of particular spaces or timing within an ecosystem to obtain a share of limited resources. “The exceptionally rich body of ecological literature on the niche is based in part on the idea that competition can drive the evolution of niche differentiation, thus allowing species to coexist” [Thorpe 2011: 732].
Thorpe et al. refer to an example from a 1976 article by Parrish & Bazzaz , who “found that resource partitioning, as estimated from spatial overlap among root systems, was higher in stable prairie communities with a long community history than in early successional old-field communities composed of species without a common history” [Thorpe 2011: 731]. In other words, plants with a long coexistence history more efficiently divvy up resources than do species lacking a common community history.
The primary hypothesis for positive diversity–ecosystem function relationships has been niche ‘complementarity’, the idea that different species or functional groups occupy niches different enough from each other to more fully utilize resources or space, increasing and stabilizing productivity, and making it more difficult for other species to enter the community [Thorpe 2011: 733].
The authors are somewhat inconclusive, however, about what drives niche complementarity (resource partitioning).
We do not yet know whether complementarity is produced by interactions causing evolutionary shifts in niche space (and thus coexistence and more complete resource use) or by sorting of the existing species pool [Thorpe 2011: 733].
Plants can also adapt to one another’s allelopathic substances over time, a fact that contributes to the argument that plant-plant interactions produce evolutionary changes. “Recent experiments raise the possibility that some invaders may exude allelochemicals that are relatively ineffective against neighbors in natural communities, but highly inhibitory to plants in invaded communities” [Thorpe 2011: 734].
Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities, Burghardt et al. 2010
This research evaluates the impact of the invasion of non-native plants in the distribution of lepidopteran (butterfly, skipper, and moth) communities. The authors assert that although the introduction of non-native plants has not resulted in a “global extinction”, they have had a considerable impact on how ecosystems function—they often result in significant bottom-up reductions of energy available in local food webs.
The experiment established four gardens near mature woodlots containing most, if not all, of the native species planted within the treatment. The richness and abundance were then compared for lepidopteran communities found on native versus corresponding non-native congener[4] species of 13 woody plant genera. For example, the genus Acer (maple) was selected for this study because the native and non-native maples were widespread in that area. In separate plots, the researchers also compared native plants and unrelated (non-congeneric) non-native plants for lepidopteran richness and abundance.
The study found that lepidopterans suffer from the replacement of native plants by non-natives, especially when those non-natives are unrelated to any native plant species. The authors explain that “insect herbivores adapted to the chemical challenges [toxic plant defenses] of particular native hosts may be able to adopt a novel plant species as a host if its phytochemistry is sufficiently similar to the original hosts” [Burghardt 2010: 10]. Over the two-year study, lepidopteran abundance and richness were depressed both on congener and (unrelated) non-congener non-native plants, but especially on the latter.
The study found that lepidopterans suffer from the replacement of native plants by non-natives, especially when those non-natives are unrelated to any native plant species.
Specialist lepidopteran species, which require specific diet and habitat conditions to survive, fared worse on non-natives than did generalists, which can eat a variety of foods and survive in many different habitats. The authors note, for example, that “geographically novel congeners were acceptable hosts to less than half of the generalists and only one fourth of the specialists that we found on native congeners in 2009” [Burghardt 2010: 11]. Only 7% of specialist species used non-congener non-natives as hosts.
The authors argue that the loss of lepidopteran diversity and abundance due to the displacement of native plant species with non-natives can ripple up the food chain, reducing diversity at higher trophic levels. Reduced diversity leads to lower ecosystem productivity and stability, thus disrupting the whole system.
The authors argue that the loss of lepidopteran diversity and abundance due to the displacement of native plant species with non-natives can ripple up the food chain, reducing diversity at higher trophic levels. Reduced diversity leads to lower ecosystem productivity and stability, thus disrupting the whole system.
Because insect herbivores are near the hub of most terrestrial food webs, comprising essential food stuffs for an incredible diversity of insect predators and parasitoids, spiders, amphibians, lizards, rodents, bats, birds, and even higher predators such as foxes and bears, it is particularly important to understand changes wrought by non-native plants on this critical taxon [Burghardt 2010: 13].
Impact of Native Plants on Bird and Butterfly Biodiversity in Suburban Landscapes, Burghardt, Tallamy & Shriver 2008
In this study, the insect and bird populations of six pairs of suburban yards were measured. Each pair contained one conventionally landscaped yard containing native canopy trees and a mixture of native and non-native shrubs, grasses and understory trees; and one yard with native species only (canopy, understory, shrub and grasses). The level of plant diversity was comparable between each of the pair; only the proportion of native species differed. The authors found that:
Avian abundance, diversity, richness, and biomass (particularly bird species of conservation concern) were all greater on native properties. Native nesting birds that are mostly dependent on insect populations to feed their young were more abundant on native properties. Lepidoptera [butterfly and moth species] abundance and diversity were also higher on native properties, suggesting that food availability might account for the differences detected in the bird communities between native and conventionally landscaped sites [Burghardt 2008: 223].
These results support the authors’ hypothesis based on an understanding of the co-evolutionary roots of species interactions.
Theory backed by decades of empirical evidence predicts that up to 90% of all species of insect herbivores can successfully reproduce only on plant lineages with which they have shared an evolutionary history [Burghardt 2008: 220].
Native plants improve breeding and foraging habitat for an insectivorous bird, Narango, Tallamy & Marra 2017
This study examined whether non-native plants in residential Washington DC limited the presence of the Carolina chickadee, a local breeding insectivore.
We predicted that areas with more native plants would support more chickadees, and chickadees would forage more often in the most insect-producing native plants [Narango 2017: 43].
The authors had also considered the possibility that non-native plants could promote increases in other food items (e.g. non-native arthropods), keeping overall prey biomass similar between native and non-native plants. What they found, though, affirmed their prediction: native plants produce more caterpillars, which in turn support more chickadees. In fact, the birds avoided foraging in non-native plants, including non-native species of the same tree genera: the chickadees preferred maples native to the eastern US compared to European-origin maples.
Native plants produce more caterpillars, which in turn support more chickadees.
Native plants were more likely to host a higher biomass of caterpillars compared to non-native plants, and chickadees strongly preferred to forage in native plants that supported the most caterpillars. In addition, chickadees were less likely to breed in yards as the dominance of non-native plants increased [Narango 2017: 42].
Also unique to our study is that we measured the probability of caterpillar occurrence between congeneric species (e.g. native vs. non-native Acer [maple]). This is particularly important considering the popularity and invasive qualities of congeneric species in this region such as Acer platanoides and Quercus acutissima. Although non-native congeners support more caterpillars in comparison to plants unrelated to any native species, congeners had a 47% (CI: 34%–59%) lower probability of having caterpillars compared to native species [Narango 2017: 47].
The authors state that local insects are adapted to local plants, presumably due to their shared co-evolutionary history.
This occurs in part because herbivorous insects have adapted to circumvent the phytochemical defenses of particular plant lineages, resulting in a radiation of specialized plant-insect associations. During urban conversion, native plants are replaced by non-native species with novel chemical, physical, and phenological features for which native herbivorous arthropods have few physiological or behavioral adaptations [Narango 2017: 42].
Do non-native plants contribute to insect declines? Tallamy, Narango & Mitchell 2020
The widespread distribution of plants outside of their native range due to human activity is a significant yet underrecognized cause of global insect decline, according to this article. To illuminate the issue, the authors: “examine the evidence for and against the hypothesis that long term changes in the species composition of plant assemblages have contributed to local and global declines in the abundance and diversity of the insect communities dependent upon those assemblages” [Tallamy 2020: 2].
To be sure, insect conservationists have long noted the importance of habitat containing appropriate native host plants, but the widespread replacement of native host plants with non-native species has yet to penetrate the growing literature on insect declines in any meaningful way [Tallamy 2020: 1].
It is not simply the absence of native plants harms plant-eating insects, however, but also the presence of non-natives. While some insects feed successfully on non-native plants, this is the minority. Most either avoid non-native plants, or do use them and are killed or malnourished by doing so. For example,
Swallowworts (Vincetoxicum spp.) are confamilials of milkweeds (Asclepias spp.) and have become invasive in parts of the northeastern United States. Similar phytochemistry between swallowworts and milkweeds can lead monarch butterflies (Danaus plexxipus) and milkweed beetles (Chrysochus auratus) to fatally mistake these chemically protected plants as hosts. The degree to which Vincetoxicum act as ecological traps for these taxa is likely to become more pronounced as the plants become dominant and displace milkweeds in the landscape [Tallamy 2020: 3].
Species that share a particular environment over hundreds or thousands of years evolve in relation to one another. For plant-eating insects, adapting to certain plants meant developing “traits to detect and tolerate plant defenses over time” [Tallamy 2020: 2]. Most herbivorous insects adapted to only a particular set of plants, specializing in feeding on those plant hosts.
The diet of most insects is constrained to a single plant family in any one habitat or location, with dietary specialization even narrower both in many temperate lineages and hyper-diverse tropical lineages. In fact, diet specialization increases with decreasing latitudes, concurrent with theories of increased plant and animal diversity in the tropics [Tallamy 2020: 2].
When native plants are displaced in the landscape by non-native species, phytophagous [plant-eating] insects typically do not recognize the novel host for feeding or oviposition [egg laying], or may be unable to overcome novel plant defenses. The concurrent loss of native plant hosts and dominance of non-native plants can lead to local extirpation of phytophagous insects and thus to changes in the composition and structure of local food webs [Tallamy 2020: 2].
The most likely successful substitute for a native plant is a non-native plant in the same genus or family.
Non-native congeners [members of the same genus] or confamilials [members of the same family] that are similar in foliar chemistry and nutrition, phenology, and morphology, may occasionally serve as novel hosts for herbivorous insects and support higher diversity and abundance than non-native, non-congeners. However, novel use of congeners may increase larval mortality, extend development or pupation time, reduce biomass, and reduce fitness compared to that of native hosts [Tallamy 2020: 3].
The narrower the native plant diet an insect species has, the less likely to tolerate novel, non-native food sources. However, there are more species of specialist insects than of generalists, meaning a larger proportion of susceptible species. Adaptability to exotic host plants also depends on an insects’ feeding habits.
Insects with chewing (mandibulate) mouthparts are typically more susceptible to defensive secondary metabolites contained in leaf vacuoles than are insects with sucking (haustelate) mouthparts that tap into poorly defended xylem or phloem fluids. Thus, sucking insects find novel non-native plants to be acceptable hosts more often than do chewing species [Tallamy 2020: 4].
Considering that there are more than 4.5 times as many mandibulate insect herbivores as haustelate species, there is reason for concern when non-native plants replace native hosts; the largest guild of insect herbivores is also the most vulnerable to non-native plants and the most valuable to insectivores [Tallamy 2020: 5].
“The dispersal and spread of invasive plants has been driven by global trade networks and colonialism” [Tallamy 2020: 6] and, more specifically, from agroforestry, forestry, agriculture, and horticulture.
Although plants have always distributed themselves around the globe, the increased temporal and spatial mobility of humans has resulted in an extraordinary increase in the rate of plant movements and most species’ introductions have happened in the last 200 years. Habitat is rapidly being converted from coevolved native ecosystems into novel assemblages of plants and animals, making the conversion of native plant communities into plant assemblages dominated by non-native species one of the most ubiquitous threats to biodiversity today. The introduction of non-native plants has completely transformed the composition of present-day plant communities in both natural and human-dominated ecosystems around the globe and the magnitude of introductions is staggering. An estimated 13,168 plant species (about 3.9% of global vascular flora) have been introduced and naturalized beyond their native ranges as a result of human activity [Tallamy 2020: 6].
Global exchange and accumulation of non-native species, van Kleunen et al. 2015
The ecological, economic, and social damage of human-mediated dispersal of species into new regions, where they possess the ability to naturalize (become self-sustaining their new homeland), is one of the defining features of the Anthropocene Epoch. Globally, human activity has led to the naturalization of nearly 13,168 plant species (equal in size to the native European flora). The results from this research provide a baseline for monitoring global changes in biodiversity while highlighting the immediate action that has to be taken to comprehend and determine the spread of alien species on an international scale.
The ecological, economic, and social damage of human-mediated dispersal of species into new regions, where they possess the ability to naturalize (become self-sustaining their new homeland), is one of the defining features of the Anthropocene Epoch.
At least 3.9% of all currently known vascular plant species have become naturalized outside their natural ranges as a result of human activity. With the continued practice of international traffic and trade and globalization, the likelihood of more and more species being introduced and getting naturalized outside their native range is high.
To assess the accumulation of naturalized species in each continent as well as which continents have been the major donors of alien naturalized plant species globally, the researchers used a novel database, Global Naturalized Alien Flora (GloNAF), in addition to the data on the origin of naturalized species and estimates of the number of native species per continent. They found that when not taking into account the differences in total area, North America has accumulated the highest number of naturalized species (n=5,958). However, when considering the difference in total area, Australasia (a region comprising Australia, New Zealand, and neighboring islands) was found to have more extra-continental species than North America.
One possible explanation is that Australia’s long biogeographical isolation and drying climate have resulted in a native flora that is phylogenetically distinct, but not well-adapted to exploit the novel habitats created by European settlers [van Kleunen 2015: 101].
The major donors of alien species are Europe and temperate Asia, while North America is also a significant donor.
Ecological and evolutionary consequences of biotic homogenization, Olden et al. 2004
Anthropogenic environmental change and global dispersal of a wide variety of species outside their native ranges has expanded the range of “cosmopolitan,” non-native species and shrunk the range of regional and endemic species. “This replacement of specific native forms by generalist non-natives in space and time has mixed the taxonomic composition of once disparate biotas, an occurrence termed ‘biotic homogenization’” [Olden 2004: 18].
The authors explore the effect of this “global erosion of regional distinctiveness” [Olden 2004: 18] at three levels: Genetic homogenization reduces genetic variability within species or among populations of species, while taxonomic homogenization reduces distinctiveness among communities. Functional homogenization refers to a reduction of functional traits within an ecosystem. The identity of species making up a community, along with their respective functional traits, determines “ecosystem functions (such as nutrient retention or energy flow)” [Olden 2004: 20], so that narrowing species compositions risks diminishing ecosystem function.
A decrease in functional diversity might reduce overall community and ecosystem functioning, stability and resistance to environmental change by simply narrowing the available range of species-specific responses. Consider a severe drought that strongly affects a subset of species in a community that has (or lacks) a particular suite of functional traits. Historical communities, with much greater breadth in functional space, should exhibit higher resistance or resilience when compared with homogenized communities [Olden 2004: 20].
Genetic homogenization occurs when two distinct locally adapted populations of the same species interbreed. It also occurs when a single variety (such as captive fish bred in a central location) are released in many places to replenish dwindling native stocks. While such mixing has the potential to increase species diversity, this outcome is not assured.
Intraspecific hybridization can homogenize the unique characteristics of geographically distinct populations, as well as compromise the fitness of individuals by disrupting local adaptations [Olden 2004: 19].
This book draws lessons from ecological succession theory to inform ecological restoration, stating that: “restoration is fundamentally the management of succession” [Walker 2007: vi]. The latter is the natural process by which plants first colonize “new” land (post landslide, glacial retreat or volcanic eruption, for example) or degraded land, and over time develop into mature ecosystems through a series of changing plant communities. Ecological restoration is a human-led initiative to restore functioning ecosystems, or at least vegetation, on land degraded through human activity. The ultimate goal of restoration is to “establish a self-sufficient ecosystem that requires minimal or no continuing human inputs in order to provide a continuing supply of goods and services” [Hobbs 2007: 177].
Effective ecosystem restoration requires ecological knowledge. Likewise, the outcomes of such projects demonstrate our comprehension, or lack thereof, of ecological concepts: “Restoration is the acid test of our ability to understand not only how ecosystems are assembled and held together but also how they change over time” [Walker 2007: vi]. The authors contend, however, that restoration projects are more often guided by engineering, horticulture, and agronomy than by ecology. Aiming to clarify the ways in which ecological succession theory can and should inform restoration, this book poses the question: “What is the minimum amount of biophysical and successional information needed to restore a specific landscape or area” [Walker 2007b: 2]?
Succession comprises many ecological processes that underpin all ecological restoration and ecological restoration is a manipulation of these processes to achieve its goals. This means it is essential to understand how succession operates, and when and how to manipulate it [Prach 2007: 121].
Restoration can explicitly embrace a hands-off approach, where land is simply left to repair itself through natural ecological succession. On the other hand, understanding the successional process allows manipulation of various stages to speed up the process. For example, in the first stage of primary succession “winds deposit dust, pollen, seeds, and insects crucial to reducing infertility” [del Moral 2007: 23], on bare, inhospitable ground. Tough pioneer plants are able to establish then create shade, trap sediment, and deposit organic matter when they die, creating slightly better conditions for the next wave of colonizing plants. To mimic this first stage of site “amelioration”, the site can be physically manipulated by reshaping the ground for improved drainage or adding organic matter, for example.
Biological manipulation involves sowing or planting local/native varieties of later successional species that may not be otherwise present in the area due to human transformation of the broader landscape. While earlier successional species tend to have small, easily transported seeds, the later successional species (such as large canopy trees) that are often the target of restoration efforts often have large, less mobile seeds. Thus, if those plants are not present in the immediate environment as seed stock, they may never establish in the restored site without human assistance.
Near-Natural Silviculture: Sustainable Approach for Urban Re-naturalization Assessment Based on 10 Years Recovering Dynamics and Eco-Benefits in Shanghai, Guo et. al 2015
As one of China’s major cities, Shanghai’s natural sub-ecosystem[5] has suffered drastic damage due to human activities and urbanization. Although urban re-naturalization has gained attention from city leaders, urban tree planting has largely consisted of two methods with limited ecological potential. One favors fast-growing monocultures to produce timber products and other benefits, while the other approach is to plant non-native species for decorative purposes. The authors believe the restoration progress of the natural sub-ecosystem could be further improved by adopting the “near-natural” method based on the concepts of potential natural vegetation[6] and ecological succession.
The near-natural forest uses all native species and aims to create a complex structure with high biodiversity, high biomass and multiplayer canopies. It was adopted successfully in many countries, but the authors thought long-term studies of these forests were lacking. Therefore, they conducted a 10-year study at a near-natural forest established in 2000 in Pudong New Area of Shanghai to investigate the effectiveness of the forest in providing ecological benefits.
Results showed that the near-natural forest had higher sustainability value than artificial (“even-aged, managed”) forest in Shanghai based on its ecological and economic benefits. The high tree density and multiple vertical structures of the forest improved the air quality and soil fertility and decreased the concentrations of air bacteria and dust. It also had a much lower planting and maintenance cost than artificial traditional methods. Although the near-natural forest could not transcend the benefits of the natural forest, the study successfully proved its important role in urban re-naturalization by bridging the difference between the artificial and natural forests.
Results showed that the near-natural forest had higher sustainability value than artificial (“even-aged, managed”) forest in Shanghai based on its ecological and economic benefits
Over the course of the study, authors discovered a potential limitation of the approach, at least in its application in Shanghai. They observed high evergreen seedlings mortality, attributable to over-exposure to sunlight. Therefore, in subsequent plantings in 2003 and 2004, the authors modified the approach to optimize it to local conditions:
The key to the new method is to create a mixed deciduous–evergreen community by simultaneously planting shade-tolerant evergreen broad-leaved species and light-demanding deciduous broad-leaved species, but using smaller individuals for the former and larger individuals for the latter to form a multilayer vegetation structure. The shade-tolerant evergreen species benefit from the rapid growth of the light-demanding deciduous species, which offer shade and nutrients in the form of litter layer-based fertilizer, improving the soil for the evergreen species [Guo 2015: 5].
Overall, they suggest that the near-natural forest is a very sustainable method to be applied in Shanghai.
Tree planting is not a simple science, Holl & Brancalion 2020
Well-planned tree-planting projects are an important component of global efforts to improve ecological and human well-being. But tree planting becomes problematic when it is promoted as a simple, silver bullet solution and overshadows other actions that have greater potential for addressing the drivers of specific environmental problems, such as taking bold and rapid steps to reduce deforestation and greenhouse gas emissions [Holl 2020: 580].
Some of the pitfalls to avoid in tree planting initiatives, according to the authors, include:
Use of non-native species, which does not result in a true forest and can result in ground water depletion in arid environments.
Planting trees in historic grasslands and savannas, harming those native ecosystems and species.
Abandoning trees after they are planted, which can result in high mortality due to insufficient water for developing saplings, being shaded out by faster growing herbaceous plants, grazing, or being re-cleared.
Planting trees in agricultural land, which risks pushing crop production into native forest land, which is then deforested.
The authors insist that reforestation takes careful planning, stakeholder engagement, clear goal-setting, and long-term monitoring and adaptive management of planted tree stands to ensure their survival. Above all, existing mature, native forests should be preserved.
The authors insist that reforestation takes careful planning, stakeholder engagement, clear goal-setting, and long-term monitoring and adaptive management of planted tree stands to ensure their survival. Above all, existing mature, native forests should be preserved.
The first priority to increase the overall number of trees on the planet must be to reduce the current rapid rate of forest clearing and degradation in many areas of the world. The immediate response of the G7 nations to the 2019 Amazon fires was to offer funding to reforest these areas, rather than to address the core issues of enforcing laws, protecting lands of indigenous people, and providing incentives to landowners to maintain forest cover. The simplistic assumption that tree planting can immediately compensate for clearing intact forest is not uncommon. Nonetheless, a large body of literature shows that even the best-planned restoration projects rarely fully recover the biodiversity of intact forest, owing to a lack of sources of forest-dependent flora and fauna in deforested landscapes, as well as degraded abiotic conditions resulting from anthropogenic activities [Holl 2020: 581].
“The tamarin is unable to do anything to save its own species. And we, human beings, are the ones who are destroying their environment,” says conservationist Gabriela Rezende. “So, when I got the opportunity to see this animal in the wild, I felt partly responsible for its future.”
Rezende works with the Institute for Ecological Research in the Brazilian state of Sao Paolo to create ecological corridors connecting the forest fragments where the world’s only 1,800 black lion tamarin live in isolated populations. Since 1984, the institute has worked to protect this small primate species, which had reached a low point of 100 individuals and was listed as “critically endangered.” In addition to research and forest restoration, the institute also does environmental education with the local communities. This includes collaboration on nine tree nurseries administered by local people as small businesses that also provide school kids the chance to learn about local forest species that will be planted in corridors.
Leveraging a state policy requiring 20% of privately owned property to be in nature reserves, Rezende worked with landowners to identify patches to be restored that would physically connect forest fragments. Once corridors are complete, the total amount of land in connected habitat will be 111,000 acres. Rezende estimates the black lion tamarin population could increase 30% once it’s able to use the whole forest corridor. The restoration project will benefit other species too, including anteaters, tapirs (a pig-like animal with a short trunk), pumas, and ocelots (another wild cat species).
The Belize government approved a plan in February 2018 to create a 110-square-kilometer biological corridor connecting two nature reserves in the northeast of the country. This outcome resulted from collaboration among NGOs, the government and private property owners. The latter agreed to conserve (to not deforest or otherwise degrade) the parts of their land that would become part of the wildlife corridor. In exchange, the government would not collect taxes on this land. This corridor, which was initiated in the context of the larger Mesoamerican Biological Corridor project, is meant to protect jaguars, cougars and tapirs, among other wildlife.
One of the world’s largest corridor projects is the Mesoamerican Biological Corridor (MBC). Initiated in the 1990s, the MBC aims to connect protected areas between southeastern Mexico and Panama [Meyer 2019: 2].
The ecological functionality of the MBC has not been much assessed, in part because direct approaches to measuring connectivity are costly and challenging. In this study, researchers used a simpler, indirect approach to measure forest connectivity through Panama for nine mammals. Using camera traps (cameras that are automatically triggered by a change in some activity in the vicinity, like the presence of an animal), they documented the presence (or absence) of these mammals in 28 forest sites along the Atlantic coast. The corridor was presumed to be functioning for animals whose presence was established across the entire length of the monitored range.
The species monitored in this study are forest specialists, including ungulates, carnivores and an insectivore, all of which are threatened by habitat loss and hunting, some more than others. Of the 43% of land in Panama that is forested, 44% is protected, mostly along the Atlantic coast. Steady economic development threatens remaining ecosystems with investments in large infrastructure projects, real estate, mining, tourism, and energy.
Large mammals are an indicator species for the success of conservation efforts. This is because:
Large mammals are generally at a higher risk of extinction in disturbed landscapes than other taxa because their large home ranges and low population densities at broad spatial scales mean their populations are more likely to be fragmented and because they are heavily hunted [Meyer 2019: 3].
The researchers found that even the four most prevalent species in the study are susceptible to population fragmentation by any further habitat loss.
We found that there was little connectivity for white-lipped peccary [a pig-like animal] and white-tailed deer and that, although 4 of the species (collared peccary, red brocket deer, puma, and ocelot [a wild cat]) occurred in most of the sites, a small decrease in connectivity of 20% would disrupt their continuous distributions across Panama. White-lipped peccary, giant anteater, white-tailed deer, jaguar, and tapir [a pig-like animal with a short trunk] had lower probability of occurring in all the sites and were therefore even more at risk of connectivity loss, as evidenced by >1 connectivity gap. This indicates the MBC may not function for the majority of species, especially considering we did not account for potential effects of hunting, which would make connectivity even more challenging [Meyer 2019: 8].
Citing imminent development projects, such as a new road that will pass through the forested northern coast and associated large hotel projects, the authors predict that ongoing loss of connectivity is likely. Moreover, the deteriorating condition of the corridor in Panama bodes poorly for the MBC overall.
The disruption of connectivity between tropical forests in Central America, and hence the possible separation of mammal populations, is an indicator of the overall functioning of the MBC for wildlife [Meyer 2019: 11].
Written by an anthropologist working in Central American conservation efforts for more than 10 years, this article describes the Mesoamerican Biological Corridor (MBC) project as having succumbed to a neoliberal agenda. Although originally spearheaded by Central American environmentalists, the notion of cross-border environmental collaboration was adopted by the World Bank and large international conservation organizations working in Central America in the 1990s. In the hands of these international giants, the biological corridor initiative became a bureaucratic, top-down project, deaf to the voices of local communities.
With all this new bureaucracy, a broad and unfocused agenda, and the challenges of high-level political coordination, the MBC quickly lost its potential to inject a stronger environmental justice component into regional biodiversity conservation programs.
Indeed, the MBC that emerged from the World Bank’s incubator was decidedly more business-oriented than initial proposals for Central American environmental coordination at the 1992 Earth Summit [Grandia 2007: 486].
In this context, the MBC’s conservation efforts have focused more on securing land for protected parks and less on community-based initiatives. The author suggests that in addition to land protection, the MBC should engage farmers in capacity building for eco-agriculture with a view toward achieving landscape-wide ecological connectivity.
The corridor approach might also draw greater attention to the agrarian contexts outside of parks, which may be just as ecologically important as what happens inside parks. By bringing agricultural systems into conservation debates, corridors may present new opportunities for supporting fair-trade projects and other small-scale agroforestry systems compatible with conservation. In other words, corridors could offer a method for moving beyond protectionism to embrace a mosaic vision for conservation that includes local people more explicitly. Corridor planning frameworks also could provide more democratic conservation forums [Grandia 2007: 484].
At the end of the 1980s, as a period of severe conflict in Central America was winding down, most countries in the isthmus signed the Charter Agreement for the Protection of the Environment, which established a sustainable development commission. At the same time, the “Central American Protected Areas System (SICAP) created approximately 11.5 million hectares of protected areas throughout the region” [Dettman 2006: 18].
This paved the way for international attention and investment in what became the Mesoamerican Biological Corridor (MBC). The original intention was to promote biodiversity and economic development in tandem through investment in local projects. However, in the 2000s, the international coordinators of the MBC shifted the focus from biodiversity protection (although the establishment of ecological corridors remains an objective) to a greater emphasis on economic development. This author explains that the institution’s decision-making process is overly top-down, and would benefit from input from local people who are implementing projects on the ground.
The concept of sustainable development presumes that human economic systems and overall wellbeing depend on functioning ecosystems. Therefore, ecological rhythms should not be transgressed to the point that they fail to provide the vital services needed today and in future generations.
According to this model, economic development becomes a necessary but insufficient condition for society to progress [Beauvais & Matagne 1999: 6, translated].
Costa Rica holds at least 5% of the world’s species, in spite of making up 0.03% of its land surface. As an isthmus, Costa Rica is influenced by weather patterns from two oceans, as well as a north-south migration route. In addition to this, its mountainous terrain creates a heterogenous mosaic of habitats and niches. However, the country has been severely deforested. Forest covered 66% of land surface in 1940, and only 25% by 1987; the loss of forest led to extreme erosion.
As presented in this article, an ecological corridor consists of at least two protected ecosystem patches that are connected by a protected vegetated strip of at least a few kilometers in width, and the whole area surrounded by a buffer zone. Multiple units of two connected patches could in turn be connected, stretching into a corridor that the whole length of the country. A green Costa Rican corridor could connect to green corridors in adjacent countries, ultimately recreating the entire isthmic corridor that once existed.
However, the tone of this article is not optimistic about conservation, citing several political obstacles to conservation and ecosystem restoration. According to the authors, a combination of neocolonialist pressure, poverty, corruption, and capitalistic interests allow for trees to be cut even in protected areas and prevent the establishment of new protected areas and corridors.
The fate of biodiversity within protected areas is therefore inextricably linked to the broader landscape context, including how the surrounding agricultural matrix is designed and managed [Harvey 2007: 8].
Rather than discussing ecological corridors per se, this article emphasizes the importance of a whole-landscape approach to biodiversity conservation. Pointing out that protected nature reserves are weakened when isolated, these authors focus on the role of the entire surrounding agricultural matrix for restoring connectivity.
In contrast to the prevailing trend of managing protected areas and productive lands separately, we propose integrated landscape management in which conservation and production units within the agricultural matrix are managed jointly for long-term sustainability. We do not advocate agricultural intensification to spare further forest conversion because this approach is unlikely to have the intended outcome, for reasons discussed. Instead, conservation efforts should be based on the recognition that how agriculture is conducted and how different land uses are distributed spatially and temporally determine the region’s biodiversity. Lasting conservation will therefore require alliances among conservation biologists, farmers, and land managers to actively plan the future of Mesoamerican landscapes [Harvey 2007: 9].
The sections of the agricultural matrix the authors prioritize for biodiversity conservation include areas near riparian and other key ecological corridors, and they recommend leveraging support for the Mesoamerican Biological Corridor to spur regional action. Priority conservation areas are also more likely to encompass landscapes with a high diversity of indigenous and traditional cropping systems than those dedicated to industrial agriculture because “the chances of reconciling farming and biodiversity conservation there [agro-industrial systems] are slim” [Harvey 2007: 10].
The authors argue that, in contrast to large-scale, export-oriented industrial production, small-holder and indigenous agricultural systems are more compatible with biodiversity conservation, increased food production and rural income. The authors propose economic and regulatory instruments and greater regional collaboration to enhance native tree cover on farms, promote traditional, ecologically based farming practices, and to protect remaining intact habitat and restore degraded lands. The overarching vision is to accomplish conservation and agricultural production objectives for the region in mutually reinforcing ways.
The fate of biodiversity within protected areas is therefore inextricably linked to the broader landscape context, including how the surrounding agricultural matrix is designed and managed [Harvey 2007: 8].
In spite of its obvious benefits, agriculture, which covers one third of the Earth’s land surface, damages biodiversity and ecosystem services. In some regions, land degradation and depletion of water resources from irrigation have been so great that historical levels of food production in these regions risk decline. Some areas of previously productive farmland will likely need to be retired from use. Within this context, maintaining and enhancing natural corridors and promoting semi-natural, multifunctional landscapes can significantly contribute to recovering biodiversity and mitigating air and water pollution.
Using California’s San Joaquin Valley (SJV) as a case study, this paper illustrates a pragmatic approach to incorporating ecological corridors into working landscapes. The authors offer a new analytical approach that simultaneously incorporates resource-constrained (water, in this case) land-use change (LUC) modeling within the planning and optimization process. The goals are to simultaneously:
Meet water-use-reduction policy goals for the area under study within the next two decades
Identify lands for retirement that are (1) likely to be retired anyways and (2) offer high-value habitat for native species and biodiversity.
Over the past century, SJV has been transformed into one of the largest agricultural economies in the world. However, this economic success has been costly to the SJV in several ways, including:
Damaged infrastructure: high rates of groundwater extraction in the SJV have led to groundwater overdraft and unreplenished aquifers, resulting in large-scale land subsidence. Most of the subbasins in the SJV are categorized as critically overdrawn, and some regions have sunk over 8 meters since the early 20th century; this land subsidence further imperils water availability and quality by impacting water storage and delivery infrastructure.
Decreased human health, as a result of impaired air and water quality, leading to chronic health problems
Threats to wildlife and biodiversity; for example, some species have lost up to 98% of their habitat range, and over 35 native species are listed as threatened or endangered
“In response to these challenges, and amid significant drought-driven fallowing, California passed the Sustainable Groundwater Management Act (SGMA), which … obligates locally governed groundwater subbasins to develop plans that will achieve sustainable groundwater use over the next two decades” [Bryant 2020: 2]. To meet these requirements, many subbasins will meet with severe groundwater pumping restrictions. If these areas are not able to coordinate their pumping activities and augment water supplies, the SGMA may require a reduction in cultivation area through fallowing or permanent retirement.
Given the likely retirement of 86,000 ha of irrigated agricultural land, the authors explore spatial optimization of retired land for conservation efforts. They find that a key strategy is the identification of areas that were destined for retirement from cropping which could be shifted to restoration and habitat enhancement, as well as possibly shifting some areas destined for retirement that have “low habitat value” with regards to wildlife for areas with “high habitat value.” Priority restoration areas identified in this analysis include many that are contiguous and located near designated wildlife areas.
Importantly, the analysis presented here is “explicitly organized to help inform engagement between conservation actors and agricultural land managers about how habitat goals can be achieved in ways that benefit communities in the SJV” [Bryant 2020: 3]. The potential positive futures indicated by such analysis can be used to identify opportunities for collaboration between the conservation and agricultural communities, with a goal of guiding land use change toward achieving multiple benefits, such as recovery of imperiled natural communities, resilient agricultural production, and improved public health outcomes.
While it poses a great challenge, the impending transformation in the SJV also presents an opportunity to proactively shape the landscape in ways that not only ensure agricultural and water sustainability, but also achieve many other socio-ecological goals, such as biodiversity protection and improved human health. However, given that achievement of many of these objectives is determined by where things happen on the landscape (rather than simply the aggregate amounts of cultivation, retirement, or restoration), stakeholdersneed a systematic way to integrate these objectives to informmulti-benefit spatial planning [Bryant 2020: 4].
Produced by a regional consortium on the environment in Brittany, France, this report describes the ecological value of woody strips encircling agricultural fields and enmeshing the countryside, their decline, and ways to incentivize their protection.
Brittany is a heavily agricultural region that also features a long stretch of coastline where urban development and expansion is ongoing. Due to mechanization and enlargement of farm fields, average parcel size has increased since the 1950s, shrinking the extent of woody hedgerow (“bocage”) between fields. Between 1996 and 2008, the total length of hedgerow decreased 12%. This change is concerning because Brittany is already one of the most fragmented and least wooded parts of France.
The report explains the value of the bocage is its provisioning of habitat, connectivity between habitats, biodiversity, erosion control, groundwater recharge, and flood mitigation. Half the population of Brittany lives in areas susceptible to flooding. Furthermore, at least five endangered animal species depend on the habitat created by the bocage. Protection of this woody network is key to remedying both problems, while also providing direct benefits to farmers, such as habitat for pest predators.
The form and shape of the bocage varies throughout the region, but can include grasses, bushes and/or trees, forming one or more layers of vegetation; and heterogenous landscape features such as berms, ditches, logs, and rocks/boulders, which create microhabitats.
The network of hedge and berms, accompanied by fields, ponds and wetlands, constitutes an important natural environment because of its heterogeneity and potential for complex exchanges. It has the particularity of being able to reach a myriad of increasingly isolated natural spaces in the heart of a changing agricultural countryside subject to ongoing urbanization. Similar to a forest edge environment, the richness of the bocage can be explained by the diversity of habitats it adjoins [OEB 2018: 8, translated].
Regulatory and incentive programs payments have sought to encourage farmers to preserve their hedgerows. However, the authors suggest that a stronger economic valuation of these linear woods is needed to protect and expand them. They suggest strategies for stimulating the market for firewood and other products harvested from sustainably managed hedgerow, where biodiversity protection is an explicit aim and co-benefit.
This article reviews the historical development of two pieces of environmental legislation in France – the use of the “mitigation hierarchy” to assess and limit environmental impact in project development and the promotion of ecological corridors. Theoretically, these two laws overlap when urban development projects in proximity to areas of ecological significance use the mitigation hierarchy (avoid, reduce, compensate) to ensure these zones are protected within the scope of the project.
1976: “Protection of Nature” law in France introduced the mitigation hierarchy, aiming to avoid or reduce harm to the environment, or to compensate if harm is unavoidable.
1992: Concept of “biodiversity” entered public discourse internationally, following the Earth Summit in Rio, Brazil.
1996: France ratified European ecological corridor strategy.
1999-2000: Concept of “sustainable development” emerged in France.
2004: National strategy for protecting biodiversity adopted.
2007: “Grenelle de l’Environnement” meeting created the “Trame Verte et Bleue” (TVB) policy (green and blue infrastructure, encompassing ecological corridors)
2016: Biodiversity law enacted, creating national agency and regional committees on biodiversity
In spite of this policy evolution, commitment to ecological corridors has yet to move from a “TVB papier” to a “TVB de projets et d’action.” In other words, much discussion and mapping efforts have not yet resulted in the development of the imagined ecological corridor network. The authors speculate as to why this is so, explaining that the resources and coordination needed for enforcement are lacking. Even though “the creation, preservation and restoration of ecological connectivity” has been integrated into urban planning code, such considerations are often sidelined. Furthermore, definitions are vague: the objective of the TVB is the “good condition” of ecological continuity, but “good condition” is not defined. Lastly, taking action in defense of ecological continuity requires pro-active collaboration among levels of government from local to regional to national.
The authors propose better integration of these to policy tools. For example, the TVB designates certain non-protected areas throughout the country that are ecologically functional and serve a role in the eco-corridor network as key areas to “preserve.” With better communication between this TVB framework, the mitigation hierarchy could be applied at the level of “avoiding” harm to places designated as preservation priorities, but lacking formal “protected” status. In projects where harm is unavoidable, the mitigation hierarchy could be applied at the level of “reducing” harm to maximize the percentage of remaining green space as well as the permeability to wildlife of the built structures (such as passageways through fences). The “compensation” level of the mitigation hierarchy could be applied in the context of regenerating ecosystem function to areas designated in the TVB schema as needing ecosystem restoration.
The authors note that advocates for the TVB are clustered at the national level and within research institutions, while the people responsible for urban planning decisions are local and are not necessarily well versed in the scientific framework for the TVB. Local actors tend to focus on priorities other than ecological continuity. One measure to address this, according to the authors, would be the training of local “relays” to transmit knowledge of ecological principles vis a vis the TVB to local urban planners.
Spurred to action by the European Union and a vision for a pan-European ecological network, France encoded the idea of the “trames vertes et bleues” into law in 2009. The national government worked with all the regional governments to develop maps showing areas with the highest levels of biodiversity. This includes protected areas, stretches of coastline, riparian zones, woods, and other undeveloped areas, whether public or private. The maps also show ecological corridors – both those in good condition needing to be preserved, and those that are highly degraded and requiring restoration.
The regional maps are meant to be integrated into urban planning at the level of city and county (department). Rather than being a regulatory tool, the maps are an information source allowing urban development to proceed in a way that limits impact on biodiversity. The ecological corridor initiative is designed as an invitation and encouragement to local governments, organizations, businesses and individuals to collaborate and to act in favor of biodiversity.
The preservation and restoration of ecosystem connectivity entails acting everywhere possible: in rural environments, in aquatic ecosystems and in urban areas [MTES 2017, translation].
While not an ecological corridor per se, the Natura 2000 network is the largest coordinated network of conservation areas in the world. Covering 17.9% of Europe’s land area and nearly 10% of the continent’s marine areas, the network includes 27,852 sites with an area of 1,358,125 km2. The terrestrial portion of the Natura 2000 network is mostly covered by forests and transitional shrublands. It also includes grasslands and wetlands, as well as pastures, cropland and a small amount of artificial surface (developed/built land).
Member States need to ensure that sufficient protection and appropriate measures are implemented in Natura 2000 sites for habitats and species of community interest and that they form a functional network [EEA 2020: 109].
However, the sites are not strictly protected by virtue of being part of the network. In fact, the sites include a variety of land uses.
Within the network, arable land and permanent crops have increased, while grasslands and forests have decreased. … Pastures and mosaic farmland (with approximately 18 %) and inland wetlands and water bodies (with approximately 10 %) have been extensively transformed into arable land and permanent crops both inside and outside the network. Recent research has shown, however, that high nature value (HNV) farmland inside Natura 2000 sites is less likely to be converted into artificial surfaces than such farmland outside the network and is more likely to maintain its pattern of mosaic farming [EEA 2020: 113].
This assessment of the network’s effectiveness found that “species and habitats are more likely to have a good conservation status if they are well covered by the Natura 2000 network” [EEA 2020: 121]. However, limited monitoring inside and outside the network prevents a more detailed analysis of Natura 2000’s effectiveness. Furthermore, due to a limited implementation of conservation measures, the network’s potential has not yet been fully “unlocked,” according to the report.
To improve Natura 2000’s potential, the authors recommend, among other measures, improving connectivity between protected areas. Noting that sites chosen for inclusion in the network are often motivated by economic rather than ecological interests.
Incoherent planning and site selection approaches between and within Member States has led to insufficient functional connectivity and spatial connectedness between neighboring countries and habitats and gaps in coherence within Member States. This highlights the need to increase connections between protected areas and the level of protection beyond the site [EEA 2020: 122].
Also recommended is increasing stakeholder participation, such as through citizen science monitoring initiatives, and better integration of biodiversity protections into other policy domains.
The resulting low awareness of the diverse benefits produced by the Natura 2000 network is often compounded by a long-standing conflict between economic or political interests and conservation goals. There is thus an urgent need to increase coherence between biodiversity policy and other policy areas, such as in the fields of agriculture and economic and rural development, and create a more integrated approach to address potential conflicts and trade-offs between various interests while fostering synergies [EEA 2020: 124].
The report’s summary conclusion recommends increasing marine and terrestrial conservation areas in the Natura 2000 network to 30% each, strictly protecting these areas, and improving connectivity among them.
Conceptually the inverse of wildlife corridors, fences aim to disconnect. They are built to separate people across national borders, livestock from predators, to delineate property lines, and even to protect wildlife conservation reserves. Globally, fences are ubiquitous, more prevalent even than roads, and proliferating. Yet their ecological impact is relatively unstudied.
Fences are often framed as a management tool rather than a globally significant ecological feature, and they are a notable omission from efforts to map global infrastructure, including the human footprint [McInturff 2020: 971].
This analysis reviews 446 studies starting from 1948 on various types of fencing to assess impacts; however, most of the studies focus on the effect of fencing on particular species (specifically, those the fencing is meant to protect), rather than on multiple species, communities or ecosystems.
Conservation and restoration fences, for example, have support within the literature for their beneficial effects for wildlife and sensitive plant species for which they are built, making such species “winners” in the fencing game. On the other hand, there is a critical lack of information on species that are not the targets for which fences are built, and our review shows that only 10.8% (48 of 446) of studies focus on nontarget species [McInturff 2020: 975].
While fences aiming to protect particular species usually achieve that goal, they inevitably hurt other species.
… often the clearest winners because of fencing are the species that humans value most, whereas losers are inevitable but may remain invisible [McInturff 2020: 975].
Broadly speaking, fences favor generalists and disturbance specialists, many of which are invasive, as well as small and small-ranged, nonmigratory species. Fences therefore heavily restrict what makes a species a winner [McInturff 2020: 975].
The deleterious effects of fences include: impeding migration, reducing gene flow between populations, restructuring community composition and obstructing interspecies interactions, such as between predators and prey. These community-level changes can have ripple effects in the ecosystem. For example, livestock fences effectively excluding dingoes in Australia led to this large predator’s eradication. “Without dingoes, researchers have tracked a continental-scale mesopredator [mid-level predator] release that has altered biodiversity and habitats over enormous areas of Australia” [McInturff 2020: 979].
While fences limit certain interspecies interactions, they concentrate others:
Even where conservation or restoration fences seemingly protect whole habitats, research still points to differential outcomes for constituent species. In addition, pathogens and parasites may spread more rapidly where species interactions are concentrated within reserves. In central Kenya, for example, smaller fenced reserves produced higher gastrointestinal parasite infection rates among impala [McInturff 2020: 977].
The authors recommend a greater research focus on the cumulative ecological effects of fencing, policy that limits fence building and encourages fence removal or fence design that is more wildlife-friendly. They caution that fencing is among the major drivers of anthropogenic change.
As fencing continues to rapidly proliferate, there is potential for a dangerous future in which fences simultaneously alter ecological processes at multiple scales, likely producing more losers than winners, and potentially resulting in ecosystem state shift or collapse [McInturff 2020: 977].
Livestock fences effectively excluding dingoes in Australia led to this large predator’s eradication. “Without dingoes, researchers have tracked a continental-scale mesopredator [a mid-level predator] release that has altered biodiversity and habitats over enormous areas of Australia” [McInturff 2020: 979].
‘Road verges’ are strips of land on either side of roads and highways that are on average 3-4m wide, but can be as narrow as a few centimeters or many meters wide. “Road verges are commonly grassland habitats, but can be shrubland, forest or artificial arrangements of trees and horticultural plants, and we use the term also to include bare earth and freshwater bodies (e.g. ditches)” [Phillips 2019: 489]. They can also be barren ground or ditch. Not all road verges are managed; when management does occur, it is typically geared toward safety – clearing vegetation to enhance visibility.
There is currently an estimated 36 million linear kilometers of road network in the world, the length of which is expected to increase by 70% by 2050; thus, the total area of road verges will increase as well. “Road and road verge construction will displace habitats and cause many negative ecological and social impacts” [Phillips 2019: 494]. However, there is potential to mitigate that impact by maximizing the ecological value of road verges. Currently, “there may well be 270,000 km2 of road verge globally (0.2% of land), which is similar to the total area of the United Kingdom” [Phillips 2019: 492], with this surface area expected to grow.
While roads run like a network of veins across landscapes, causing widespread negative ecological impacts to adjacent areas, road verges form a parallel network and have the potential both partially to mitigate negative impacts of roads and to deliver environmental benefits [Phillips 2019: 490].
Where roads cut through natural habitat, the road verges will represent a net loss of biodiversity. By contrast, verges can increase biodiversity in highly degraded environments such as cities or industrial farmland. Furthermore, because of the growing urban population, the importance of natural and semi-natural environments will be increasingly important. Road verges designed to maximize ecological value thus have an important role to play in the health and wellbeing of urban residents.
Road verges might increase connectivity in highly modified urban and agricultural landscapes if road verges of suitable size, habitat quality and continuity are created alongside roads, at least for species that are highly mobile or able to persist in narrow, linear habitats [Phillips 2019: 495].
While roads often act as barriers to wildlife and ecological connectivity, ecological corridor design could benefit by taking into account the potential benefits of road verges.
If road verges were integrated into such [ecological corridor design] projects, they might play an important future role in increasing connectivity between natural and semi-natural habitats, particularly across otherwise habitat-poor, human-dominated landscapes where roads often occur [Phillips 2019: 495].
Road verges designed to maximize ecological value thus have an important role to play in the health and wellbeing of urban residents.
Connectivity conservation pays attention to landscape connectivity to support animal species’ movements, keep ecological processes intact, and promote biodiversity. While the strategy of conserving connected, non-fragmented areas and respecting animals’ movement patterns is sound, in practice these plans are usually designed around a single species and its needs.
Brennan et al. looked at the limitations of a single-species focus, and evaluated the movement patterns of multiple species. They created connectivity maps for six large mammal species in the Kavango-Zambezi (KAZA) transfrontier conservation area straddling Angola, Zambia, Zimbabwe, Botswana, and Namibia, and assessed how each individual species’ connectivity maps correlated with that of the others.
This then allowed the authors to identify good ‘surrogate species for connectivity’ – that is, species whose connectivity maps were good representations of other species’ movements through the same area. They also took a look at different types of barriers to animal movements and determined that fences were the greatest obstacle to movement, while roads, rivers, and human-settled areas also deterred movement. Finally, they identified connectivity hotspots on the landscape, which are like bottlenecks through which multiple species pass due to barriers elsewhere. These connectivity hotspots are thus essential places to focus conservation efforts.
The researchers found the hyena and African wild dog to be the most apt surrogate species for connectivity, in spite of a popular practice of using elephants to determine the geographic targets of conservation efforts.
In our examination of connectivity across the landscape, female elephants were found to be only weakly correlated with the five other species in our study. Spotted hyena and African wild dog, in contrast, were strongly correlated with the greatest number of species. They also appeared to be complementary surrogates (i.e. they were correlated with different species), in which case combining their connectivity models could further extend the relevancy of connectivity conservation plans to other species. Thus, as both species are also charismatic, wide-ranging species of conservation concern, they may represent good umbrella species for connectivity in the KAZA region [Brennan 2020: 1707].
They went on to say that “while elephants may not be good surrogate species for connectivity across entire landscapes, they may still be effective as a surrogate at local scales where they can help protect local movement pathways or stepping-stone habitats for other species” [Brennan 2020: 1707].
Their conclusion is not that we should stop paying attention to elephants, which serve important ecological functions and are an iconic and culturally significant animal. Rather, we should look for gaps that may arise if we only conserve areas based on elephant movements, and put these techniques of comparing and combining different species’ movement patterns to use. Noting that animal movements and ecological dynamics play out at different scales, from entire landscapes and transnational parks to smaller corridors, they emphasized the importance of looking at connectivity for multiple species at multiple scales. They urged researchers and policy makers to take a more holistic multi-species approach to connectivity conservation.
Salvaging bycatch data for conservation: unexpected benefits of restored grasslands to amphibians in wetland buffer zones and ecological corridors, Mester et al. 2020
This study considers the effect of grassland restoration on amphibian populations in a 760-acre nature reserve – the Egyek-Pusztakócs Marsh System (EPMS) – established on former farmland in Hungary. The study shows that grassland restoration increased habitat range and quality for amphibians, extended hydrological supply, and limited genetic erosion among previously isolated populations. It also illustrates the role of smaller-scale ecological corridors.
Grassland restoration … creates corridors that maintain connectivity among the amphibian (sub)populations in the EPMS but it may also increase the permeability of the landscape to establish and maintain connections to other nearby metapopulations. Grassland restoration can thus also have an effect of minimizing genetic erosion of populations induced by isolation, which is one of the major causes of global amphibian decline [Mester 2020: 7].
Restoration can benefit amphibians by increasing the area of grasslands available for a variety of life activities such as foraging, burrowing, dispersal/ migration, or hiding from predators, aestivation and hibernation in the non-breeding period and by ensuring functional connectivity between wetlands both in the breeding and non-breeding periods [Mester 2020: 9].
Habitat fragmentation, a frequent consequence of habitat loss, is a primary threat to populations and species because isolated subpopulations are expected to experience reduced population viability and ultimately greater risk of extinction. Colonization and gene flow between habitat patches, however, can mitigate these effects [Gilbert-Norton 2010: 661].
This meta-analysis, consisting of 78 experiments from 35 studies, asked the question: Do ecological corridors increase movement between habitat patches, and how does that differ among taxa? The study’s results answer the first part of the research question affirmatively: “There was approximately 50% more movement between habitat patches connected by a corridor than between isolated habitat patches” [Gilbert-Norton 2010: 665].
Furthermore, corridors increase movement for all taxa. “Most corridors are created for terrestrial vertebrates, including birds, although our data suggest that invertebrates and plants also benefit from corridors” [Gilbert-Norton 2010: 665]. This study found that corridors work equally well for all taxa except birds, for whom the corridors were used less; however, birds still favored corridors compared to surrounding matrix.
While three quarters of the experiments showed corridors to be more effective for movement compared to the matrix landscape, 23% of experiments showed corridors were less effective. The authors suggest several explanations for this result. It’s possible that the “matrix habitat has been misclassified as nonhabitat for a study organism” [Gilbert-Norton 2010: 665], that the habitat quality of the corridor is not particularly high, or that the corridor is difficult to locate, given its small size compared to surrounding landscape. Furthermore, use of corridors varies by species.
That almost a quarter of the studies showed organisms used matrix habitat rather than corridors to move between habitat patches furthers the idea that although corridors may be used by many species, they are unlikely to be used by all species, and whether corridors are relevant for land managers may depend on the species of interest [Gilbert-Norton 2010: 665].
The authors also observed that organisms showed greater use of natural corridors (those existing prior to the study) compared to those created and maintained for the study. The real-world applicability of this, as the authors note, is that “it may be better to protect natural landscape features that function as corridors rather than attempting to create corridors” [Gilbert-Norton 2010: 667]. This highlights the importance of protecting natural or semi-natural lands from development.
In contrast to the Gao et al. [2017] article (above), this study maps out an ecological network spanning the entire nation of China. Most such ecological corridor analysis has previously focused at the local and regional levels, according to the authors. They note that in addition to protecting biodiversity, ecological corridors (ECs) purify air, regulate climate, and “realize the movement of material, energy, and information in the ecosystem” [Liang 2018: 23].
This study identifies a couple of dozen high priority areas for conservation based on the existing diversity and quality of the landscape. These high priority areas encompassed seven ecotones (broadleaf forest, coniferous forest, shrub, herbaceous plant, sparse vegetation, wetland, water body), while built up areas such as cities were low priorities. The authors mapped these conservation priority zones against existing formally protected areas (which cover 15% of the country), finding only 19% overlap and, thus, revealing extensive conservation gaps.
The majority of China’s nature reserves were established without a clear planning framework, and couldn’t maximize efficiency of conservation targets. … important zones for species migration are not considered as conservation goals in the current nature reserve system [Liang 2018: 26].
The ecological corridors were identified by examining the pathways with the least amount of potential resistance (such as built infrastructure) to animals moving along them. The shortest routes were not necessarily chosen given the need to bypass urban areas. The map created through this study offers useful information for national conservation planning.
From a long-term conservation perspective, in view of the rapid habitat loss and biodiversity reduction, the ecological network represents a valuable tool to protect the biotope[5] and their ecological functions in China. In this regard, our results show the importance and need to develop a national protection network maintaining connectivity among them in order to achieve high cost efficiency [Liang 2018: 27].
Changzhou is a city near the Yangtze River delta on the east coast of China that has undergone extensive urban development. “From 2006 to 2014, the built-up area in the city increased by 25.68%” [Gao 2017: 2]. This study is part of an effort to boost biodiversity and ecosystem services in the city, which, at the time of the study, had a few protected patches but no corridors connecting them.
The authors identified potential corridors by comparing three different methods for assessing the level of resistance wildlife would face in moving across the landscape from one habitat patch to another. Corridors were identified by mapping out the paths of least resistance. Potential corridors consisted mainly of riparian greenspaces, followed by forest and farmland, and included between 3.45% and 16% built-up space, depending on the method used. Corridor width was assumed to be 30m. Connection of the most important protected patches should be prioritized in corridor construction.
The International Union for Conservation of Nature (IUCN), which created these guidelines, is an international environmental network founded in 1948 that provides conservation data, assessment and analysis to governments, NGOs and private entities. IUCN also manages the Red List of Threatened Species. This connectivity guideline is part of a series of best practices for protected area land managers.
Providing a definition and context for the importance of connectivity, the authors state:
‘Ecological connectivity’ is the unimpeded movement of species and the flow of natural processes that sustain life on Earth. This is not an overstatement. Without connectivity, ecosystems cannot function properly, and without well-functioning ecosystems, biodiversity and other fundamentals of life are at risk [Hilty 2020: xii].
Moreover,
Most global, regional and national targets for biodiversity conservation, climate change and environmental sustainability cannot be met unless ecological connectivity conservation is addressed [Hilty 2020: 48].
In short, ecological connectivity undergirds the conditions for life on Earth. The authors explain that the concept of connectivity reflects an evolution in conservation science. Previously, nature conservation consisted primarily of setting aside areas of undisturbed or minimally disturbed land. While protected areas remain the foundation of nature conservation, “they are no longer considered sufficient in many places. It is now understood that active measures must also be taken to maintain, enhance or restore ecological connectivity among and between protected areas and OECMs[4] [Hilty 2020: 2].”
Hence,
These Guidelines have been drafted to help clarify and standardize a shift in conservation practice from a narrow focus on individual protected areas to considering them as essential parts of large landscape conservation networks. This is done through creating ‘ecological networks for conservation’ that are specifically designed, implemented and managed to ensure that ecological connectivity is maintained and enhanced where it is present, or restored where it has been lost. Unless systems of protected areas and OECMs retain all essential ecosystem processes, they are not sufficient [Hilty 2020: 3].
The guidelines emphasize the importance of clearly defining one or more ecological objectives for establishing a corridor, such as to facilitate gene dispersal, migration, or adaptation to climate change for particular or multiple species. Clearly defined objectives allow for a corridor to be created in a way that leads to successful outcomes vis a vis the objectives. Primary objectives should relate directly to ecological connectivity, while complementary social or economic objectives (ecosystem services, such as flood and erosion control, enhancing crop pollination, for example) may also be included.
The toolbox for connectivity conservation includes various types of formal and informal recognition, national legislation, local and regional zoning regulations, conservation easements, conservancy design and transportation planning [Hilty 2020: 48].
The importance of connectivity is increasingly recognized in international treaties, and in national and sub-national planning and policy initiatives.
Until recently, connectivity legislation was rare at the national or even sub-national level. Now, countries such as Bhutan, Costa Rica and Tanzania, and sub-national jurisdictions such as California and New Mexico (USA), have enacted corridor legislation. Additionally, site-specific legislation has been enacted in some countries. For example, the South Korea Act on the Protection of the Baekdu Daegan Mountain System, 2003 (Act no. 7038), which came into effect in 2005, designates an area of 263,427 ha. Of this, 86% is made up of 183 existing protected areas and 14% consists of new buffer and core areas that create a biodiversity corridor along the main mountain range of the Korean Peninsula [Hilty 2020: 45].
However, mostly countries have not yet effectively integrated connectivity into policy and planning. Partly this is due to the complexity of establishing ecological corridors.
Connectivity conservation requires innovative implementation approaches to conserve lands and water within the conservation matrix – across patterns of resource use, jurisdictions, cultures and geographies [Hilty 2020: 48].
These guidelines are meant as a toolbox to help local, regional, national and international entities navigate that complexity.
‘Ecological connectivity’ is the unimpeded movement of species and the flow of natural processes that sustain life on Earth. This is not an overstatement. Without connectivity, ecosystems cannot function properly, and without well-functioning ecosystems, biodiversity and other fundamentals of life are at risk [Hilty 2020: xii].
Currently, 15.1% of land on Earth is conservation protected. This article maps out an additional 35.3% of land needing near-term protection, along with ecological corridor routes connecting these areas. Half of the planet’s land is needed to serve as a Global Safety Net to biodiversity loss and stabilize the global climate.
While the parallel crises of biodiversity loss and climate change have generally been approached separately, a key solution for two of the most pressing challenges of our time is the same: conserve enough nature and in the right places [Dinerstein 2020: 1].
The “right places” were identified by mapping areas with rare or endangered species, biodiversity hotspots[3], and places with distinct species assemblages. Onto this, the authors mapped areas where wild large mammals are still able to range widely and freely, a phenomenon that has become rare globally given the extent of anthropogenic land conversion, and areas of remaining intact wilderness.
The study also maps out a system of wildlife corridors to connect conservation areas. Only half of currently protected areas are connected. “Connecting all current terrestrial protected areas via potential wildlife and climate corridors (using 2.5 km as an average corridor width) adds 5,705,206 km2 or 4.3% of the terrestrial realm” [Dinerstein 2020: 4]. Assuming the additional lands identified in this study for conservation are formally protected, the amount of land needed for connectivity would be significantly reduced.
While large conservation protections require national leadership to achieve, the need to establish connectivity presents a role for local and regional actors to restore degraded lands in their midst.
The connectivity analysis offers a template to build from and engage local and regional entities in designing programs centered on restoring connectivity. This effort could merge with global habitat restoration and native tree-planting initiatives now under way [Dinerstein 2020: 7].
Focusing restoration efforts on degraded lands that can serve as wildlife corridors could help achieve other objectives, such as the Bonn Challenge. Similarly, massive tree-planting programs, if designed using native species and planted to restore corridors, riparian and coastal vegetation, and upper watersheds, could contribute to stabilizing climate and restoring connectivity [Dinerstein 2020: 7].
At the national level, countries could use the Global Safety Net framework to map out their own corresponding national safety nets. The 20 countries with the greatest role to play in establishing the Global Safety Net include: Russia, Brazil, Indonesia, the United States, Costa Rica, Peru, and Namibia.
Investments needed for the establishment and management of additional protected areas and restoration of degraded lands, while substantial, are small compared with enormous fossil fuel subsidies. The estimated $4.7 trillion per year in fossil fuel subsidies are expected to decline as the Paris Climate Agreement is implemented, making government resources available for restoring, rather than destroying, our global climate system [Dinerstein 2020: 7].
The authors emphasize that the conservation goals of the Global Safety Net are achievable, especially if indigenous people’s land rights are honored. One third of land identified for a Global Safety Net is managed by indigenous communities in a way that preserves biodiversity and regulates Earth’s atmosphere.
A “Global Safety Net” to reverse biodiversity loss and stabilize Earth’s climate, Dinerstein et al. 2020
Currently, 15.1% of land on Earth is conservation protected. This article maps out an additional 35.3% of land needing near-term protection, along with ecological corridor routes connecting these areas. Half of the planet’s land is needed to serve as a Global Safety Net to biodiversity loss and stabilize the global climate.
While the parallel crises of biodiversity loss and climate change have generally been approached separately, a key solution for two of the most pressing challenges of our time is the same: conserve enough nature and in the right places [Dinerstein 2020: 1].
The “right places” were identified by mapping areas with rare or endangered species, biodiversity hotspots[3], and places with distinct species assemblages. Onto this, the authors mapped areas where wild large mammals are still able to range widely and freely, a phenomenon that has become rare globally given the extent of anthropogenic land conversion, and areas of remaining intact wilderness.
The study also maps out a system of wildlife corridors to connect conservation areas. Only half of currently protected areas are connected. “Connecting all current terrestrial protected areas via potential wildlife and climate corridors (using 2.5 km as an average corridor width) adds 5,705,206 km2 or 4.3% of the terrestrial realm” [Dinerstein 2020: 4]. Assuming the additional lands identified in this study for conservation are formally protected, the amount of land needed for connectivity would be significantly reduced.
While large conservation protections require national leadership to achieve, the need to establish connectivity presents a role for local and regional actors to restore degraded lands in their midst.
The connectivity analysis offers a template to build from and engage local and regional entities in designing programs centered on restoring connectivity. This effort could merge with global habitat restoration and native tree-planting initiatives now under way [Dinerstein 2020: 7].
Focusing restoration efforts on degraded lands that can serve as wildlife corridors could help achieve other objectives, such as the Bonn Challenge. Similarly, massive tree-planting programs, if designed using native species and planted to restore corridors, riparian and coastal vegetation, and upper watersheds, could contribute to stabilizing climate and restoring connectivity [Dinerstein 2020: 7].
At the national level, countries could use the Global Safety Net framework to map out their own corresponding national safety nets. The 20 countries with the greatest role to play in establishing the Global Safety Net include: Russia, Brazil, Indonesia, the United States, Costa Rica, Peru, and Namibia.
Investments needed for the establishment and management of additional protected areas and restoration of degraded lands, while substantial, are small compared with enormous fossil fuel subsidies. The estimated $4.7 trillion per year in fossil fuel subsidies are expected to decline as the Paris Climate Agreement is implemented, making government resources available for restoring, rather than destroying, our global climate system [Dinerstein 2020: 7].
The authors emphasize that the conservation goals of the Global Safety Net are achievable, especially if indigenous people’s land rights are honored. One third of land identified for a Global Safety Net is managed by indigenous communities in a way that preserves biodiversity and regulates Earth’s atmosphere.
Guidelines for conserving connectivity through ecological networks and corridors, Hilty et al. 2020
The International Union for Conservation of Nature (IUCN), which created these guidelines, is an international environmental network founded in 1948 that provides conservation data, assessment and analysis to governments, NGOs and private entities. IUCN also manages the Red List of Threatened Species. This connectivity guideline is part of a series of best practices for protected area land managers.
Providing a definition and context for the importance of connectivity, the authors state:
‘Ecological connectivity’ is the unimpeded movement of species and the flow of natural processes that sustain life on Earth. This is not an overstatement. Without connectivity, ecosystems cannot function properly, and without well-functioning ecosystems, biodiversity and other fundamentals of life are at risk [Hilty 2020: xii].
Moreover,
Most global, regional and national targets for biodiversity conservation, climate change and environmental sustainability cannot be met unless ecological connectivity conservation is addressed [Hilty 2020: 48].
In short, ecological connectivity undergirds the conditions for life on Earth. The authors explain that the concept of connectivity reflects an evolution in conservation science. Previously, nature conservation consisted primarily of setting aside areas of undisturbed or minimally disturbed land. While protected areas remain the foundation of nature conservation, “they are no longer considered sufficient in many places. It is now understood that active measures must also be taken to maintain, enhance or restore ecological connectivity among and between protected areas and OECMs[4] [Hilty 2020: 2].”
Hence,
These Guidelines have been drafted to help clarify and standardize a shift in conservation practice from a narrow focus on individual protected areas to considering them as essential parts of large landscape conservation networks. This is done through creating ‘ecological networks for conservation’ that are specifically designed, implemented and managed to ensure that ecological connectivity is maintained and enhanced where it is present, or restored where it has been lost. Unless systems of protected areas and OECMs retain all essential ecosystem processes, they are not sufficient [Hilty 2020: 3].
The guidelines emphasize the importance of clearly defining one or more ecological objectives for establishing a corridor, such as to facilitate gene dispersal, migration, or adaptation to climate change for particular or multiple species. Clearly defined objectives allow for a corridor to be created in a way that leads to successful outcomes vis a vis the objectives. Primary objectives should relate directly to ecological connectivity, while complementary social or economic objectives (ecosystem services, such as flood and erosion control, enhancing crop pollination, for example) may also be included.
The toolbox for connectivity conservation includes various types of formal and informal recognition, national legislation, local and regional zoning regulations, conservation easements, conservancy design and transportation planning [Hilty 2020: 48].
The importance of connectivity is increasingly recognized in international treaties, and in national and sub-national planning and policy initiatives.
Until recently, connectivity legislation was rare at the national or even sub-national level. Now, countries such as Bhutan, Costa Rica and Tanzania, and sub-national jurisdictions such as California and New Mexico (USA), have enacted corridor legislation. Additionally, site-specific legislation has been enacted in some countries. For example, the South Korea Act on the Protection of the Baekdu Daegan Mountain System, 2003 (Act no. 7038), which came into effect in 2005, designates an area of 263,427 ha. Of this, 86% is made up of 183 existing protected areas and 14% consists of new buffer and core areas that create a biodiversity corridor along the main mountain range of the Korean Peninsula [Hilty 2020: 45].
However, mostly countries have not yet effectively integrated connectivity into policy and planning. Partly this is due to the complexity of establishing ecological corridors.
Connectivity conservation requires innovative implementation approaches to conserve lands and water within the conservation matrix – across patterns of resource use, jurisdictions, cultures and geographies [Hilty 2020: 48].
These guidelines are meant as a toolbox to help local, regional, national and international entities navigate that complexity.
‘Ecological connectivity’ is the unimpeded movement of species and the flow of natural processes that sustain life on Earth. This is not an overstatement. Without connectivity, ecosystems cannot function properly, and without well-functioning ecosystems, biodiversity and other fundamentals of life are at risk [Hilty 2020: xii].
Constructing ecological networks based on habitat quality assessment: a case study of Changzhou, China, Gao et al. 2017
Changzhou is a city near the Yangtze River delta on the east coast of China that has undergone extensive urban development. “From 2006 to 2014, the built-up area in the city increased by 25.68%” [Gao 2017: 2]. This study is part of an effort to boost biodiversity and ecosystem services in the city, which, at the time of the study, had a few protected patches but no corridors connecting them.
The authors identified potential corridors by comparing three different methods for assessing the level of resistance wildlife would face in moving across the landscape from one habitat patch to another. Corridors were identified by mapping out the paths of least resistance. Potential corridors consisted mainly of riparian greenspaces, followed by forest and farmland, and included between 3.45% and 16% built-up space, depending on the method used. Corridor width was assumed to be 30m. Connection of the most important protected patches should be prioritized in corridor construction.
Integrating priority areas and ecological corridors into national network for conservation planning in China, Liang et al. 2018
In contrast to the Gao et al. [2017] article (above), this study maps out an ecological network spanning the entire nation of China. Most such ecological corridor analysis has previously focused at the local and regional levels, according to the authors. They note that in addition to protecting biodiversity, ecological corridors (ECs) purify air, regulate climate, and “realize the movement of material, energy, and information in the ecosystem” [Liang 2018: 23].
This study identifies a couple of dozen high priority areas for conservation based on the existing diversity and quality of the landscape. These high priority areas encompassed seven ecotones (broadleaf forest, coniferous forest, shrub, herbaceous plant, sparse vegetation, wetland, water body), while built up areas such as cities were low priorities. The authors mapped these conservation priority zones against existing formally protected areas (which cover 15% of the country), finding only 19% overlap and, thus, revealing extensive conservation gaps.
The majority of China’s nature reserves were established without a clear planning framework, and couldn’t maximize efficiency of conservation targets. … important zones for species migration are not considered as conservation goals in the current nature reserve system [Liang 2018: 26].
The ecological corridors were identified by examining the pathways with the least amount of potential resistance (such as built infrastructure) to animals moving along them. The shortest routes were not necessarily chosen given the need to bypass urban areas. The map created through this study offers useful information for national conservation planning.
From a long-term conservation perspective, in view of the rapid habitat loss and biodiversity reduction, the ecological network represents a valuable tool to protect the biotope[5] and their ecological functions in China. In this regard, our results show the importance and need to develop a national protection network maintaining connectivity among them in order to achieve high cost efficiency [Liang 2018: 27].
A meta-analytic review of corridor effectiveness, Gilbert-Norton et al. 2010
Habitat fragmentation, a frequent consequence of habitat loss, is a primary threat to populations and species because isolated subpopulations are expected to experience reduced population viability and ultimately greater risk of extinction. Colonization and gene flow between habitat patches, however, can mitigate these effects [Gilbert-Norton 2010: 661].
This meta-analysis, consisting of 78 experiments from 35 studies, asked the question: Do ecological corridors increase movement between habitat patches, and how does that differ among taxa? The study’s results answer the first part of the research question affirmatively: “There was approximately 50% more movement between habitat patches connected by a corridor than between isolated habitat patches” [Gilbert-Norton 2010: 665].
Furthermore, corridors increase movement for all taxa. “Most corridors are created for terrestrial vertebrates, including birds, although our data suggest that invertebrates and plants also benefit from corridors” [Gilbert-Norton 2010: 665]. This study found that corridors work equally well for all taxa except birds, for whom the corridors were used less; however, birds still favored corridors compared to surrounding matrix.
While three quarters of the experiments showed corridors to be more effective for movement compared to the matrix landscape, 23% of experiments showed corridors were less effective. The authors suggest several explanations for this result. It’s possible that the “matrix habitat has been misclassified as nonhabitat for a study organism” [Gilbert-Norton 2010: 665], that the habitat quality of the corridor is not particularly high, or that the corridor is difficult to locate, given its small size compared to surrounding landscape. Furthermore, use of corridors varies by species.
That almost a quarter of the studies showed organisms used matrix habitat rather than corridors to move between habitat patches furthers the idea that although corridors may be used by many species, they are unlikely to be used by all species, and whether corridors are relevant for land managers may depend on the species of interest [Gilbert-Norton 2010: 665].
The authors also observed that organisms showed greater use of natural corridors (those existing prior to the study) compared to those created and maintained for the study. The real-world applicability of this, as the authors note, is that “it may be better to protect natural landscape features that function as corridors rather than attempting to create corridors” [Gilbert-Norton 2010: 667]. This highlights the importance of protecting natural or semi-natural lands from development.
Characterizing multispecies connectivity across a transfrontier conservation landscape, Brennan et al. 2020
Connectivity conservation pays attention to landscape connectivity to support animal species’ movements, keep ecological processes intact, and promote biodiversity. While the strategy of conserving connected, non-fragmented areas and respecting animals’ movement patterns is sound, in practice these plans are usually designed around a single species and its needs.
Brennan et al. looked at the limitations of a single-species focus, and evaluated the movement patterns of multiple species. They created connectivity maps for six large mammal species in the Kavango-Zambezi (KAZA) transfrontier conservation area straddling Angola, Zambia, Zimbabwe, Botswana, and Namibia, and assessed how each individual species’ connectivity maps correlated with that of the others.
This then allowed the authors to identify good ‘surrogate species for connectivity’ – that is, species whose connectivity maps were good representations of other species’ movements through the same area. They also took a look at different types of barriers to animal movements and determined that fences were the greatest obstacle to movement, while roads, rivers, and human-settled areas also deterred movement. Finally, they identified connectivity hotspots on the landscape, which are like bottlenecks through which multiple species pass due to barriers elsewhere. These connectivity hotspots are thus essential places to focus conservation efforts.
The researchers found the hyena and African wild dog to be the most apt surrogate species for connectivity, in spite of a popular practice of using elephants to determine the geographic targets of conservation efforts.
In our examination of connectivity across the landscape, female elephants were found to be only weakly correlated with the five other species in our study. Spotted hyena and African wild dog, in contrast, were strongly correlated with the greatest number of species. They also appeared to be complementary surrogates (i.e. they were correlated with different species), in which case combining their connectivity models could further extend the relevancy of connectivity conservation plans to other species. Thus, as both species are also charismatic, wide-ranging species of conservation concern, they may represent good umbrella species for connectivity in the KAZA region [Brennan 2020: 1707].
They went on to say that “while elephants may not be good surrogate species for connectivity across entire landscapes, they may still be effective as a surrogate at local scales where they can help protect local movement pathways or stepping-stone habitats for other species” [Brennan 2020: 1707].
Their conclusion is not that we should stop paying attention to elephants, which serve important ecological functions and are an iconic and culturally significant animal. Rather, we should look for gaps that may arise if we only conserve areas based on elephant movements, and put these techniques of comparing and combining different species’ movement patterns to use. Noting that animal movements and ecological dynamics play out at different scales, from entire landscapes and transnational parks to smaller corridors, they emphasized the importance of looking at connectivity for multiple species at multiple scales. They urged researchers and policy makers to take a more holistic multi-species approach to connectivity conservation.
Salvaging bycatch data for conservation: unexpected benefits of restored grasslands to amphibians in wetland buffer zones and ecological corridors, Mester et al. 2020
This study considers the effect of grassland restoration on amphibian populations in a 760-acre nature reserve – the Egyek-Pusztakócs Marsh System (EPMS) – established on former farmland in Hungary. The study shows that grassland restoration increased habitat range and quality for amphibians, extended hydrological supply, and limited genetic erosion among previously isolated populations. It also illustrates the role of smaller-scale ecological corridors.
Grassland restoration … creates corridors that maintain connectivity among the amphibian (sub)populations in the EPMS but it may also increase the permeability of the landscape to establish and maintain connections to other nearby metapopulations. Grassland restoration can thus also have an effect of minimizing genetic erosion of populations induced by isolation, which is one of the major causes of global amphibian decline [Mester 2020: 7].
Restoration can benefit amphibians by increasing the area of grasslands available for a variety of life activities such as foraging, burrowing, dispersal/ migration, or hiding from predators, aestivation and hibernation in the non-breeding period and by ensuring functional connectivity between wetlands both in the breeding and non-breeding periods [Mester 2020: 9].
Ecosystem service provision by road verges, Phillips et al. 2019
‘Road verges’ are strips of land on either side of roads and highways that are on average 3-4m wide, but can be as narrow as a few centimeters or many meters wide. “Road verges are commonly grassland habitats, but can be shrubland, forest or artificial arrangements of trees and horticultural plants, and we use the term also to include bare earth and freshwater bodies (e.g. ditches)” [Phillips 2019: 489]. They can also be barren ground or ditch. Not all road verges are managed; when management does occur, it is typically geared toward safety – clearing vegetation to enhance visibility.
There is currently an estimated 36 million linear kilometers of road network in the world, the length of which is expected to increase by 70% by 2050; thus, the total area of road verges will increase as well. “Road and road verge construction will displace habitats and cause many negative ecological and social impacts” [Phillips 2019: 494]. However, there is potential to mitigate that impact by maximizing the ecological value of road verges. Currently, “there may well be 270,000 km2 of road verge globally (0.2% of land), which is similar to the total area of the United Kingdom” [Phillips 2019: 492], with this surface area expected to grow.
While roads run like a network of veins across landscapes, causing widespread negative ecological impacts to adjacent areas, road verges form a parallel network and have the potential both partially to mitigate negative impacts of roads and to deliver environmental benefits [Phillips 2019: 490].
Where roads cut through natural habitat, the road verges will represent a net loss of biodiversity. By contrast, verges can increase biodiversity in highly degraded environments such as cities or industrial farmland. Furthermore, because of the growing urban population, the importance of natural and semi-natural environments will be increasingly important. Road verges designed to maximize ecological value thus have an important role to play in the health and wellbeing of urban residents.
Road verges might increase connectivity in highly modified urban and agricultural landscapes if road verges of suitable size, habitat quality and continuity are created alongside roads, at least for species that are highly mobile or able to persist in narrow, linear habitats [Phillips 2019: 495].
While roads often act as barriers to wildlife and ecological connectivity, ecological corridor design could benefit by taking into account the potential benefits of road verges.
If road verges were integrated into such [ecological corridor design] projects, they might play an important future role in increasing connectivity between natural and semi-natural habitats, particularly across otherwise habitat-poor, human-dominated landscapes where roads often occur [Phillips 2019: 495].
Road verges designed to maximize ecological value thus have an important role to play in the health and wellbeing of urban residents.
Fence ecology: frameworks for understanding the ecological effects of fences, McInturff et al. 2020
Conceptually the inverse of wildlife corridors, fences aim to disconnect. They are built to separate people across national borders, livestock from predators, to delineate property lines, and even to protect wildlife conservation reserves. Globally, fences are ubiquitous, more prevalent even than roads, and proliferating. Yet their ecological impact is relatively unstudied.
Fences are often framed as a management tool rather than a globally significant ecological feature, and they are a notable omission from efforts to map global infrastructure, including the human footprint [McInturff 2020: 971].
This analysis reviews 446 studies starting from 1948 on various types of fencing to assess impacts; however, most of the studies focus on the effect of fencing on particular species (specifically, those the fencing is meant to protect), rather than on multiple species, communities or ecosystems.
Conservation and restoration fences, for example, have support within the literature for their beneficial effects for wildlife and sensitive plant species for which they are built, making such species “winners” in the fencing game. On the other hand, there is a critical lack of information on species that are not the targets for which fences are built, and our review shows that only 10.8% (48 of 446) of studies focus on nontarget species [McInturff 2020: 975].
While fences aiming to protect particular species usually achieve that goal, they inevitably hurt other species.
… often the clearest winners because of fencing are the species that humans value most, whereas losers are inevitable but may remain invisible [McInturff 2020: 975].
Broadly speaking, fences favor generalists and disturbance specialists, many of which are invasive, as well as small and small-ranged, nonmigratory species. Fences therefore heavily restrict what makes a species a winner [McInturff 2020: 975].
The deleterious effects of fences include: impeding migration, reducing gene flow between populations, restructuring community composition and obstructing interspecies interactions, such as between predators and prey. These community-level changes can have ripple effects in the ecosystem. For example, livestock fences effectively excluding dingoes in Australia led to this large predator’s eradication. “Without dingoes, researchers have tracked a continental-scale mesopredator [mid-level predator] release that has altered biodiversity and habitats over enormous areas of Australia” [McInturff 2020: 979].
While fences limit certain interspecies interactions, they concentrate others:
Even where conservation or restoration fences seemingly protect whole habitats, research still points to differential outcomes for constituent species. In addition, pathogens and parasites may spread more rapidly where species interactions are concentrated within reserves. In central Kenya, for example, smaller fenced reserves produced higher gastrointestinal parasite infection rates among impala [McInturff 2020: 977].
The authors recommend a greater research focus on the cumulative ecological effects of fencing, policy that limits fence building and encourages fence removal or fence design that is more wildlife-friendly. They caution that fencing is among the major drivers of anthropogenic change.
As fencing continues to rapidly proliferate, there is potential for a dangerous future in which fences simultaneously alter ecological processes at multiple scales, likely producing more losers than winners, and potentially resulting in ecosystem state shift or collapse [McInturff 2020: 977].
Livestock fences effectively excluding dingoes in Australia led to this large predator’s eradication. “Without dingoes, researchers have tracked a continental-scale mesopredator [a mid-level predator] release that has altered biodiversity and habitats over enormous areas of Australia” [McInturff 2020: 979].
European Context
Status of the Natura 2000 network (from State of Nature in the EU report), EEA (European Environmental Agency) 2020
While not an ecological corridor per se, the Natura 2000 network is the largest coordinated network of conservation areas in the world. Covering 17.9% of Europe’s land area and nearly 10% of the continent’s marine areas, the network includes 27,852 sites with an area of 1,358,125 km2. The terrestrial portion of the Natura 2000 network is mostly covered by forests and transitional shrublands. It also includes grasslands and wetlands, as well as pastures, cropland and a small amount of artificial surface (developed/built land).
Member States need to ensure that sufficient protection and appropriate measures are implemented in Natura 2000 sites for habitats and species of community interest and that they form a functional network [EEA 2020: 109].
However, the sites are not strictly protected by virtue of being part of the network. In fact, the sites include a variety of land uses.
Within the network, arable land and permanent crops have increased, while grasslands and forests have decreased. … Pastures and mosaic farmland (with approximately 18 %) and inland wetlands and water bodies (with approximately 10 %) have been extensively transformed into arable land and permanent crops both inside and outside the network. Recent research has shown, however, that high nature value (HNV) farmland inside Natura 2000 sites is less likely to be converted into artificial surfaces than such farmland outside the network and is more likely to maintain its pattern of mosaic farming [EEA 2020: 113].
This assessment of the network’s effectiveness found that “species and habitats are more likely to have a good conservation status if they are well covered by the Natura 2000 network” [EEA 2020: 121]. However, limited monitoring inside and outside the network prevents a more detailed analysis of Natura 2000’s effectiveness. Furthermore, due to a limited implementation of conservation measures, the network’s potential has not yet been fully “unlocked,” according to the report.
To improve Natura 2000’s potential, the authors recommend, among other measures, improving connectivity between protected areas. Noting that sites chosen for inclusion in the network are often motivated by economic rather than ecological interests.
Incoherent planning and site selection approaches between and within Member States has led to insufficient functional connectivity and spatial connectedness between neighboring countries and habitats and gaps in coherence within Member States. This highlights the need to increase connections between protected areas and the level of protection beyond the site [EEA 2020: 122].
Also recommended is increasing stakeholder participation, such as through citizen science monitoring initiatives, and better integration of biodiversity protections into other policy domains.
The resulting low awareness of the diverse benefits produced by the Natura 2000 network is often compounded by a long-standing conflict between economic or political interests and conservation goals. There is thus an urgent need to increase coherence between biodiversity policy and other policy areas, such as in the fields of agriculture and economic and rural development, and create a more integrated approach to address potential conflicts and trade-offs between various interests while fostering synergies [EEA 2020: 124].
The report’s summary conclusion recommends increasing marine and terrestrial conservation areas in the Natura 2000 network to 30% each, strictly protecting these areas, and improving connectivity among them.
Blue and green corridors [Les trames vertes et bleues] in France, Ministry of Ecological Transition 2017
Spurred to action by the European Union and a vision for a pan-European ecological network, France encoded the idea of the “trames vertes et bleues” into law in 2009. The national government worked with all the regional governments to develop maps showing areas with the highest levels of biodiversity. This includes protected areas, stretches of coastline, riparian zones, woods, and other undeveloped areas, whether public or private. The maps also show ecological corridors – both those in good condition needing to be preserved, and those that are highly degraded and requiring restoration.
The regional maps are meant to be integrated into urban planning at the level of city and county (department). Rather than being a regulatory tool, the maps are an information source allowing urban development to proceed in a way that limits impact on biodiversity. The ecological corridor initiative is designed as an invitation and encouragement to local governments, organizations, businesses and individuals to collaborate and to act in favor of biodiversity.
The preservation and restoration of ecosystem connectivity entails acting everywhere possible: in rural environments, in aquatic ecosystems and in urban areas [MTES 2017, translation].
Articulating the politics of green and blue infrastructure and the mitigation hierarchy for effective biodiversity preservation in France [Articuler la politique Trame verte et bleue et la séquence Éviter-réduire-compenser: complémentarités et limites pour une préservation efficace de la biodiversité en France], Chaurand & Bigard 2019
This article reviews the historical development of two pieces of environmental legislation in France – the use of the “mitigation hierarchy” to assess and limit environmental impact in project development and the promotion of ecological corridors. Theoretically, these two laws overlap when urban development projects in proximity to areas of ecological significance use the mitigation hierarchy (avoid, reduce, compensate) to ensure these zones are protected within the scope of the project.
1976: “Protection of Nature” law in France introduced the mitigation hierarchy, aiming to avoid or reduce harm to the environment, or to compensate if harm is unavoidable.
1992: Concept of “biodiversity” entered public discourse internationally, following the Earth Summit in Rio, Brazil.
1996: France ratified European ecological corridor strategy.
1999-2000: Concept of “sustainable development” emerged in France.
2004: National strategy for protecting biodiversity adopted.
2007: “Grenelle de l’Environnement” meeting created the “Trame Verte et Bleue” (TVB) policy (green and blue infrastructure, encompassing ecological corridors)
2016: Biodiversity law enacted, creating national agency and regional committees on biodiversity
In spite of this policy evolution, commitment to ecological corridors has yet to move from a “TVB papier” to a “TVB de projets et d’action.” In other words, much discussion and mapping efforts have not yet resulted in the development of the imagined ecological corridor network. The authors speculate as to why this is so, explaining that the resources and coordination needed for enforcement are lacking. Even though “the creation, preservation and restoration of ecological connectivity” has been integrated into urban planning code, such considerations are often sidelined. Furthermore, definitions are vague: the objective of the TVB is the “good condition” of ecological continuity, but “good condition” is not defined. Lastly, taking action in defense of ecological continuity requires pro-active collaboration among levels of government from local to regional to national.
The authors propose better integration of these to policy tools. For example, the TVB designates certain non-protected areas throughout the country that are ecologically functional and serve a role in the eco-corridor network as key areas to “preserve.” With better communication between this TVB framework, the mitigation hierarchy could be applied at the level of “avoiding” harm to places designated as preservation priorities, but lacking formal “protected” status. In projects where harm is unavoidable, the mitigation hierarchy could be applied at the level of “reducing” harm to maximize the percentage of remaining green space as well as the permeability to wildlife of the built structures (such as passageways through fences). The “compensation” level of the mitigation hierarchy could be applied in the context of regenerating ecosystem function to areas designated in the TVB schema as needing ecosystem restoration.
The authors note that advocates for the TVB are clustered at the national level and within research institutions, while the people responsible for urban planning decisions are local and are not necessarily well versed in the scientific framework for the TVB. Local actors tend to focus on priorities other than ecological continuity. One measure to address this, according to the authors, would be the training of local “relays” to transmit knowledge of ecological principles vis a vis the TVB to local urban planners.
Woods and hedgerows of Brittany countryside [Le bocage Bretagne], OEB (L’Observatoire de l’Environnement en Bretagne) 2018
Produced by a regional consortium on the environment in Brittany, France, this report describes the ecological value of woody strips encircling agricultural fields and enmeshing the countryside, their decline, and ways to incentivize their protection.
Brittany is a heavily agricultural region that also features a long stretch of coastline where urban development and expansion is ongoing. Due to mechanization and enlargement of farm fields, average parcel size has increased since the 1950s, shrinking the extent of woody hedgerow (“bocage”) between fields. Between 1996 and 2008, the total length of hedgerow decreased 12%. This change is concerning because Brittany is already one of the most fragmented and least wooded parts of France.
The report explains the value of the bocage is its provisioning of habitat, connectivity between habitats, biodiversity, erosion control, groundwater recharge, and flood mitigation. Half the population of Brittany lives in areas susceptible to flooding. Furthermore, at least five endangered animal species depend on the habitat created by the bocage. Protection of this woody network is key to remedying both problems, while also providing direct benefits to farmers, such as habitat for pest predators.
The form and shape of the bocage varies throughout the region, but can include grasses, bushes and/or trees, forming one or more layers of vegetation; and heterogenous landscape features such as berms, ditches, logs, and rocks/boulders, which create microhabitats.
The network of hedge and berms, accompanied by fields, ponds and wetlands, constitutes an important natural environment because of its heterogeneity and potential for complex exchanges. It has the particularity of being able to reach a myriad of increasingly isolated natural spaces in the heart of a changing agricultural countryside subject to ongoing urbanization. Similar to a forest edge environment, the richness of the bocage can be explained by the diversity of habitats it adjoins [OEB 2018: 8, translated].
Regulatory and incentive programs payments have sought to encourage farmers to preserve their hedgerows. However, the authors suggest that a stronger economic valuation of these linear woods is needed to protect and expand them. They suggest strategies for stimulating the market for firewood and other products harvested from sustainably managed hedgerow, where biodiversity protection is an explicit aim and co-benefit.
Americas Context
Shaping land use change (LUC) and ecosystem restoration in a water-stressed agricultural landscape to achieve multiple benefits, Bryant et al. 2020
In spite of its obvious benefits, agriculture, which covers one third of the Earth’s land surface, damages biodiversity and ecosystem services. In some regions, land degradation and depletion of water resources from irrigation have been so great that historical levels of food production in these regions risk decline. Some areas of previously productive farmland will likely need to be retired from use. Within this context, maintaining and enhancing natural corridors and promoting semi-natural, multifunctional landscapes can significantly contribute to recovering biodiversity and mitigating air and water pollution.
Using California’s San Joaquin Valley (SJV) as a case study, this paper illustrates a pragmatic approach to incorporating ecological corridors into working landscapes. The authors offer a new analytical approach that simultaneously incorporates resource-constrained (water, in this case) land-use change (LUC) modeling within the planning and optimization process. The goals are to simultaneously:
Meet water-use-reduction policy goals for the area under study within the next two decades
Identify lands for retirement that are (1) likely to be retired anyways and (2) offer high-value habitat for native species and biodiversity.
Over the past century, SJV has been transformed into one of the largest agricultural economies in the world. However, this economic success has been costly to the SJV in several ways, including:
Damaged infrastructure: high rates of groundwater extraction in the SJV have led to groundwater overdraft and unreplenished aquifers, resulting in large-scale land subsidence. Most of the subbasins in the SJV are categorized as critically overdrawn, and some regions have sunk over 8 meters since the early 20th century; this land subsidence further imperils water availability and quality by impacting water storage and delivery infrastructure.
Decreased human health, as a result of impaired air and water quality, leading to chronic health problems
Threats to wildlife and biodiversity; for example, some species have lost up to 98% of their habitat range, and over 35 native species are listed as threatened or endangered
“In response to these challenges, and amid significant drought-driven fallowing, California passed the Sustainable Groundwater Management Act (SGMA), which … obligates locally governed groundwater subbasins to develop plans that will achieve sustainable groundwater use over the next two decades” [Bryant 2020: 2]. To meet these requirements, many subbasins will meet with severe groundwater pumping restrictions. If these areas are not able to coordinate their pumping activities and augment water supplies, the SGMA may require a reduction in cultivation area through fallowing or permanent retirement.
Given the likely retirement of 86,000 ha of irrigated agricultural land, the authors explore spatial optimization of retired land for conservation efforts. They find that a key strategy is the identification of areas that were destined for retirement from cropping which could be shifted to restoration and habitat enhancement, as well as possibly shifting some areas destined for retirement that have “low habitat value” with regards to wildlife for areas with “high habitat value.” Priority restoration areas identified in this analysis include many that are contiguous and located near designated wildlife areas.
Importantly, the analysis presented here is “explicitly organized to help inform engagement between conservation actors and agricultural land managers about how habitat goals can be achieved in ways that benefit communities in the SJV” [Bryant 2020: 3]. The potential positive futures indicated by such analysis can be used to identify opportunities for collaboration between the conservation and agricultural communities, with a goal of guiding land use change toward achieving multiple benefits, such as recovery of imperiled natural communities, resilient agricultural production, and improved public health outcomes.
While it poses a great challenge, the impending transformation in the SJV also presents an opportunity to proactively shape the landscape in ways that not only ensure agricultural and water sustainability, but also achieve many other socio-ecological goals, such as biodiversity protection and improved human health. However, given that achievement of many of these objectives is determined by where things happen on the landscape (rather than simply the aggregate amounts of cultivation, retirement, or restoration), stakeholdersneed a systematic way to integrate these objectives to informmulti-benefit spatial planning [Bryant 2020: 4].
Integrating Agricultural Landscapes with Biodiversity Conservation in the Mesoamerican Hotspot, Harvey et al. 2007
The fate of biodiversity within protected areas is therefore inextricably linked to the broader landscape context, including how the surrounding agricultural matrix is designed and managed [Harvey 2007: 8].
Rather than discussing ecological corridors per se, this article emphasizes the importance of a whole-landscape approach to biodiversity conservation. Pointing out that protected nature reserves are weakened when isolated, these authors focus on the role of the entire surrounding agricultural matrix for restoring connectivity.
In contrast to the prevailing trend of managing protected areas and productive lands separately, we propose integrated landscape management in which conservation and production units within the agricultural matrix are managed jointly for long-term sustainability. We do not advocate agricultural intensification to spare further forest conversion because this approach is unlikely to have the intended outcome, for reasons discussed. Instead, conservation efforts should be based on the recognition that how agriculture is conducted and how different land uses are distributed spatially and temporally determine the region’s biodiversity. Lasting conservation will therefore require alliances among conservation biologists, farmers, and land managers to actively plan the future of Mesoamerican landscapes [Harvey 2007: 9].
The sections of the agricultural matrix the authors prioritize for biodiversity conservation include areas near riparian and other key ecological corridors, and they recommend leveraging support for the Mesoamerican Biological Corridor to spur regional action. Priority conservation areas are also more likely to encompass landscapes with a high diversity of indigenous and traditional cropping systems than those dedicated to industrial agriculture because “the chances of reconciling farming and biodiversity conservation there [agro-industrial systems] are slim” [Harvey 2007: 10].
The authors argue that, in contrast to large-scale, export-oriented industrial production, small-holder and indigenous agricultural systems are more compatible with biodiversity conservation, increased food production and rural income. The authors propose economic and regulatory instruments and greater regional collaboration to enhance native tree cover on farms, promote traditional, ecologically based farming practices, and to protect remaining intact habitat and restore degraded lands. The overarching vision is to accomplish conservation and agricultural production objectives for the region in mutually reinforcing ways.
The fate of biodiversity within protected areas is therefore inextricably linked to the broader landscape context, including how the surrounding agricultural matrix is designed and managed [Harvey 2007: 8].
The concept of green corridor and sustainable development in Costa Rica, Beauvais & Matagne 1999
The concept of sustainable development presumes that human economic systems and overall wellbeing depend on functioning ecosystems. Therefore, ecological rhythms should not be transgressed to the point that they fail to provide the vital services needed today and in future generations.
According to this model, economic development becomes a necessary but insufficient condition for society to progress [Beauvais & Matagne 1999: 6, translated].
Costa Rica holds at least 5% of the world’s species, in spite of making up 0.03% of its land surface. As an isthmus, Costa Rica is influenced by weather patterns from two oceans, as well as a north-south migration route. In addition to this, its mountainous terrain creates a heterogenous mosaic of habitats and niches. However, the country has been severely deforested. Forest covered 66% of land surface in 1940, and only 25% by 1987; the loss of forest led to extreme erosion.
As presented in this article, an ecological corridor consists of at least two protected ecosystem patches that are connected by a protected vegetated strip of at least a few kilometers in width, and the whole area surrounded by a buffer zone. Multiple units of two connected patches could in turn be connected, stretching into a corridor that the whole length of the country. A green Costa Rican corridor could connect to green corridors in adjacent countries, ultimately recreating the entire isthmic corridor that once existed.
However, the tone of this article is not optimistic about conservation, citing several political obstacles to conservation and ecosystem restoration. According to the authors, a combination of neocolonialist pressure, poverty, corruption, and capitalistic interests allow for trees to be cut even in protected areas and prevent the establishment of new protected areas and corridors.
The Mesoamerican Biological Corridor in Panama and Costa Rica, Dettman 2006
At the end of the 1980s, as a period of severe conflict in Central America was winding down, most countries in the isthmus signed the Charter Agreement for the Protection of the Environment, which established a sustainable development commission. At the same time, the “Central American Protected Areas System (SICAP) created approximately 11.5 million hectares of protected areas throughout the region” [Dettman 2006: 18].
This paved the way for international attention and investment in what became the Mesoamerican Biological Corridor (MBC). The original intention was to promote biodiversity and economic development in tandem through investment in local projects. However, in the 2000s, the international coordinators of the MBC shifted the focus from biodiversity protection (although the establishment of ecological corridors remains an objective) to a greater emphasis on economic development. This author explains that the institution’s decision-making process is overly top-down, and would benefit from input from local people who are implementing projects on the ground.
Between Bolivar and Bureaucracy: The Mesoamerican Biological Corridor, Liza Grandia 2007
Written by an anthropologist working in Central American conservation efforts for more than 10 years, this article describes the Mesoamerican Biological Corridor (MBC) project as having succumbed to a neoliberal agenda. Although originally spearheaded by Central American environmentalists, the notion of cross-border environmental collaboration was adopted by the World Bank and large international conservation organizations working in Central America in the 1990s. In the hands of these international giants, the biological corridor initiative became a bureaucratic, top-down project, deaf to the voices of local communities.
With all this new bureaucracy, a broad and unfocused agenda, and the challenges of high-level political coordination, the MBC quickly lost its potential to inject a stronger environmental justice component into regional biodiversity conservation programs.
Indeed, the MBC that emerged from the World Bank’s incubator was decidedly more business-oriented than initial proposals for Central American environmental coordination at the 1992 Earth Summit [Grandia 2007: 486].
In this context, the MBC’s conservation efforts have focused more on securing land for protected parks and less on community-based initiatives. The author suggests that in addition to land protection, the MBC should engage farmers in capacity building for eco-agriculture with a view toward achieving landscape-wide ecological connectivity.
The corridor approach might also draw greater attention to the agrarian contexts outside of parks, which may be just as ecologically important as what happens inside parks. By bringing agricultural systems into conservation debates, corridors may present new opportunities for supporting fair-trade projects and other small-scale agroforestry systems compatible with conservation. In other words, corridors could offer a method for moving beyond protectionism to embrace a mosaic vision for conservation that includes local people more explicitly. Corridor planning frameworks also could provide more democratic conservation forums [Grandia 2007: 484].
Effectiveness of Panama as an intercontinental land bridge for large mammals, Meyer et al. 2019
One of the world’s largest corridor projects is the Mesoamerican Biological Corridor (MBC). Initiated in the 1990s, the MBC aims to connect protected areas between southeastern Mexico and Panama [Meyer 2019: 2].
The ecological functionality of the MBC has not been much assessed, in part because direct approaches to measuring connectivity are costly and challenging. In this study, researchers used a simpler, indirect approach to measure forest connectivity through Panama for nine mammals. Using camera traps (cameras that are automatically triggered by a change in some activity in the vicinity, like the presence of an animal), they documented the presence (or absence) of these mammals in 28 forest sites along the Atlantic coast. The corridor was presumed to be functioning for animals whose presence was established across the entire length of the monitored range.
The species monitored in this study are forest specialists, including ungulates, carnivores and an insectivore, all of which are threatened by habitat loss and hunting, some more than others. Of the 43% of land in Panama that is forested, 44% is protected, mostly along the Atlantic coast. Steady economic development threatens remaining ecosystems with investments in large infrastructure projects, real estate, mining, tourism, and energy.
Large mammals are an indicator species for the success of conservation efforts. This is because:
Large mammals are generally at a higher risk of extinction in disturbed landscapes than other taxa because their large home ranges and low population densities at broad spatial scales mean their populations are more likely to be fragmented and because they are heavily hunted [Meyer 2019: 3].
The researchers found that even the four most prevalent species in the study are susceptible to population fragmentation by any further habitat loss.
We found that there was little connectivity for white-lipped peccary [a pig-like animal] and white-tailed deer and that, although 4 of the species (collared peccary, red brocket deer, puma, and ocelot [a wild cat]) occurred in most of the sites, a small decrease in connectivity of 20% would disrupt their continuous distributions across Panama. White-lipped peccary, giant anteater, white-tailed deer, jaguar, and tapir [a pig-like animal with a short trunk] had lower probability of occurring in all the sites and were therefore even more at risk of connectivity loss, as evidenced by >1 connectivity gap. This indicates the MBC may not function for the majority of species, especially considering we did not account for potential effects of hunting, which would make connectivity even more challenging [Meyer 2019: 8].
Citing imminent development projects, such as a new road that will pass through the forested northern coast and associated large hotel projects, the authors predict that ongoing loss of connectivity is likely. Moreover, the deteriorating condition of the corridor in Panama bodes poorly for the MBC overall.
The disruption of connectivity between tropical forests in Central America, and hence the possible separation of mammal populations, is an indicator of the overall functioning of the MBC for wildlife [Meyer 2019: 11].
Belize creates one of Central America’s largest biological corridors, Dasgupta 2018
The Belize government approved a plan in February 2018 to create a 110-square-kilometer biological corridor connecting two nature reserves in the northeast of the country. This outcome resulted from collaboration among NGOs, the government and private property owners. The latter agreed to conserve (to not deforest or otherwise degrade) the parts of their land that would become part of the wildlife corridor. In exchange, the government would not collect taxes on this land. This corridor, which was initiated in the context of the larger Mesoamerican Biological Corridor project, is meant to protect jaguars, cougars and tapirs, among other wildlife.
The woman building the forest corridors saving Brazil’s black lion tamarin, Zanon 2020
“The tamarin is unable to do anything to save its own species. And we, human beings, are the ones who are destroying their environment,” says conservationist Gabriela Rezende. “So, when I got the opportunity to see this animal in the wild, I felt partly responsible for its future.”
Rezende works with the Institute for Ecological Research in the Brazilian state of Sao Paolo to create ecological corridors connecting the forest fragments where the world’s only 1,800 black lion tamarin live in isolated populations. Since 1984, the institute has worked to protect this small primate species, which had reached a low point of 100 individuals and was listed as “critically endangered.” In addition to research and forest restoration, the institute also does environmental education with the local communities. This includes collaboration on nine tree nurseries administered by local people as small businesses that also provide school kids the chance to learn about local forest species that will be planted in corridors.
Leveraging a state policy requiring 20% of privately owned property to be in nature reserves, Rezende worked with landowners to identify patches to be restored that would physically connect forest fragments. Once corridors are complete, the total amount of land in connected habitat will be 111,000 acres. Rezende estimates the black lion tamarin population could increase 30% once it’s able to use the whole forest corridor. The restoration project will benefit other species too, including anteaters, tapirs (a pig-like animal with a short trunk), pumas, and ocelots (another wild cat species).
Establishing ecological corridors is a way to mitigate the effects of habitat loss and fragmentation. Ecological corridors are linear landscape elements connecting otherwise isolated habitat patches within a larger matrix of environmentally degraded lands (urban or agricultural, for example). The corridors facilitate gene dispersal and migration, while also expanding habitat range for species constrained by patch size. They can take the form of riparian (river) ecosystems, hedge networks, forest edges, or grassy bands.
Ecological (wildlife/biological) corridors are based on the concept of connectivity, which is essential to ecosystem functioning, and thus to the persistence of life on earth. When connectivity is disrupted, so are “ecological processes such as gene flow, pollination, [and] wildlife dispersal” [Estreguil 2013: 5]. Dinerstein et al. [2020] recommend establishing a global safety net against the intertwined threats of climate change and biodiversity loss. For this, they suggest the protection of 50% of land surface as intact, interconnected ecosystems.
The field of conservation has recently integrated the idea of connectivity, thus moving beyond efforts to simply set aside protected patches. Beginning in the 1990s, the study of ecological corridors has investigated their effect on biodiversity, their traction in the realm of public policy, and strategies for mapping corridors. Most studies show that ecological corridors are indeed effective in protecting species, although some species benefit more than others, depending on corridor design. Achieving political salience for such initiatives, however, appears to be as difficult as for any other conservation project.
Nevertheless, the mapping and development of ecological corridors is a way to enter into conversation with multiple stakeholders about conservation and the potential for collaboration. While establishing large protected areas often falls under the jurisdiction of national governments, restoring strips of land to connect these areas is well suited to the work of communities, local planners and public and private land managers. Within a regional, national or international context, establishing a network of connected ecosystems calls for coordination at multiple levels of government.
In 2009, the French government encoded the idea of ecological corridors into law, thus initiating a process for mapping out degraded areas and those with higher levels of ecosystem intactness. In collaboration with local and national actors, each regional government engaged in a process of mapping out land, whether public or private, where ecosystems are in relatively good condition and should be preserved, and where land is in poor condition and should be restored. The maps also include potential ecological corridor routes.
The combining of the regional maps into a whole country map thus creates a coherent ecological framework for land protection and restoration efforts. It is a tool to help local and regional planners prioritize conservation efforts in the places likely to best protect wildlife and ecosystem processes. While urban planning documents are expected to take these maps into account, the manner and extent to which ecological concerns are weighted in urban planning decisions is subject to local interpretation. Thus, the maps can serve as a tool for those motivated to protect the environment, yet they remain easily sidelined by the less motivated.
In their 2019 assessment of French ecological corridor efforts, Chaurand & Bigard suggest that part of the reason corridor development has not been more readily adopted at the local level is that much of the knowledge and enthusiasm for the idea exists at the national level among researchers and program leaders. To remedy this, they recommend investing in a system to relay information between local and national actors.
Similarly, critiques of the MesoAmerican Biological Corridor (MBC) have noted the shortcomings of a top-heavy approach [Dettman 2006, Grandia 2007, Harvey 2007, Beauvais & Matagne 1999]. Funded by the World Bank and led largely by international conservation organizations, development of this ecological corridor appears not to have effectively partnered with nor sought adequate input from local communities.
Consequently, the MBC has met with limited success in terms of biodiversity conservation. Connectivity for nine mammals in the Panama portion of the MBC is already severely fractured and predicted to decline further with planned road construction and other development projects on the horizon. In light of these findings, the authors of a Panama study [Meyer 2019] are not particularly optimistic about the condition of ecological connectivity in the MBC overall.
On a brighter note, Belize has recently approved a plan under the rubric of the MBC to establish a 110 km2 ecological corridor, which will be one of Central America’s largest. This was the outcome of collaboration between landowners, government and NGOs. South of the MBC, another ecological corridor is being developed in Brazil [Zanon 2020]. Thanks to the work of a local NGO that combines conservation with local community education, additional corridors will create a total of 44,920 ha2 of connected habitat. This is expected to increase the population of the threatened target species (black lion tamarin) by 30%.
These two examples notwithstanding, it is ironic that ecological corridors (designed to mitigate the effects of habitat destruction) are subject to the same pressures and obstacles as the problem they aim to remedy. Difficulty stems from the multiplicity of economic interests in land and a mosaic of ownership titles across a given landscape. Furthermore, the ecological concept of connectivity is not necessarily understood by the general public or policy makers, resulting in its low ranking as policy priority.
The ecological concept of connectivity is not necessarily understood by the general public or policy makers, resulting in its low ranking as policy priority.
The European Natura 2000 network, which helped spur France’s ecological corridor mapping initiative, is a continent-wide network of natural or semi-natural areas. However, the vast majority of these sites (86%) are unconnected to one another [Estreguil 2013], a fact which severely limits their value to wildlife. In spite of covering 18% of the land area of Europe, the nearly 28,000 sites in the network are isolated islands of nature within a generally hostile surrounding landscape.
There is the need to acknowledge nature as a system rather than individual parts. The establishment of N2K [Natura 2000] network (i.e. the sum of the individual sites) should be distinguished from the establishment of the overall ecological coherence of the network [Estreguil 2013: 5].
Furthermore, the areas themselves are not formally protected and can theoretically be developed; however, they are less likely to be developed by virtue of being a Natura 2000 site. A 2018 EEA assessment suggested that to “unlock” the potential of this ecological network, biodiversity policy should be thoroughly integrated by member states with economic development and other public policy domains, and that greater stakeholder engagement should be sought such as through citizen science biodiversity monitoring initiatives.
The enormous potential of ecological corridors fully depends on concerted efforts to communicate the meaning and importance of connectivity to stakeholders at every level of government and within the general public.
There is the need to acknowledge nature as a system rather than individual parts [Estreguil 2013: 5].
Ecological corridor article summaries
A “Global Safety Net” to reverse biodiversity loss and stabilize Earth’s climate, Dinerstein et al. 2020
Currently, 15.1% of land on Earth is conservation protected. This article maps out an additional 35.3% of land needing near-term protection, along with ecological corridor routes connecting these areas. Half of the planet’s land is needed to serve as a Global Safety Net to biodiversity loss and stabilize the global climate.
While the parallel crises of biodiversity loss and climate change have generally been approached separately, a key solution for two of the most pressing challenges of our time is the same: conserve enough nature and in the right places [Dinerstein 2020: 1].
The “right places” were identified by mapping areas with rare or endangered species, biodiversity hotspots[3], and places with distinct species assemblages. Onto this, the authors mapped areas where wild large mammals are still able to range widely and freely, a phenomenon that has become rare globally given the extent of anthropogenic land conversion, and areas of remaining intact wilderness.
The study also maps out a system of wildlife corridors to connect conservation areas. Only half of currently protected areas are connected. “Connecting all current terrestrial protected areas via potential wildlife and climate corridors (using 2.5 km as an average corridor width) adds 5,705,206 km2 or 4.3% of the terrestrial realm” [Dinerstein 2020: 4]. Assuming the additional lands identified in this study for conservation are formally protected, the amount of land needed for connectivity would be significantly reduced.
While large conservation protections require national leadership to achieve, the need to establish connectivity presents a role for local and regional actors to restore degraded lands in their midst.
The connectivity analysis offers a template to build from and engage local and regional entities in designing programs centered on restoring connectivity. This effort could merge with global habitat restoration and native tree-planting initiatives now under way [Dinerstein 2020: 7].
Focusing restoration efforts on degraded lands that can serve as wildlife corridors could help achieve other objectives, such as the Bonn Challenge. Similarly, massive tree-planting programs, if designed using native species and planted to restore corridors, riparian and coastal vegetation, and upper watersheds, could contribute to stabilizing climate and restoring connectivity [Dinerstein 2020: 7].
At the national level, countries could use the Global Safety Net framework to map out their own corresponding national safety nets. The 20 countries with the greatest role to play in establishing the Global Safety Net include: Russia, Brazil, Indonesia, the United States, Costa Rica, Peru, and Namibia.
Investments needed for the establishment and management of additional protected areas and restoration of degraded lands, while substantial, are small compared with enormous fossil fuel subsidies. The estimated $4.7 trillion per year in fossil fuel subsidies are expected to decline as the Paris Climate Agreement is implemented, making government resources available for restoring, rather than destroying, our global climate system [Dinerstein 2020: 7].
The authors emphasize that the conservation goals of the Global Safety Net are achievable, especially if indigenous people’s land rights are honored. One third of land identified for a Global Safety Net is managed by indigenous communities in a way that preserves biodiversity and regulates Earth’s atmosphere.
Guidelines for conserving connectivity through ecological networks and corridors, Hilty et al. 2020
The International Union for Conservation of Nature (IUCN), which created these guidelines, is an international environmental network founded in 1948 that provides conservation data, assessment and analysis to governments, NGOs and private entities. IUCN also manages the Red List of Threatened Species. This connectivity guideline is part of a series of best practices for protected area land managers.
Providing a definition and context for the importance of connectivity, the authors state:
‘Ecological connectivity’ is the unimpeded movement of species and the flow of natural processes that sustain life on Earth. This is not an overstatement. Without connectivity, ecosystems cannot function properly, and without well-functioning ecosystems, biodiversity and other fundamentals of life are at risk [Hilty 2020: xii].
Moreover,
Most global, regional and national targets for biodiversity conservation, climate change and environmental sustainability cannot be met unless ecological connectivity conservation is addressed [Hilty 2020: 48].
In short, ecological connectivity undergirds the conditions for life on Earth. The authors explain that the concept of connectivity reflects an evolution in conservation science. Previously, nature conservation consisted primarily of setting aside areas of undisturbed or minimally disturbed land. While protected areas remain the foundation of nature conservation, “they are no longer considered sufficient in many places. It is now understood that active measures must also be taken to maintain, enhance or restore ecological connectivity among and between protected areas and OECMs[4] [Hilty 2020: 2].”
Hence,
These Guidelines have been drafted to help clarify and standardize a shift in conservation practice from a narrow focus on individual protected areas to considering them as essential parts of large landscape conservation networks. This is done through creating ‘ecological networks for conservation’ that are specifically designed, implemented and managed to ensure that ecological connectivity is maintained and enhanced where it is present, or restored where it has been lost. Unless systems of protected areas and OECMs retain all essential ecosystem processes, they are not sufficient [Hilty 2020: 3].
The guidelines emphasize the importance of clearly defining one or more ecological objectives for establishing a corridor, such as to facilitate gene dispersal, migration, or adaptation to climate change for particular or multiple species. Clearly defined objectives allow for a corridor to be created in a way that leads to successful outcomes vis a vis the objectives. Primary objectives should relate directly to ecological connectivity, while complementary social or economic objectives (ecosystem services, such as flood and erosion control, enhancing crop pollination, for example) may also be included.
The toolbox for connectivity conservation includes various types of formal and informal recognition, national legislation, local and regional zoning regulations, conservation easements, conservancy design and transportation planning [Hilty 2020: 48].
The importance of connectivity is increasingly recognized in international treaties, and in national and sub-national planning and policy initiatives.
Until recently, connectivity legislation was rare at the national or even sub-national level. Now, countries such as Bhutan, Costa Rica and Tanzania, and sub-national jurisdictions such as California and New Mexico (USA), have enacted corridor legislation. Additionally, site-specific legislation has been enacted in some countries. For example, the South Korea Act on the Protection of the Baekdu Daegan Mountain System, 2003 (Act no. 7038), which came into effect in 2005, designates an area of 263,427 ha. Of this, 86% is made up of 183 existing protected areas and 14% consists of new buffer and core areas that create a biodiversity corridor along the main mountain range of the Korean Peninsula [Hilty 2020: 45].
However, mostly countries have not yet effectively integrated connectivity into policy and planning. Partly this is due to the complexity of establishing ecological corridors.
Connectivity conservation requires innovative implementation approaches to conserve lands and water within the conservation matrix – across patterns of resource use, jurisdictions, cultures and geographies [Hilty 2020: 48].
These guidelines are meant as a toolbox to help local, regional, national and international entities navigate that complexity.
‘Ecological connectivity’ is the unimpeded movement of species and the flow of natural processes that sustain life on Earth. This is not an overstatement. Without connectivity, ecosystems cannot function properly, and without well-functioning ecosystems, biodiversity and other fundamentals of life are at risk [Hilty 2020: xii].
Constructing ecological networks based on habitat quality assessment: a case study of Changzhou, China, Gao et al. 2017
Changzhou is a city near the Yangtze River delta on the east coast of China that has undergone extensive urban development. “From 2006 to 2014, the built-up area in the city increased by 25.68%” [Gao 2017: 2]. This study is part of an effort to boost biodiversity and ecosystem services in the city, which, at the time of the study, had a few protected patches but no corridors connecting them.
The authors identified potential corridors by comparing three different methods for assessing the level of resistance wildlife would face in moving across the landscape from one habitat patch to another. Corridors were identified by mapping out the paths of least resistance. Potential corridors consisted mainly of riparian greenspaces, followed by forest and farmland, and included between 3.45% and 16% built-up space, depending on the method used. Corridor width was assumed to be 30m. Connection of the most important protected patches should be prioritized in corridor construction.
Integrating priority areas and ecological corridors into national network for conservation planning in China, Liang et al. 2018
In contrast to the Gao et al. [2017] article (above), this study maps out an ecological network spanning the entire nation of China. Most such ecological corridor analysis has previously focused at the local and regional levels, according to the authors. They note that in addition to protecting biodiversity, ecological corridors (ECs) purify air, regulate climate, and “realize the movement of material, energy, and information in the ecosystem” [Liang 2018: 23].
This study identifies a couple of dozen high priority areas for conservation based on the existing diversity and quality of the landscape. These high priority areas encompassed seven ecotones (broadleaf forest, coniferous forest, shrub, herbaceous plant, sparse vegetation, wetland, water body), while built up areas such as cities were low priorities. The authors mapped these conservation priority zones against existing formally protected areas (which cover 15% of the country), finding only 19% overlap and, thus, revealing extensive conservation gaps.
The majority of China’s nature reserves were established without a clear planning framework, and couldn’t maximize efficiency of conservation targets. … important zones for species migration are not considered as conservation goals in the current nature reserve system [Liang 2018: 26].
The ecological corridors were identified by examining the pathways with the least amount of potential resistance (such as built infrastructure) to animals moving along them. The shortest routes were not necessarily chosen given the need to bypass urban areas. The map created through this study offers useful information for national conservation planning.
From a long-term conservation perspective, in view of the rapid habitat loss and biodiversity reduction, the ecological network represents a valuable tool to protect the biotope[5] and their ecological functions in China. In this regard, our results show the importance and need to develop a national protection network maintaining connectivity among them in order to achieve high cost efficiency [Liang 2018: 27].
A meta-analytic review of corridor effectiveness, Gilbert-Norton et al. 2010
Habitat fragmentation, a frequent consequence of habitat loss, is a primary threat to populations and species because isolated subpopulations are expected to experience reduced population viability and ultimately greater risk of extinction. Colonization and gene flow between habitat patches, however, can mitigate these effects [Gilbert-Norton 2010: 661].
This meta-analysis, consisting of 78 experiments from 35 studies, asked the question: Do ecological corridors increase movement between habitat patches, and how does that differ among taxa? The study’s results answer the first part of the research question affirmatively: “There was approximately 50% more movement between habitat patches connected by a corridor than between isolated habitat patches” [Gilbert-Norton 2010: 665].
Furthermore, corridors increase movement for all taxa. “Most corridors are created for terrestrial vertebrates, including birds, although our data suggest that invertebrates and plants also benefit from corridors” [Gilbert-Norton 2010: 665]. This study found that corridors work equally well for all taxa except birds, for whom the corridors were used less; however, birds still favored corridors compared to surrounding matrix.
While three quarters of the experiments showed corridors to be more effective for movement compared to the matrix landscape, 23% of experiments showed corridors were less effective. The authors suggest several explanations for this result. It’s possible that the “matrix habitat has been misclassified as nonhabitat for a study organism” [Gilbert-Norton 2010: 665], that the habitat quality of the corridor is not particularly high, or that the corridor is difficult to locate, given its small size compared to surrounding landscape. Furthermore, use of corridors varies by species.
That almost a quarter of the studies showed organisms used matrix habitat rather than corridors to move between habitat patches furthers the idea that although corridors may be used by many species, they are unlikely to be used by all species, and whether corridors are relevant for land managers may depend on the species of interest [Gilbert-Norton 2010: 665].
The authors also observed that organisms showed greater use of natural corridors (those existing prior to the study) compared to those created and maintained for the study. The real-world applicability of this, as the authors note, is that “it may be better to protect natural landscape features that function as corridors rather than attempting to create corridors” [Gilbert-Norton 2010: 667]. This highlights the importance of protecting natural or semi-natural lands from development.
Characterizing multispecies connectivity across a transfrontier conservation landscape, Brennan et al. 2020
Connectivity conservation pays attention to landscape connectivity to support animal species’ movements, keep ecological processes intact, and promote biodiversity. While the strategy of conserving connected, non-fragmented areas and respecting animals’ movement patterns is sound, in practice these plans are usually designed around a single species and its needs.
Brennan et al. looked at the limitations of a single-species focus, and evaluated the movement patterns of multiple species. They created connectivity maps for six large mammal species in the Kavango-Zambezi (KAZA) transfrontier conservation area straddling Angola, Zambia, Zimbabwe, Botswana, and Namibia, and assessed how each individual species’ connectivity maps correlated with that of the others.
This then allowed the authors to identify good ‘surrogate species for connectivity’ – that is, species whose connectivity maps were good representations of other species’ movements through the same area. They also took a look at different types of barriers to animal movements and determined that fences were the greatest obstacle to movement, while roads, rivers, and human-settled areas also deterred movement. Finally, they identified connectivity hotspots on the landscape, which are like bottlenecks through which multiple species pass due to barriers elsewhere. These connectivity hotspots are thus essential places to focus conservation efforts.
The researchers found the hyena and African wild dog to be the most apt surrogate species for connectivity, in spite of a popular practice of using elephants to determine the geographic targets of conservation efforts.
In our examination of connectivity across the landscape, female elephants were found to be only weakly correlated with the five other species in our study. Spotted hyena and African wild dog, in contrast, were strongly correlated with the greatest number of species. They also appeared to be complementary surrogates (i.e. they were correlated with different species), in which case combining their connectivity models could further extend the relevancy of connectivity conservation plans to other species. Thus, as both species are also charismatic, wide-ranging species of conservation concern, they may represent good umbrella species for connectivity in the KAZA region [Brennan 2020: 1707].
They went on to say that “while elephants may not be good surrogate species for connectivity across entire landscapes, they may still be effective as a surrogate at local scales where they can help protect local movement pathways or stepping-stone habitats for other species” [Brennan 2020: 1707].
Their conclusion is not that we should stop paying attention to elephants, which serve important ecological functions and are an iconic and culturally significant animal. Rather, we should look for gaps that may arise if we only conserve areas based on elephant movements, and put these techniques of comparing and combining different species’ movement patterns to use. Noting that animal movements and ecological dynamics play out at different scales, from entire landscapes and transnational parks to smaller corridors, they emphasized the importance of looking at connectivity for multiple species at multiple scales. They urged researchers and policy makers to take a more holistic multi-species approach to connectivity conservation.
Salvaging bycatch data for conservation: unexpected benefits of restored grasslands to amphibians in wetland buffer zones and ecological corridors, Mester et al. 2020
This study considers the effect of grassland restoration on amphibian populations in a 760-acre nature reserve – the Egyek-Pusztakócs Marsh System (EPMS) – established on former farmland in Hungary. The study shows that grassland restoration increased habitat range and quality for amphibians, extended hydrological supply, and limited genetic erosion among previously isolated populations. It also illustrates the role of smaller-scale ecological corridors.
Grassland restoration … creates corridors that maintain connectivity among the amphibian (sub)populations in the EPMS but it may also increase the permeability of the landscape to establish and maintain connections to other nearby metapopulations. Grassland restoration can thus also have an effect of minimizing genetic erosion of populations induced by isolation, which is one of the major causes of global amphibian decline [Mester 2020: 7].
Restoration can benefit amphibians by increasing the area of grasslands available for a variety of life activities such as foraging, burrowing, dispersal/ migration, or hiding from predators, aestivation and hibernation in the non-breeding period and by ensuring functional connectivity between wetlands both in the breeding and non-breeding periods [Mester 2020: 9].
Ecosystem service provision by road verges, Phillips et al. 2019
‘Road verges’ are strips of land on either side of roads and highways that are on average 3-4m wide, but can be as narrow as a few centimeters or many meters wide. “Road verges are commonly grassland habitats, but can be shrubland, forest or artificial arrangements of trees and horticultural plants, and we use the term also to include bare earth and freshwater bodies (e.g. ditches)” [Phillips 2019: 489]. They can also be barren ground or ditch. Not all road verges are managed; when management does occur, it is typically geared toward safety – clearing vegetation to enhance visibility.
There is currently an estimated 36 million linear kilometers of road network in the world, the length of which is expected to increase by 70% by 2050; thus, the total area of road verges will increase as well. “Road and road verge construction will displace habitats and cause many negative ecological and social impacts” [Phillips 2019: 494]. However, there is potential to mitigate that impact by maximizing the ecological value of road verges. Currently, “there may well be 270,000 km2 of road verge globally (0.2% of land), which is similar to the total area of the United Kingdom” [Phillips 2019: 492], with this surface area expected to grow.
While roads run like a network of veins across landscapes, causing widespread negative ecological impacts to adjacent areas, road verges form a parallel network and have the potential both partially to mitigate negative impacts of roads and to deliver environmental benefits [Phillips 2019: 490].
Where roads cut through natural habitat, the road verges will represent a net loss of biodiversity. By contrast, verges can increase biodiversity in highly degraded environments such as cities or industrial farmland. Furthermore, because of the growing urban population, the importance of natural and semi-natural environments will be increasingly important. Road verges designed to maximize ecological value thus have an important role to play in the health and wellbeing of urban residents.
Road verges might increase connectivity in highly modified urban and agricultural landscapes if road verges of suitable size, habitat quality and continuity are created alongside roads, at least for species that are highly mobile or able to persist in narrow, linear habitats [Phillips 2019: 495].
While roads often act as barriers to wildlife and ecological connectivity, ecological corridor design could benefit by taking into account the potential benefits of road verges.
If road verges were integrated into such [ecological corridor design] projects, they might play an important future role in increasing connectivity between natural and semi-natural habitats, particularly across otherwise habitat-poor, human-dominated landscapes where roads often occur [Phillips 2019: 495].
Road verges designed to maximize ecological value thus have an important role to play in the health and wellbeing of urban residents.
Fence ecology: frameworks for understanding the ecological effects of fences, McInturff et al. 2020
Conceptually the inverse of wildlife corridors, fences aim to disconnect. They are built to separate people across national borders, livestock from predators, to delineate property lines, and even to protect wildlife conservation reserves. Globally, fences are ubiquitous, more prevalent even than roads, and proliferating. Yet their ecological impact is relatively unstudied.
Fences are often framed as a management tool rather than a globally significant ecological feature, and they are a notable omission from efforts to map global infrastructure, including the human footprint [McInturff 2020: 971].
This analysis reviews 446 studies starting from 1948 on various types of fencing to assess impacts; however, most of the studies focus on the effect of fencing on particular species (specifically, those the fencing is meant to protect), rather than on multiple species, communities or ecosystems.
Conservation and restoration fences, for example, have support within the literature for their beneficial effects for wildlife and sensitive plant species for which they are built, making such species “winners” in the fencing game. On the other hand, there is a critical lack of information on species that are not the targets for which fences are built, and our review shows that only 10.8% (48 of 446) of studies focus on nontarget species [McInturff 2020: 975].
While fences aiming to protect particular species usually achieve that goal, they inevitably hurt other species.
… often the clearest winners because of fencing are the species that humans value most, whereas losers are inevitable but may remain invisible [McInturff 2020: 975].
Broadly speaking, fences favor generalists and disturbance specialists, many of which are invasive, as well as small and small-ranged, nonmigratory species. Fences therefore heavily restrict what makes a species a winner [McInturff 2020: 975].
The deleterious effects of fences include: impeding migration, reducing gene flow between populations, restructuring community composition and obstructing interspecies interactions, such as between predators and prey. These community-level changes can have ripple effects in the ecosystem. For example, livestock fences effectively excluding dingoes in Australia led to this large predator’s eradication. “Without dingoes, researchers have tracked a continental-scale mesopredator [mid-level predator] release that has altered biodiversity and habitats over enormous areas of Australia” [McInturff 2020: 979].
While fences limit certain interspecies interactions, they concentrate others:
Even where conservation or restoration fences seemingly protect whole habitats, research still points to differential outcomes for constituent species. In addition, pathogens and parasites may spread more rapidly where species interactions are concentrated within reserves. In central Kenya, for example, smaller fenced reserves produced higher gastrointestinal parasite infection rates among impala [McInturff 2020: 977].
The authors recommend a greater research focus on the cumulative ecological effects of fencing, policy that limits fence building and encourages fence removal or fence design that is more wildlife-friendly. They caution that fencing is among the major drivers of anthropogenic change.
As fencing continues to rapidly proliferate, there is potential for a dangerous future in which fences simultaneously alter ecological processes at multiple scales, likely producing more losers than winners, and potentially resulting in ecosystem state shift or collapse [McInturff 2020: 977].
Livestock fences effectively excluding dingoes in Australia led to this large predator’s eradication. “Without dingoes, researchers have tracked a continental-scale mesopredator [a mid-level predator] release that has altered biodiversity and habitats over enormous areas of Australia” [McInturff 2020: 979].
European Context
Status of the Natura 2000 network (from State of Nature in the EU report), EEA (European Environmental Agency) 2020
While not an ecological corridor per se, the Natura 2000 network is the largest coordinated network of conservation areas in the world. Covering 17.9% of Europe’s land area and nearly 10% of the continent’s marine areas, the network includes 27,852 sites with an area of 1,358,125 km2. The terrestrial portion of the Natura 2000 network is mostly covered by forests and transitional shrublands. It also includes grasslands and wetlands, as well as pastures, cropland and a small amount of artificial surface (developed/built land).
Member States need to ensure that sufficient protection and appropriate measures are implemented in Natura 2000 sites for habitats and species of community interest and that they form a functional network [EEA 2020: 109].
However, the sites are not strictly protected by virtue of being part of the network. In fact, the sites include a variety of land uses.
Within the network, arable land and permanent crops have increased, while grasslands and forests have decreased. … Pastures and mosaic farmland (with approximately 18 %) and inland wetlands and water bodies (with approximately 10 %) have been extensively transformed into arable land and permanent crops both inside and outside the network. Recent research has shown, however, that high nature value (HNV) farmland inside Natura 2000 sites is less likely to be converted into artificial surfaces than such farmland outside the network and is more likely to maintain its pattern of mosaic farming [EEA 2020: 113].
This assessment of the network’s effectiveness found that “species and habitats are more likely to have a good conservation status if they are well covered by the Natura 2000 network” [EEA 2020: 121]. However, limited monitoring inside and outside the network prevents a more detailed analysis of Natura 2000’s effectiveness. Furthermore, due to a limited implementation of conservation measures, the network’s potential has not yet been fully “unlocked,” according to the report.
To improve Natura 2000’s potential, the authors recommend, among other measures, improving connectivity between protected areas. Noting that sites chosen for inclusion in the network are often motivated by economic rather than ecological interests.
Incoherent planning and site selection approaches between and within Member States has led to insufficient functional connectivity and spatial connectedness between neighboring countries and habitats and gaps in coherence within Member States. This highlights the need to increase connections between protected areas and the level of protection beyond the site [EEA 2020: 122].
Also recommended is increasing stakeholder participation, such as through citizen science monitoring initiatives, and better integration of biodiversity protections into other policy domains.
The resulting low awareness of the diverse benefits produced by the Natura 2000 network is often compounded by a long-standing conflict between economic or political interests and conservation goals. There is thus an urgent need to increase coherence between biodiversity policy and other policy areas, such as in the fields of agriculture and economic and rural development, and create a more integrated approach to address potential conflicts and trade-offs between various interests while fostering synergies [EEA 2020: 124].
The report’s summary conclusion recommends increasing marine and terrestrial conservation areas in the Natura 2000 network to 30% each, strictly protecting these areas, and improving connectivity among them.
Blue and green corridors [Les trames vertes et bleues] in France, Ministry of Ecological Transition 2017
Spurred to action by the European Union and a vision for a pan-European ecological network, France encoded the idea of the “trames vertes et bleues” into law in 2009. The national government worked with all the regional governments to develop maps showing areas with the highest levels of biodiversity. This includes protected areas, stretches of coastline, riparian zones, woods, and other undeveloped areas, whether public or private. The maps also show ecological corridors – both those in good condition needing to be preserved, and those that are highly degraded and requiring restoration.
The regional maps are meant to be integrated into urban planning at the level of city and county (department). Rather than being a regulatory tool, the maps are an information source allowing urban development to proceed in a way that limits impact on biodiversity. The ecological corridor initiative is designed as an invitation and encouragement to local governments, organizations, businesses and individuals to collaborate and to act in favor of biodiversity.
The preservation and restoration of ecosystem connectivity entails acting everywhere possible: in rural environments, in aquatic ecosystems and in urban areas [MTES 2017, translation].
Articulating the politics of green and blue infrastructure and the mitigation hierarchy for effective biodiversity preservation in France [Articuler la politique Trame verte et bleue et la séquence Éviter-réduire-compenser: complémentarités et limites pour une préservation efficace de la biodiversité en France], Chaurand & Bigard 2019
This article reviews the historical development of two pieces of environmental legislation in France – the use of the “mitigation hierarchy” to assess and limit environmental impact in project development and the promotion of ecological corridors. Theoretically, these two laws overlap when urban development projects in proximity to areas of ecological significance use the mitigation hierarchy (avoid, reduce, compensate) to ensure these zones are protected within the scope of the project.
1976: “Protection of Nature” law in France introduced the mitigation hierarchy, aiming to avoid or reduce harm to the environment, or to compensate if harm is unavoidable.
1992: Concept of “biodiversity” entered public discourse internationally, following the Earth Summit in Rio, Brazil.
1996: France ratified European ecological corridor strategy.
1999-2000: Concept of “sustainable development” emerged in France.
2004: National strategy for protecting biodiversity adopted.
2007: “Grenelle de l’Environnement” meeting created the “Trame Verte et Bleue” (TVB) policy (green and blue infrastructure, encompassing ecological corridors)
2016: Biodiversity law enacted, creating national agency and regional committees on biodiversity
In spite of this policy evolution, commitment to ecological corridors has yet to move from a “TVB papier” to a “TVB de projets et d’action.” In other words, much discussion and mapping efforts have not yet resulted in the development of the imagined ecological corridor network. The authors speculate as to why this is so, explaining that the resources and coordination needed for enforcement are lacking. Even though “the creation, preservation and restoration of ecological connectivity” has been integrated into urban planning code, such considerations are often sidelined. Furthermore, definitions are vague: the objective of the TVB is the “good condition” of ecological continuity, but “good condition” is not defined. Lastly, taking action in defense of ecological continuity requires pro-active collaboration among levels of government from local to regional to national.
The authors propose better integration of these to policy tools. For example, the TVB designates certain non-protected areas throughout the country that are ecologically functional and serve a role in the eco-corridor network as key areas to “preserve.” With better communication between this TVB framework, the mitigation hierarchy could be applied at the level of “avoiding” harm to places designated as preservation priorities, but lacking formal “protected” status. In projects where harm is unavoidable, the mitigation hierarchy could be applied at the level of “reducing” harm to maximize the percentage of remaining green space as well as the permeability to wildlife of the built structures (such as passageways through fences). The “compensation” level of the mitigation hierarchy could be applied in the context of regenerating ecosystem function to areas designated in the TVB schema as needing ecosystem restoration.
The authors note that advocates for the TVB are clustered at the national level and within research institutions, while the people responsible for urban planning decisions are local and are not necessarily well versed in the scientific framework for the TVB. Local actors tend to focus on priorities other than ecological continuity. One measure to address this, according to the authors, would be the training of local “relays” to transmit knowledge of ecological principles vis a vis the TVB to local urban planners.
Woods and hedgerows of Brittany countryside [Le bocage Bretagne], OEB (L’Observatoire de l’Environnement en Bretagne) 2018
Produced by a regional consortium on the environment in Brittany, France, this report describes the ecological value of woody strips encircling agricultural fields and enmeshing the countryside, their decline, and ways to incentivize their protection.
Brittany is a heavily agricultural region that also features a long stretch of coastline where urban development and expansion is ongoing. Due to mechanization and enlargement of farm fields, average parcel size has increased since the 1950s, shrinking the extent of woody hedgerow (“bocage”) between fields. Between 1996 and 2008, the total length of hedgerow decreased 12%. This change is concerning because Brittany is already one of the most fragmented and least wooded parts of France.
The report explains the value of the bocage is its provisioning of habitat, connectivity between habitats, biodiversity, erosion control, groundwater recharge, and flood mitigation. Half the population of Brittany lives in areas susceptible to flooding. Furthermore, at least five endangered animal species depend on the habitat created by the bocage. Protection of this woody network is key to remedying both problems, while also providing direct benefits to farmers, such as habitat for pest predators.
The form and shape of the bocage varies throughout the region, but can include grasses, bushes and/or trees, forming one or more layers of vegetation; and heterogenous landscape features such as berms, ditches, logs, and rocks/boulders, which create microhabitats.
The network of hedge and berms, accompanied by fields, ponds and wetlands, constitutes an important natural environment because of its heterogeneity and potential for complex exchanges. It has the particularity of being able to reach a myriad of increasingly isolated natural spaces in the heart of a changing agricultural countryside subject to ongoing urbanization. Similar to a forest edge environment, the richness of the bocage can be explained by the diversity of habitats it adjoins [OEB 2018: 8, translated].
Regulatory and incentive programs payments have sought to encourage farmers to preserve their hedgerows. However, the authors suggest that a stronger economic valuation of these linear woods is needed to protect and expand them. They suggest strategies for stimulating the market for firewood and other products harvested from sustainably managed hedgerow, where biodiversity protection is an explicit aim and co-benefit.
Americas Context
Shaping land use change (LUC) and ecosystem restoration in a water-stressed agricultural landscape to achieve multiple benefits, Bryant et al. 2020
In spite of its obvious benefits, agriculture, which covers one third of the Earth’s land surface, damages biodiversity and ecosystem services. In some regions, land degradation and depletion of water resources from irrigation have been so great that historical levels of food production in these regions risk decline. Some areas of previously productive farmland will likely need to be retired from use. Within this context, maintaining and enhancing natural corridors and promoting semi-natural, multifunctional landscapes can significantly contribute to recovering biodiversity and mitigating air and water pollution.
Using California’s San Joaquin Valley (SJV) as a case study, this paper illustrates a pragmatic approach to incorporating ecological corridors into working landscapes. The authors offer a new analytical approach that simultaneously incorporates resource-constrained (water, in this case) land-use change (LUC) modeling within the planning and optimization process. The goals are to simultaneously:
Meet water-use-reduction policy goals for the area under study within the next two decades
Identify lands for retirement that are (1) likely to be retired anyways and (2) offer high-value habitat for native species and biodiversity.
Over the past century, SJV has been transformed into one of the largest agricultural economies in the world. However, this economic success has been costly to the SJV in several ways, including:
Damaged infrastructure: high rates of groundwater extraction in the SJV have led to groundwater overdraft and unreplenished aquifers, resulting in large-scale land subsidence. Most of the subbasins in the SJV are categorized as critically overdrawn, and some regions have sunk over 8 meters since the early 20th century; this land subsidence further imperils water availability and quality by impacting water storage and delivery infrastructure.
Decreased human health, as a result of impaired air and water quality, leading to chronic health problems
Threats to wildlife and biodiversity; for example, some species have lost up to 98% of their habitat range, and over 35 native species are listed as threatened or endangered
“In response to these challenges, and amid significant drought-driven fallowing, California passed the Sustainable Groundwater Management Act (SGMA), which … obligates locally governed groundwater subbasins to develop plans that will achieve sustainable groundwater use over the next two decades” [Bryant 2020: 2]. To meet these requirements, many subbasins will meet with severe groundwater pumping restrictions. If these areas are not able to coordinate their pumping activities and augment water supplies, the SGMA may require a reduction in cultivation area through fallowing or permanent retirement.
Given the likely retirement of 86,000 ha of irrigated agricultural land, the authors explore spatial optimization of retired land for conservation efforts. They find that a key strategy is the identification of areas that were destined for retirement from cropping which could be shifted to restoration and habitat enhancement, as well as possibly shifting some areas destined for retirement that have “low habitat value” with regards to wildlife for areas with “high habitat value.” Priority restoration areas identified in this analysis include many that are contiguous and located near designated wildlife areas.
Importantly, the analysis presented here is “explicitly organized to help inform engagement between conservation actors and agricultural land managers about how habitat goals can be achieved in ways that benefit communities in the SJV” [Bryant 2020: 3]. The potential positive futures indicated by such analysis can be used to identify opportunities for collaboration between the conservation and agricultural communities, with a goal of guiding land use change toward achieving multiple benefits, such as recovery of imperiled natural communities, resilient agricultural production, and improved public health outcomes.
While it poses a great challenge, the impending transformation in the SJV also presents an opportunity to proactively shape the landscape in ways that not only ensure agricultural and water sustainability, but also achieve many other socio-ecological goals, such as biodiversity protection and improved human health. However, given that achievement of many of these objectives is determined by where things happen on the landscape (rather than simply the aggregate amounts of cultivation, retirement, or restoration), stakeholdersneed a systematic way to integrate these objectives to informmulti-benefit spatial planning [Bryant 2020: 4].
Integrating Agricultural Landscapes with Biodiversity Conservation in the Mesoamerican Hotspot, Harvey et al. 2007
The fate of biodiversity within protected areas is therefore inextricably linked to the broader landscape context, including how the surrounding agricultural matrix is designed and managed [Harvey 2007: 8].
Rather than discussing ecological corridors per se, this article emphasizes the importance of a whole-landscape approach to biodiversity conservation. Pointing out that protected nature reserves are weakened when isolated, these authors focus on the role of the entire surrounding agricultural matrix for restoring connectivity.
In contrast to the prevailing trend of managing protected areas and productive lands separately, we propose integrated landscape management in which conservation and production units within the agricultural matrix are managed jointly for long-term sustainability. We do not advocate agricultural intensification to spare further forest conversion because this approach is unlikely to have the intended outcome, for reasons discussed. Instead, conservation efforts should be based on the recognition that how agriculture is conducted and how different land uses are distributed spatially and temporally determine the region’s biodiversity. Lasting conservation will therefore require alliances among conservation biologists, farmers, and land managers to actively plan the future of Mesoamerican landscapes [Harvey 2007: 9].
The sections of the agricultural matrix the authors prioritize for biodiversity conservation include areas near riparian and other key ecological corridors, and they recommend leveraging support for the Mesoamerican Biological Corridor to spur regional action. Priority conservation areas are also more likely to encompass landscapes with a high diversity of indigenous and traditional cropping systems than those dedicated to industrial agriculture because “the chances of reconciling farming and biodiversity conservation there [agro-industrial systems] are slim” [Harvey 2007: 10].
The authors argue that, in contrast to large-scale, export-oriented industrial production, small-holder and indigenous agricultural systems are more compatible with biodiversity conservation, increased food production and rural income. The authors propose economic and regulatory instruments and greater regional collaboration to enhance native tree cover on farms, promote traditional, ecologically based farming practices, and to protect remaining intact habitat and restore degraded lands. The overarching vision is to accomplish conservation and agricultural production objectives for the region in mutually reinforcing ways.
The fate of biodiversity within protected areas is therefore inextricably linked to the broader landscape context, including how the surrounding agricultural matrix is designed and managed [Harvey 2007: 8].
The concept of green corridor and sustainable development in Costa Rica, Beauvais & Matagne 1999
The concept of sustainable development presumes that human economic systems and overall wellbeing depend on functioning ecosystems. Therefore, ecological rhythms should not be transgressed to the point that they fail to provide the vital services needed today and in future generations.
According to this model, economic development becomes a necessary but insufficient condition for society to progress [Beauvais & Matagne 1999: 6, translated].
Costa Rica holds at least 5% of the world’s species, in spite of making up 0.03% of its land surface. As an isthmus, Costa Rica is influenced by weather patterns from two oceans, as well as a north-south migration route. In addition to this, its mountainous terrain creates a heterogenous mosaic of habitats and niches. However, the country has been severely deforested. Forest covered 66% of land surface in 1940, and only 25% by 1987; the loss of forest led to extreme erosion.
As presented in this article, an ecological corridor consists of at least two protected ecosystem patches that are connected by a protected vegetated strip of at least a few kilometers in width, and the whole area surrounded by a buffer zone. Multiple units of two connected patches could in turn be connected, stretching into a corridor that the whole length of the country. A green Costa Rican corridor could connect to green corridors in adjacent countries, ultimately recreating the entire isthmic corridor that once existed.
However, the tone of this article is not optimistic about conservation, citing several political obstacles to conservation and ecosystem restoration. According to the authors, a combination of neocolonialist pressure, poverty, corruption, and capitalistic interests allow for trees to be cut even in protected areas and prevent the establishment of new protected areas and corridors.
The Mesoamerican Biological Corridor in Panama and Costa Rica, Dettman 2006
At the end of the 1980s, as a period of severe conflict in Central America was winding down, most countries in the isthmus signed the Charter Agreement for the Protection of the Environment, which established a sustainable development commission. At the same time, the “Central American Protected Areas System (SICAP) created approximately 11.5 million hectares of protected areas throughout the region” [Dettman 2006: 18].
This paved the way for international attention and investment in what became the Mesoamerican Biological Corridor (MBC). The original intention was to promote biodiversity and economic development in tandem through investment in local projects. However, in the 2000s, the international coordinators of the MBC shifted the focus from biodiversity protection (although the establishment of ecological corridors remains an objective) to a greater emphasis on economic development. This author explains that the institution’s decision-making process is overly top-down, and would benefit from input from local people who are implementing projects on the ground.
Between Bolivar and Bureaucracy: The Mesoamerican Biological Corridor, Liza Grandia 2007
Written by an anthropologist working in Central American conservation efforts for more than 10 years, this article describes the Mesoamerican Biological Corridor (MBC) project as having succumbed to a neoliberal agenda. Although originally spearheaded by Central American environmentalists, the notion of cross-border environmental collaboration was adopted by the World Bank and large international conservation organizations working in Central America in the 1990s. In the hands of these international giants, the biological corridor initiative became a bureaucratic, top-down project, deaf to the voices of local communities.
With all this new bureaucracy, a broad and unfocused agenda, and the challenges of high-level political coordination, the MBC quickly lost its potential to inject a stronger environmental justice component into regional biodiversity conservation programs.
Indeed, the MBC that emerged from the World Bank’s incubator was decidedly more business-oriented than initial proposals for Central American environmental coordination at the 1992 Earth Summit [Grandia 2007: 486].
In this context, the MBC’s conservation efforts have focused more on securing land for protected parks and less on community-based initiatives. The author suggests that in addition to land protection, the MBC should engage farmers in capacity building for eco-agriculture with a view toward achieving landscape-wide ecological connectivity.
The corridor approach might also draw greater attention to the agrarian contexts outside of parks, which may be just as ecologically important as what happens inside parks. By bringing agricultural systems into conservation debates, corridors may present new opportunities for supporting fair-trade projects and other small-scale agroforestry systems compatible with conservation. In other words, corridors could offer a method for moving beyond protectionism to embrace a mosaic vision for conservation that includes local people more explicitly. Corridor planning frameworks also could provide more democratic conservation forums [Grandia 2007: 484].
Effectiveness of Panama as an intercontinental land bridge for large mammals, Meyer et al. 2019
One of the world’s largest corridor projects is the Mesoamerican Biological Corridor (MBC). Initiated in the 1990s, the MBC aims to connect protected areas between southeastern Mexico and Panama [Meyer 2019: 2].
The ecological functionality of the MBC has not been much assessed, in part because direct approaches to measuring connectivity are costly and challenging. In this study, researchers used a simpler, indirect approach to measure forest connectivity through Panama for nine mammals. Using camera traps (cameras that are automatically triggered by a change in some activity in the vicinity, like the presence of an animal), they documented the presence (or absence) of these mammals in 28 forest sites along the Atlantic coast. The corridor was presumed to be functioning for animals whose presence was established across the entire length of the monitored range.
The species monitored in this study are forest specialists, including ungulates, carnivores and an insectivore, all of which are threatened by habitat loss and hunting, some more than others. Of the 43% of land in Panama that is forested, 44% is protected, mostly along the Atlantic coast. Steady economic development threatens remaining ecosystems with investments in large infrastructure projects, real estate, mining, tourism, and energy.
Large mammals are an indicator species for the success of conservation efforts. This is because:
Large mammals are generally at a higher risk of extinction in disturbed landscapes than other taxa because their large home ranges and low population densities at broad spatial scales mean their populations are more likely to be fragmented and because they are heavily hunted [Meyer 2019: 3].
The researchers found that even the four most prevalent species in the study are susceptible to population fragmentation by any further habitat loss.
We found that there was little connectivity for white-lipped peccary [a pig-like animal] and white-tailed deer and that, although 4 of the species (collared peccary, red brocket deer, puma, and ocelot [a wild cat]) occurred in most of the sites, a small decrease in connectivity of 20% would disrupt their continuous distributions across Panama. White-lipped peccary, giant anteater, white-tailed deer, jaguar, and tapir [a pig-like animal with a short trunk] had lower probability of occurring in all the sites and were therefore even more at risk of connectivity loss, as evidenced by >1 connectivity gap. This indicates the MBC may not function for the majority of species, especially considering we did not account for potential effects of hunting, which would make connectivity even more challenging [Meyer 2019: 8].
Citing imminent development projects, such as a new road that will pass through the forested northern coast and associated large hotel projects, the authors predict that ongoing loss of connectivity is likely. Moreover, the deteriorating condition of the corridor in Panama bodes poorly for the MBC overall.
The disruption of connectivity between tropical forests in Central America, and hence the possible separation of mammal populations, is an indicator of the overall functioning of the MBC for wildlife [Meyer 2019: 11].
Belize creates one of Central America’s largest biological corridors, Dasgupta 2018
The Belize government approved a plan in February 2018 to create a 110-square-kilometer biological corridor connecting two nature reserves in the northeast of the country. This outcome resulted from collaboration among NGOs, the government and private property owners. The latter agreed to conserve (to not deforest or otherwise degrade) the parts of their land that would become part of the wildlife corridor. In exchange, the government would not collect taxes on this land. This corridor, which was initiated in the context of the larger Mesoamerican Biological Corridor project, is meant to protect jaguars, cougars and tapirs, among other wildlife.
The woman building the forest corridors saving Brazil’s black lion tamarin, Zanon 2020
“The tamarin is unable to do anything to save its own species. And we, human beings, are the ones who are destroying their environment,” says conservationist Gabriela Rezende. “So, when I got the opportunity to see this animal in the wild, I felt partly responsible for its future.”
Rezende works with the Institute for Ecological Research in the Brazilian state of Sao Paolo to create ecological corridors connecting the forest fragments where the world’s only 1,800 black lion tamarin live in isolated populations. Since 1984, the institute has worked to protect this small primate species, which had reached a low point of 100 individuals and was listed as “critically endangered.” In addition to research and forest restoration, the institute also does environmental education with the local communities. This includes collaboration on nine tree nurseries administered by local people as small businesses that also provide school kids the chance to learn about local forest species that will be planted in corridors.
Leveraging a state policy requiring 20% of privately owned property to be in nature reserves, Rezende worked with landowners to identify patches to be restored that would physically connect forest fragments. Once corridors are complete, the total amount of land in connected habitat will be 111,000 acres. Rezende estimates the black lion tamarin population could increase 30% once it’s able to use the whole forest corridor. The restoration project will benefit other species too, including anteaters, tapirs (a pig-like animal with a short trunk), pumas, and ocelots (another wild cat species).
A well-watered mulberry tree has been credited with averting the danger of destructive wildfires from destroying Brett Hawkins’ home during 2020’s unprecedented fire season in Australia. When massive fires raged through the bush through the summer, many homes were completely engulfed. However, Hawkins attested that when he returned to his home after evacuating,
‘I could see straight away the house was intact — the roof was intact, but everything else around it was burnt, with the exception of the mulberry tree.’ He described the stark scene greeting him upon arriving back home, ‘It was apocalyptic,’ Mr. Hawkins said. ‘There was not a tree left, ash on the ground and smouldering embers everywhere.’ But among the blackened trees, Mr. Hawkins found his mudbrick house and mulberry tree in full leaf [Aubrey 2020].
In the season’s drought, he had been rationing water, but sparing some to keep his tree hydrated and healthy, which may have been a contributing factor in its resistance. According to the article, “Mr. Hawkins believed that by heavily watering the tree, combined with luck regarding which direction the fires came, the full heat of the bushfires was shifted.” [Aubrey 2020]
Another important feature is the mulberry’s lack of dried leaves and brush at its base that might pose a danger of igniting. Other species, like eucalyptus, with oily leaves that could dry out, or pine trees whose long branches catch dry leaves, are less ideal.
A tree expert from the Fenner School of Environment at the Australian National University analyzed some of the possible factors leading to the survival of this mulberry tree and what it might teach those wishing to fortify the fire resilience of their homes and properties. “While there does not seem to be a clear answer on what to plant to ‘fire-proof’ your house, Professor Brack said a well-watered tree, with a clear trunk and no loose, dry leaves or branches is a good start” [Aubrey 2020].
A project in Queensland, Australia has met with success in its efforts to rehydrate the landscape on the farmland property of Worona Station, improving biodiversity, water retention, and resistance to erosion and fire. Worona Station had been degraded and faced serious erosion issues, so Chris Le Feuvre, the owner, partnered with consultancy groups of NQ Dry Tropics and the Mulloon Institute in a project to rehydrate his land.
The project team has used a combination of planned grazing and small, low-tech dams to combat erosion problems. The grazing technique involves:
Splitting paddocks into small sizes and using large mobs of cattle grazing on rotation … grazing pasture more intensively while giving it longer to rest, [thereby] increasing carrying capacity.
Grazing in this way (which is evocative of Allan Savory’s Holistic Planned Grazing methodology) has resulted in increased pasture species diversity and boosted plant growth, allowing the Le Feuvre to double his herd size. Planned grazing has also reduced sediment runoff from the property. Sam Skeat, a grazing officer with NQ Dry Tropics, attests to the importance of grazing.
The plug-and-pond technique — also known as leaky weirs — involves small dam-like structures to lift the bed level of the water, which is then run onto the floodplain to grow pasture and recharge aquifers. While weirs have been strategically constructed, Mr. Skeat said grazing management was the most important tool to improve water retention in a landscape. ‘If you can use cattle as a tool to regenerate the grassland, you’ll get more infiltration, slow the flow, hold water up in the landscape and have you growing grass for longer’ [Major 2020].
Rehydrating landscapes can improve their resilience to extreme events, and improve their quality in the face of chronic problems like erosion. According to the Mulloon Institute Chairman Gary Nairn, the issue of degraded gullies and streams is a national concern. Gullies are created when parched land is unable to absorb rainwater, allowing it to run off. The sediment-filled runoff ends up in the ocean, polluting it.
Nairn sees land rehydration through planned grazing and related techniques as a better solution than building a massive, industrial-scale dam to retain water. The Australian government has been looking into building new large dams. Levels at Warragamba Dam, which supplies about 80 percent of Sydney’s water, have dropped to less than half capacity.
‘We’ve been able to demonstrate in Mulloon, if we repaired and rehydrated the catchment through to the Sydney water supply, you could store the equivalent of Warragamba Dam,’ he said [Major 2020].
Nader et al. survey herbicides, prescribed fire, mechanized treatments, hand cutting, and grazing animals as fire management techniques. Managing vegetation involves “changing the plant community to decrease the flame height when fire occurs,” favoring native species that may be more resilient to fire, and altering the landscape to create fuel breaks, which are patches across which it is hard for fire to jump [Nader 2007: 18].
Focusing their analysis on grazing and the contexts in which it is most useful, the authors note that there are many site and animal specific factors to take into account for successful implementation.
[Grazing]is a complex, dynamic tool with many plant and animal variables, and it requires sufficient knowledge of the critical control points to reach treatment objectives. Those control points involve the species of livestock grazed (cattle, sheep, goats, or a combination); the animals’ previous grazing experience (which can affect their preferences for certain plants); time of year as it relates to plant physiology (animal consumption is directed by the seasonal nutrient content); animal concentration or stocking density during grazing; grazing duration; plant secondary compounds; and animal physiological state [Nader 2007: 19].
Grazing has the advantage of keeping nutrients in the ecosystem, unlike mechanical methods that harvest vegetation to be sold as biomass chips (like wood chips). This means that when animals digest vegetation and excrete on the landscape, they participate in the local nutrient cycle. Animals also trample soil, which can crush fine fuel and mix it into the soil, where it cannot contribute to ignition, which reduces one contributing factor to persistent and destructive blazes. Animals do more than just remove extra vegetation – they can have many beneficial interactions within a given ecosystem.
Any grazing plan designed for fuel reduction needs to consider the grazing impacts on parameters other than just simply reduction. The effects of the grazing management should be studied for their impact on water quality, compaction, riparian vegetation, disease interaction with wildlife (bluetongue, pasturella), and weed transmission. The positive aspects of grazing over other treatments also should be weighed, including recycling of nutrients into the products of food and fiber [Nader 2007: 22].
By introducing grazing animals into a landscape or agricultural system, managers can affect biodiversity in complex ways. The authors mention that “Hadar et al. reported that light grazing increased plant diversity on treated sites. Thus, when proposing a stocking rate for treatment consumption, the environmental impact needs to be considered” [Nader 2007: 22].
Nader et al. conclude that “grazing is best used when addressing vegetation with stems of smaller diameters that make up the 1- and 10-hour fuels. These two fuel classes are important because they can greatly impact the rate of spread of a fire, as well as flame height” [Nader 2007: 19]. While they call for further research to validate anecdotal accounts supporting grazing and understand its best practice, they maintain that “prescribed grazing has the potential to be an ecologically and economically sustainable management tool for reduction of fuel loads” [Nader 2007: 20].
This study examines the positive effects of beaver damming on the resistance of landscapes to wildfire damage. The authors find that in riparian corridors (areas along rivers), the presence of beavers and their dams can create refuges that withstand blazes that consume surrounding vegetation.
Beavers play an important role in wetland habitats and are known as ecosystem engineers for the way they can shape landscapes with their activities. Beaver dams slow down water moving across a landscape, holding it in place for longer and allowing water to infiltrate into the soil, which raises water tables.
The combination of building flow obstructions (dams), accumulating water (ponds), and spreading that water out in the landscape (channels) gives beavers the unique potential to modulate environmental extremes such as flood and drought. When it comes to water, beavers slow it, spread it, and store it.
Due to the fact that beaver channels and dams spread water out in the landscape and store it broadly in adjacent soils, the vegetation near beaver ponds doesn’t experience as much reduced water availability during drought. Drought-stricken vegetation burns more easily than lush, green vegetation, so it follows that the vegetation around beaver ponds would be more difficult to burn than vegetation around undammed creeks [Fairfax and Whittle 2020: 1].
Fairfax and Whittle observed the effects of beaver dams on preventing fire spread to the areas where they had built dams, examples of which are shown in satellite imagery below.
The authors quantify the effects of beaver activity in fireproofing areas by examining the Normalized Difference Vegetation Index (NDVI) observed in satellite imagery before, during, and after wildfire years in the American West. They found that while vegetation is able to reestablish itself a year after fire damage regardless of beaver activity due to its own resilience to fire, areas in beaver dammed zones maintained vegetation even during wildfires, demonstrating actual resistance to blazes, not just the ability to recover after damage. They note how vital this is for those ecosystems and the life within them.
These ribbons of fire-resistant riparian corridor may be particularly important for species that are unable to physically escape wildfire. They can provide temporary habitat for fish, amphibians, reptiles, small mammals, wild and domestic ungulates, and birds that are unable to outrun/outfly the spread of flames. While we found that beaver activity does play a significant role in maintaining vegetation greenness during wildfires, it does not appear to play a significant role in the ability for a riparian corridor to rebound in the year following fire. Riparian vegetation NDVI rebounded in the year following the fire regardless of proximity to beaver activity. Thus, we would describe beaver activity as creating refugia during wildfire, but not necessarily changing the long-term landscape outcomes [Fairfax and Whittle 2020: 7].
The survival of wildlife is crucial to these ecosystems, and beaver activity uniquely contributes to the creation of refuge areas that resist burning and can provide shelter for animals during these destructive events. The authors conclude,
As it stands today, wetland habitat is very limited and beavers can create and maintain wetland habitat that persists through flood, drought, and, as we have shown in this study, fire. This has immediate relevance to scientists and practitioners across North America and Eurasia, particularly in places with increasing wildfire risk and existing or planned beaver populations. Perhaps instead of relying solely on human engineering and management to create and maintain fire-resistant landscape patches, we could benefit from beavers’ ecosystem engineering to achieve the same goals at a lower cost [Fairfax and Whittle 2020: 7].
It has long been suspected that the increasing abundance of invasive grass species may contribute to wildfires in the United States by adding abundant new fuels to ecosystems, increasing the range of conditions that lead to fire ignition, and enabling the development of larger, hotter fires. The new fire regimes (patterns of fire duration, intensity, and spread) that emerge can in turn destabilize wildlife and lead to local extinctions while expanding favorable habitat for the invasive species, for many of these grasses recover quickly after fires, providing renewed fuel and potentially increasing the frequency of fires.
The authors of this paper provide a comprehensive analysis on the impact of 12 non-native grasses on the occurrence (whether a fire occurred in a particular place), frequency (how many times a place burned), and size of wildfires. The research was conducted across 29 US ecoregions, including deserts, temperate forests, wetlands, woodlands, river valleys, shrublands, and coastal plains. Data were collected and combined from fire records and records of invasive grasses, and results from “invaded” regions and nearby “uninvaded” regions were compared. The authors also considered human activities and ecological factors related to fire.
One of the most notorious impacts of nonnative, invasive grasses is the alteration of fire regimes. Yet, most evidence of these impacts comes from local-scale studies, making it unclear whether they have broader implications for national and regional fire management. Our analysis of 12 invasive grasses documents regional-scale alteration of fire regimes for 8 species, which are already increasing fire occurrence by up to 230% and fire frequency by up to 150%. These impacts were demonstrated across US ecoregions and vegetation types, suggesting that many ecosystems are vulnerable to a novel grass-fire cycle. Managing existing grass invasions and preventing future introductions presents a key opportunity to remediate the ecological and economic consequences of invasive species and fire [Fusco 2019: 23594]
The results of this analysis showed that 8 of the 12 invasive grass species examined were associated with significantly higher fire occurrence and fire frequency, and that fire occurrence more than doubled for two of these species. Three of the species did not impact fire occurrence, and a decrease in fires was associated with one species (a wetlands grass species). The impact on fire size was variable, with two species associated with larger fires, three species associated with smaller fires.
Individually, climate change is expected to increase the potential for fire occurrence by 150% by the end of the century based on projected changes in temperature and precipitation. Here we show that 8 invasive grass species are already associated with increased rates of fire occurrence by 27 to 230%, and 6 invasive grass species are associated with increased mean fire frequency by 24 to 150%, compounding current and future fire risk across the United States. [Fusco et al. 2019: 23595]
The authors suggest that fire and invasive species managers work together to create integrated management plans; otherwise, the convergence of human activities, climate change, and invasive species will continue to promote wildfires across the United States.
Indigenous groups across the world have developed ecological knowledge linked to the places they inhabit, including prescribed fire practices used to maintain healthy ecosystems. Mistry et al. examine the challenges Indigenous communities in South America face in managing the landscape through fire and preserving such knowledge across generations in sometimes hostile political climates. However, there is growing recognition that Indigenous people have a vital role to play in combating climate change and supporting biodiversity and healthy ecosystems.
Emerging research shows the fundamental role of Indigenous land-use practices for controlling deforestation and reducing CO2 emissions—analysis of satellite imagery suggests that Indigenous lands have reduced rates of deforestation and habitat conversion, and lower greenhouse gas (GHG) emissions, compared with surrounding areas [Mistry 2016: 1].
While indigenous groups’ use of prescribed fire early in the dry season to prevent destructive out-of-control fires is gaining broad recognition, that hasn’t necessarily translated into greater respect or autonomy for those communities. Instead, Indigenous people may be given auxiliary roles in fire management, or have their knowledge utilized but implemented by non-local organizations in structures that fail to benefit or empower the local communities themselves. While this may still achieve desired wildfire management results, it weakens intergenerational knowledge transfer and undermines the social and spiritual role of prescribed fire within communities.
Mistry et al. argue that “Indigenous fire management is effective in that it is an emergent property of a linked social-ecological system where Indigenous knowledge and culture, and associated livelihoods, are intimately interconnected with landscape management practices” [Mistry 2016: 4]. Precisely because prescribed fire matters to Indigenous communities as something more than a tool in the toolkit of managing wildfires, it is effective when carried out by those communities in reducing risk of destructive wildfires and supporting healthy and biodiverse ecosystems.
Importantly, the numerous uses of fire mean that burning is a relatively constant activity, particularly during the dry season, generally at low levels, thereby helping to prevent the build-up of flammable fuel and incidents of large-scale uncontrollable wildfires. Experimental studies of fire behaviour suggest that this patch mosaic burning not only reduces the occurrence of dangerous fires, but also increases spatial and temporal vegetation heterogeneity and biodiversity [Mistry 2016: 4].
These authors distinguish between Indigenous relationships to ecosystems and market-based approaches to ecosystem services valuation, which attempt to incentivize conservation through payment. While the goal of the market-based approach is to monitor and preserve functioning ecosystems, “their ideological foundations within a neoliberal agenda that promotes ‘selling nature to save it’ is in stark contradiction with Indigenous ontologies based on human–nonhuman–spiritual relationships” [Mistry 2016: 2].
Within Indigenous communities, fire plays a role in social bonding, intergenerational knowledge transfer, and agricultural practices. Mistry et al. argue that
savanna and forest ecosystems are being protected within Indigenous lands not because they are being ‘managed’ in a direct and active way, but as the indirect outcome of a healthy social–ecological system, i.e. the outcome of practices that maintain social and ecological integrity, or what can be termed ‘community owned solutions [Mistry 2016: 4].
But challenges, including loss of fire knowledge by younger generations within Indigenous groups because of outside pressures and encroachment, pose a threat to these fire management practices. For example, in Venezuela and Brazil,
young Wapishana and Makushi and some community leaders were more critical about the use of fire as they had more regular contact with state natural resource management officials and environmental organizations that promoted antifire discourses. As with the Krahô, changing Indigenous values to focus on fire prevention and suppression could have the effect of making the problem worse [Mistry 2016: 4].
That is, when prescribed burning is taken out of its original context and represented to younger generations of Indigenous people and land stewards as simply a well-incentivized tool, the Indigenous communities themselves are diminished, along with the robustness of their ecological knowledge that gets passed forward.
In spite of lingering antagonistic views in Brazil and Venezuela toward indigenous fire management, attitudes are changing.
Not only is there a move away from categorizing all fire as ‘bad’; there is also a recognition that Indigenous fire knowledge is a valid form of knowledge that could inform policy-making [Mistry et al. 2016: 6].
Mistry et al. suggest the best way to achieve both ecological and communal health might be through power-sharing arrangements. By empowering Indigenous communities, national governments could in turn work toward their fire management and biodiversity conservation goals. This might require evaluating ecological health in ways beyond just quantitative metrics, which reduce these complex systems down to a set of standardized numbers, as well as the recognition that the well-being of these ecosystems is tied to the Indigenous communities that inhabit them, according to the authors:
There needs to be enabling policies that focus on legitimizing and strengthening Indigenous fire management as a community owned solution. Critically, as community owned fire management is intricately linked with Indigenous survival strategies, so too must firefighting and prescribed burning be grounded in local social–ecological systems. We believe it is necessary to define long-term actions to support the integrated functioning and survival of Indigenous communities as a whole, rather than focusing on isolated issues (e.g. carbon retention) or benefits for some individuals (e.g. hiring Indigenous firefighters) [Mistry 2016: 8].
This systems approach may well be the key to successful long-term fire management. The authors offer this challenge:
What we want to do is not promote one over the other, but encourage decision-makers to engage with, and appreciate, Indigenous perspectives and worldviews on fire management. Community owned solutions acknowledge collectivity, spirituality, process orientation and locality, whereas many expert-led fire management interventions often result in promoting individualism, ethnocentrism, rationality, efficiency, commercialism and globalization. The question we raise is this: can the ‘community owned solutions’ approach be the mechanism through which Indigenous perspectives can be represented within fire management [Mistry et al. 2016: 8]?
Wildfire is a challenge that threatens human settlement at an increasing scale, but planning and development does not always address this threat. In fact, policy around land use is in large part responsible for the destruction of homes and property and the threat to human life that occurs in wildland-urban interfaces (WUIs). While there is much literature on how to suppress fires, mitigate their damage, or manage for less destructive fire seasons, a more far-reaching strategy would be to stop building in fire prone areas. Land use decisions can be improved to lessen the risk of infrastructure loss and foster healthy ecosystem function.
Land use planning is an alternative that represents a further shift in thinking, beyond the preparation of communities to withstand an inevitable fire, to preventing new residential structures from being exposed to fire in the first place. The reason homes are vulnerable to fires at the wildland-urban interface is a function of its very definition: “where homes meet or intermingle with wildland vegetation”. In other words, the location and pattern of homes influence their fire risk, and past land-use decision-making has allowed homes to be constructed in highly flammable areas. [Syphard 2013: 1-2]
In many areas, including in California, we have come to expect fire, but have not necessarily learned to live with it. The authors of this study analyzed what types of human development carried out in the next several years might contribute to or avert the risk of fire damage. They examined the South Coast Ecoregion of San Diego County, which they describe as:
topographically diverse with high levels of biodiversity, and urban development has been the primary cause of natural habitat loss and species extinction. Owing to the Mediterranean climate, with mild, wet winters and long summer droughts, the native shrublands dominating the landscape are extremely fire-prone [Syphard et al. 2013: 2].
This study acknowledges the responsibility humans have in shaping the landscape and its biodiversity, and in contributing to fire activity by building into wild areas and expanding WUI. They sought to understand how patterns of development and housing density might influence future fire spread and intensity. They found that
structures in areas with low- to intermediate- housing density were most likely to burn, potentially due to intermingling with wildland vegetation or difficulty of firefighter access. Fire frequency also tends to be highest at low to intermediate housing density, at least in regions where humans are the primary cause of ignitions [Syphard 2013: 2].
Though it is impossible to reverse the effects of policies that have shaped the fire landscape we have today, understanding the way human behavior contributes to our own risk of harm from wildfire can help us plan intelligently going forward. The authors conclude that
With projections of substantial global change in climate and human development, we recommend that land use planning should be considered as an important component to fire risk management, potentially to become as successful as the prevention of building on flood plains. History has shown us that preventing fires is impossible in areas where large wildfires are a natural ecological process. As Roger Kennedy put it, “the problem isn’t fires; the problem is people in the wrong places [Syphard 2013: 10-11].
In this podcast interview, Dr. Chad Hanson, an ecologist and fire researcher, shares his perspective on the 2018 wildfires in the American West and some myths that have circulated about fire management in their wake.
First, there is a perception that wildfires in forested regions are so devastating that they reverse the ‘carbon sink’ effect of forests, releasing the carbon of the burned biomass back into the atmosphere. Forests still sequester large amounts of carbon, even if they experience wildfires, because most of the forest remains intact even through blazes. Models that fault wildfires for turning forests into net carbon emitters rest on the assumption that all of the carbon that would usually be stored in a forest is combusted during a fire, but this is far from the reality, in which just a small fraction of a forest’s biomass is consumed. As Hanson says,
In fact, even in the most intensely burned patches where a fire kills all the trees (which in reality, even in the biggest fires, it’s only a small portion, a minor portion of the overall fire)… But even in those areas, only about two or three percent of the above ground biomass is actually consumed, in other words, ends up as carbon. The trees are still standing there [Hanson 2018].
Second, the intensity of fires has not been universally increasing in recent years. We are experiencing a lot of geographically large fires, but these are not necessarily high intensity fires. The percentage of high intensity fire today is similar to historical precedent, and overall, there is much less fire in our landscapes now than in Earth’s geological history. Further, even in the highest intensity fires, forests are never decimated past the point of no return. Trees and vegetation are reestablished after fires, and the ash left behind is dense with nutrients, promoting new growth. Even dead trees, which have long been thought to be responsible for contributing to high intensity fires, are actually not shown to drive fire intensity, according to Hanson.
This series of myths – that forest fires are raging with high intensities, that they burn up so much biomass as to make forests ineffective sources of carbon sequestration, and that the only way to manage forests to avoid these outcomes is to thin out the trees – hinder our understanding of forest management and allow false and harmful solutions to propagate. As a result of these perceptions, proponents of logging have pushed to expand logging operations, purportedly as a fire management strategy. However, according to Hanson, logging is actually linked to greater fire intensity. He explains that small materials, like twigs, are more flammable than trunks.
Tree trunks are not combustible. They really just don’t burn. Again, outer bark can burn, but the trees themselves don’t burn. What logging does is it removes noncombustible material essentially from the forest and leaves behind very combustible kindling, like slash debris – the branches and small twigs and things like that that are not possible to get up off the forest floor after the tree trunks are removed and that’s very combustible.
The other thing that logging does is that it reduces the cooling shade of the forest canopy. By removing a lot of trees, you have more sunlight reaching the forest floor, and what that does is it creates hotter and drier conditions and that means everything on the forest floor gets more dried out, more potentially combustible, and logging also spreads invasive weeds like cheatgrass, which is very, very flammable. Cheatgrass loves a lot of sunlight and so you get a lot of that after intensive logging.
And the last [problem with logging] is a little bit more technical, but basically when you have a lot more trees, it cuts down on the wind speeds that drive fires. It has a buffering effect in a sense. And when a lot of the trees are removed, that buffering effect is reduced or eliminated and fire spreads through those forests faster [Hanson 2018].
This three-fold effect of logging makes forests more vulnerable, and it is important to dispel the concept that removing trees is the best way to keep people safe from fires. Hanson criticizes the opportunism of using these fire myths to advance an agenda of logging. He cautions that when fire science and policy emerge from the U.S. Forest Service, which manages national forests and gains a good deal of revenue from logging, there is a perverse incentive to keep practicing logging as fire management. He calls for clearer and more public communication from scientists to dispel fire myths and share recent findings that have been shifting so much of what we know about fire science.
Hanson says that the best strategy to ensure the protection of homes from wildfire is to focus on the homes themselves. This can be done by using fire resistant building materials, fire-proofing roofs, erecting rain gutter guards to prevent the accumulation of small fuels like pine needles, pruning the vegetation in a 100-foot radius of a house, and removing small shrubs and branches of mature trees, while leaving those trees standing. Fire management interventions at this level are shown to be far more effective at preventing damage than attempts to control the fuel load of fires within forests.
Hanson points out the need to decouple fires that occur in remote forest ecosystems and those that rage through human settlements and urban communities, because thinning out vegetation in attempts to suppress the former do not actually protect against the latter. In fact, thinning forests undermines the ecological processes that fire serves in forest systems. When discussing fires that have devastated homes and lives, he says
I mean, where’s the forest in Malibu? There’s no forest. These are chaparral ecosystems, most of the fires that are burned homes and lives have been lost are not in forest. In fact, they’re mostly nowhere near forests. They’re in grasslands, chaparral shrub habitat, oak woodlands. But the areas that are in forest, where we’ve had tragic loss of homes and lives, these are mostly areas where we’ve had intensive logging, and it’s like I mentioned earlier, you know, more logging is typically associated with more intense fire at a faster rate of spread. [Hanson 2018]
He advocates for a greater focus on fire prevention around homes and communities themselves, in what is known as ‘defensible space.’ He points out that such measures are a great source of jobs, as well as an effective intervention curbing the destructiveness of fires. With a shift in focus from forest thinning to fire-proofing, and better understanding and communication of fire science, we can let go of some of the fire myths that have been dictating policy and failing to meet public needs.
More logging is typically associated with more intense fire at a faster rate of spread [Hanson 2018].
Steven J. Pyne is an emeritus professor at Arizona State University and the author of several books on fire history and policy. He wrote this opinion piece as a protest against the prevention and suppression of wildfires in our land management process. He argues that revising our perception of fire and accepting its presence in ecosystems is critical to our ongoing relationship with our planet.
He describes “a paradox at the core of Earth’s unraveling firescapes,” that “we have too many bad fires — fires that kill people, burn towns, and trash valued landscapes. We have too few good ones — fires that enhance ecological integrity and hold fires within their historic ranges” [Pyne 2020]. Operating under a paradigm of total fire suppression leads us astray in managing landscapes, while we so readily accept fire in the form of fossil fuel combustion in so much of our lives. Pyne sees these behaviors as evidence that our relationship with fire is out of whack.
He stresses the importance of distinguishing between burning in living ecosystems and burning the fossils of life from past ages.
The critical contrast lies in a deeper dialectic than burned and unburned landscapes. It is a dialectic between burning living biomass and burning fossil biomass. We are taking stuff out of the geologic past, burning it in the present with all kinds of little understood consequences, and passing the effluent into the geologic future.
…
Fires in living landscapes come with ecological checks and balances. Fires in lithic landscapes have no boundaries save those humans impose on themselves [Pyne 2020].
Pyne associates three paradoxes with our current fire policies. First, abandoning a traditional lore of “light burning” has removed good fires and left us with only bad and harmful ones. When controlled burns are not practiced regularly to manage landscapes, vegetation can build up and fuel the intensity and spread of uncontrolled blazes that spark.
Second and surprisingly: “The Earth does not have more fire today than before fossil fuels emerged as a primary source of energy: It has significantly less” [Pyne 2020]. That is, the amount of land burned in fires has actually decreased, while the presence of intense “feral flames” has increased. The decrease in the scope of fire is largely due to the move away from fire’s use in agriculture and its replacement with modern techniques, including machinery powered by combustion. As Pyne describes,
Farmers had relied on fire to fertilize, fumigate, and alter microclimates. Fire did all this in one catalytic process that self-propagated. But with the transition to fossil biomass, modern agriculture found surrogates with artificial fertilizers, pesticides, and herbicides, and it now had the fossil-fuel-powered machines to distribute them. Production became more efficient; transport, more dense. As agriculture joins a modern economy, working flames recede [Pyne 2020].
What we are left with is intense, destructive wildfire, rather than helpful working fire. Pyne points out that we now only see one half of fire’s possibilities, since the working fire shaping landscapes and agricultural systems is notably absent. He says: “Landscape fire fades; what fire persists tends to be outbreaks of feral fire. We see those oft-disastrous flames. We don’t see the lost fires or the sublimated fires in machines that removed them” [Pyne 2020].
The third paradox is that as we reduce our use of fossil fuels going forward (as one sort of fire), we will have a greater need “to manage fire in living landscapes.” So Pyne calls for us “to reinstate the right kind of fire, and … adapt to fire’s presence and let it do the work for us.” He pushes for the recognition that transitioning away from our reliance on fossil fuel burning is an important but incomplete step in balancing ecosystem health. Fires in living systems have an important role to play, and according to Pyne, “the need is not just to reduce fuels to help contain wildfires; those missing fires did biological work for which no single surrogate exists” [Pyne 2020]. He asserts that we will need to reintroduce fire as a staple tool on our landscapes.
Fires in living systems have an important role to play, and according to Pyne, “the need is not just to reduce fuels to help contain wildfires; those missing fires did biological work for which no single surrogate exists.” He asserts that we will need to reintroduce fire as a staple tool on our landscapes.
Pyne concludes with a call for the overhaul of our conceptual and policy treatment of fire.
Anthropogenic fire needs more room to maneuver – more geographic space, more legal space, more political space, more conceptual space. … Equally, society needs to rethink liability law to reduce the risks incurred by fire officers doing a necessary job …; adapt air quality regulations …; and tweak National Environmental Policy Act review processes … [to] accommodate the realities of restored fire at a landscape scale. … Communities in the fire equivalents of floodplains need hardening [Pyne 2020].
He proposes that fire restoration jobs can replace those lost from forestry and fire suppression. He admits that our understanding of fire biology requires more research, and that our greatest need is for “a working fire culture … that ensures fire’s proper place in the landscape” by renewing “our ancient alliance” with fire and making it “an indispensable friend.”
Our burning planet: why we must learn to live with fire, Pyne 2020
Steven J. Pyne is an emeritus professor at Arizona State University and the author of several books on fire history and policy. He wrote this opinion piece as a protest against the prevention and suppression of wildfires in our land management process. He argues that revising our perception of fire and accepting its presence in ecosystems is critical to our ongoing relationship with our planet.
He describes “a paradox at the core of Earth’s unraveling firescapes,” that “we have too many bad fires — fires that kill people, burn towns, and trash valued landscapes. We have too few good ones — fires that enhance ecological integrity and hold fires within their historic ranges” [Pyne 2020]. Operating under a paradigm of total fire suppression leads us astray in managing landscapes, while we so readily accept fire in the form of fossil fuel combustion in so much of our lives. Pyne sees these behaviors as evidence that our relationship with fire is out of whack.
He stresses the importance of distinguishing between burning in living ecosystems and burning the fossils of life from past ages.
The critical contrast lies in a deeper dialectic than burned and unburned landscapes. It is a dialectic between burning living biomass and burning fossil biomass. We are taking stuff out of the geologic past, burning it in the present with all kinds of little understood consequences, and passing the effluent into the geologic future.
…
Fires in living landscapes come with ecological checks and balances. Fires in lithic landscapes have no boundaries save those humans impose on themselves [Pyne 2020].
Pyne associates three paradoxes with our current fire policies. First, abandoning a traditional lore of “light burning” has removed good fires and left us with only bad and harmful ones. When controlled burns are not practiced regularly to manage landscapes, vegetation can build up and fuel the intensity and spread of uncontrolled blazes that spark.
Second and surprisingly: “The Earth does not have more fire today than before fossil fuels emerged as a primary source of energy: It has significantly less” [Pyne 2020]. That is, the amount of land burned in fires has actually decreased, while the presence of intense “feral flames” has increased. The decrease in the scope of fire is largely due to the move away from fire’s use in agriculture and its replacement with modern techniques, including machinery powered by combustion. As Pyne describes,
Farmers had relied on fire to fertilize, fumigate, and alter microclimates. Fire did all this in one catalytic process that self-propagated. But with the transition to fossil biomass, modern agriculture found surrogates with artificial fertilizers, pesticides, and herbicides, and it now had the fossil-fuel-powered machines to distribute them. Production became more efficient; transport, more dense. As agriculture joins a modern economy, working flames recede [Pyne 2020].
What we are left with is intense, destructive wildfire, rather than helpful working fire. Pyne points out that we now only see one half of fire’s possibilities, since the working fire shaping landscapes and agricultural systems is notably absent. He says: “Landscape fire fades; what fire persists tends to be outbreaks of feral fire. We see those oft-disastrous flames. We don’t see the lost fires or the sublimated fires in machines that removed them” [Pyne 2020].
The third paradox is that as we reduce our use of fossil fuels going forward (as one sort of fire), we will have a greater need “to manage fire in living landscapes.” So Pyne calls for us “to reinstate the right kind of fire, and … adapt to fire’s presence and let it do the work for us.” He pushes for the recognition that transitioning away from our reliance on fossil fuel burning is an important but incomplete step in balancing ecosystem health. Fires in living systems have an important role to play, and according to Pyne, “the need is not just to reduce fuels to help contain wildfires; those missing fires did biological work for which no single surrogate exists” [Pyne 2020]. He asserts that we will need to reintroduce fire as a staple tool on our landscapes.
Fires in living systems have an important role to play, and according to Pyne, “the need is not just to reduce fuels to help contain wildfires; those missing fires did biological work for which no single surrogate exists.” He asserts that we will need to reintroduce fire as a staple tool on our landscapes.
Pyne concludes with a call for the overhaul of our conceptual and policy treatment of fire.
Anthropogenic fire needs more room to maneuver – more geographic space, more legal space, more political space, more conceptual space. … Equally, society needs to rethink liability law to reduce the risks incurred by fire officers doing a necessary job …; adapt air quality regulations …; and tweak National Environmental Policy Act review processes … [to] accommodate the realities of restored fire at a landscape scale. … Communities in the fire equivalents of floodplains need hardening [Pyne 2020].
He proposes that fire restoration jobs can replace those lost from forestry and fire suppression. He admits that our understanding of fire biology requires more research, and that our greatest need is for “a working fire culture … that ensures fire’s proper place in the landscape” by renewing “our ancient alliance” with fire and making it “an indispensable friend.”
Fire Myths, Hanson 2018
In this podcast interview, Dr. Chad Hanson, an ecologist and fire researcher, shares his perspective on the 2018 wildfires in the American West and some myths that have circulated about fire management in their wake.
First, there is a perception that wildfires in forested regions are so devastating that they reverse the ‘carbon sink’ effect of forests, releasing the carbon of the burned biomass back into the atmosphere. Forests still sequester large amounts of carbon, even if they experience wildfires, because most of the forest remains intact even through blazes. Models that fault wildfires for turning forests into net carbon emitters rest on the assumption that all of the carbon that would usually be stored in a forest is combusted during a fire, but this is far from the reality, in which just a small fraction of a forest’s biomass is consumed. As Hanson says,
In fact, even in the most intensely burned patches where a fire kills all the trees (which in reality, even in the biggest fires, it’s only a small portion, a minor portion of the overall fire)… But even in those areas, only about two or three percent of the above ground biomass is actually consumed, in other words, ends up as carbon. The trees are still standing there [Hanson 2018].
Second, the intensity of fires has not been universally increasing in recent years. We are experiencing a lot of geographically large fires, but these are not necessarily high intensity fires. The percentage of high intensity fire today is similar to historical precedent, and overall, there is much less fire in our landscapes now than in Earth’s geological history. Further, even in the highest intensity fires, forests are never decimated past the point of no return. Trees and vegetation are reestablished after fires, and the ash left behind is dense with nutrients, promoting new growth. Even dead trees, which have long been thought to be responsible for contributing to high intensity fires, are actually not shown to drive fire intensity, according to Hanson.
This series of myths – that forest fires are raging with high intensities, that they burn up so much biomass as to make forests ineffective sources of carbon sequestration, and that the only way to manage forests to avoid these outcomes is to thin out the trees – hinder our understanding of forest management and allow false and harmful solutions to propagate. As a result of these perceptions, proponents of logging have pushed to expand logging operations, purportedly as a fire management strategy. However, according to Hanson, logging is actually linked to greater fire intensity. He explains that small materials, like twigs, are more flammable than trunks.
Tree trunks are not combustible. They really just don’t burn. Again, outer bark can burn, but the trees themselves don’t burn. What logging does is it removes noncombustible material essentially from the forest and leaves behind very combustible kindling, like slash debris – the branches and small twigs and things like that that are not possible to get up off the forest floor after the tree trunks are removed and that’s very combustible.
The other thing that logging does is that it reduces the cooling shade of the forest canopy. By removing a lot of trees, you have more sunlight reaching the forest floor, and what that does is it creates hotter and drier conditions and that means everything on the forest floor gets more dried out, more potentially combustible, and logging also spreads invasive weeds like cheatgrass, which is very, very flammable. Cheatgrass loves a lot of sunlight and so you get a lot of that after intensive logging.
And the last [problem with logging] is a little bit more technical, but basically when you have a lot more trees, it cuts down on the wind speeds that drive fires. It has a buffering effect in a sense. And when a lot of the trees are removed, that buffering effect is reduced or eliminated and fire spreads through those forests faster [Hanson 2018].
This three-fold effect of logging makes forests more vulnerable, and it is important to dispel the concept that removing trees is the best way to keep people safe from fires. Hanson criticizes the opportunism of using these fire myths to advance an agenda of logging. He cautions that when fire science and policy emerge from the U.S. Forest Service, which manages national forests and gains a good deal of revenue from logging, there is a perverse incentive to keep practicing logging as fire management. He calls for clearer and more public communication from scientists to dispel fire myths and share recent findings that have been shifting so much of what we know about fire science.
Hanson says that the best strategy to ensure the protection of homes from wildfire is to focus on the homes themselves. This can be done by using fire resistant building materials, fire-proofing roofs, erecting rain gutter guards to prevent the accumulation of small fuels like pine needles, pruning the vegetation in a 100-foot radius of a house, and removing small shrubs and branches of mature trees, while leaving those trees standing. Fire management interventions at this level are shown to be far more effective at preventing damage than attempts to control the fuel load of fires within forests.
Hanson points out the need to decouple fires that occur in remote forest ecosystems and those that rage through human settlements and urban communities, because thinning out vegetation in attempts to suppress the former do not actually protect against the latter. In fact, thinning forests undermines the ecological processes that fire serves in forest systems. When discussing fires that have devastated homes and lives, he says
I mean, where’s the forest in Malibu? There’s no forest. These are chaparral ecosystems, most of the fires that are burned homes and lives have been lost are not in forest. In fact, they’re mostly nowhere near forests. They’re in grasslands, chaparral shrub habitat, oak woodlands. But the areas that are in forest, where we’ve had tragic loss of homes and lives, these are mostly areas where we’ve had intensive logging, and it’s like I mentioned earlier, you know, more logging is typically associated with more intense fire at a faster rate of spread. [Hanson 2018]
He advocates for a greater focus on fire prevention around homes and communities themselves, in what is known as ‘defensible space.’ He points out that such measures are a great source of jobs, as well as an effective intervention curbing the destructiveness of fires. With a shift in focus from forest thinning to fire-proofing, and better understanding and communication of fire science, we can let go of some of the fire myths that have been dictating policy and failing to meet public needs.
More logging is typically associated with more intense fire at a faster rate of spread [Hanson 2018].
Land use planning and wildfire: development policies influence future probability of housing loss, Syphard et al. 2013
Wildfire is a challenge that threatens human settlement at an increasing scale, but planning and development does not always address this threat. In fact, policy around land use is in large part responsible for the destruction of homes and property and the threat to human life that occurs in wildland-urban interfaces (WUIs). While there is much literature on how to suppress fires, mitigate their damage, or manage for less destructive fire seasons, a more far-reaching strategy would be to stop building in fire prone areas. Land use decisions can be improved to lessen the risk of infrastructure loss and foster healthy ecosystem function.
Land use planning is an alternative that represents a further shift in thinking, beyond the preparation of communities to withstand an inevitable fire, to preventing new residential structures from being exposed to fire in the first place. The reason homes are vulnerable to fires at the wildland-urban interface is a function of its very definition: “where homes meet or intermingle with wildland vegetation”. In other words, the location and pattern of homes influence their fire risk, and past land-use decision-making has allowed homes to be constructed in highly flammable areas. [Syphard 2013: 1-2]
In many areas, including in California, we have come to expect fire, but have not necessarily learned to live with it. The authors of this study analyzed what types of human development carried out in the next several years might contribute to or avert the risk of fire damage. They examined the South Coast Ecoregion of San Diego County, which they describe as:
topographically diverse with high levels of biodiversity, and urban development has been the primary cause of natural habitat loss and species extinction. Owing to the Mediterranean climate, with mild, wet winters and long summer droughts, the native shrublands dominating the landscape are extremely fire-prone [Syphard et al. 2013: 2].
This study acknowledges the responsibility humans have in shaping the landscape and its biodiversity, and in contributing to fire activity by building into wild areas and expanding WUI. They sought to understand how patterns of development and housing density might influence future fire spread and intensity. They found that
structures in areas with low- to intermediate- housing density were most likely to burn, potentially due to intermingling with wildland vegetation or difficulty of firefighter access. Fire frequency also tends to be highest at low to intermediate housing density, at least in regions where humans are the primary cause of ignitions [Syphard 2013: 2].
Though it is impossible to reverse the effects of policies that have shaped the fire landscape we have today, understanding the way human behavior contributes to our own risk of harm from wildfire can help us plan intelligently going forward. The authors conclude that
With projections of substantial global change in climate and human development, we recommend that land use planning should be considered as an important component to fire risk management, potentially to become as successful as the prevention of building on flood plains. History has shown us that preventing fires is impossible in areas where large wildfires are a natural ecological process. As Roger Kennedy put it, “the problem isn’t fires; the problem is people in the wrong places [Syphard 2013: 10-11].
Community owned solutions for fire management in tropical ecosystems: case studies from Indigenous communities of South America, Mistry et al. 2016
Indigenous groups across the world have developed ecological knowledge linked to the places they inhabit, including prescribed fire practices used to maintain healthy ecosystems. Mistry et al. examine the challenges Indigenous communities in South America face in managing the landscape through fire and preserving such knowledge across generations in sometimes hostile political climates. However, there is growing recognition that Indigenous people have a vital role to play in combating climate change and supporting biodiversity and healthy ecosystems.
Emerging research shows the fundamental role of Indigenous land-use practices for controlling deforestation and reducing CO2 emissions—analysis of satellite imagery suggests that Indigenous lands have reduced rates of deforestation and habitat conversion, and lower greenhouse gas (GHG) emissions, compared with surrounding areas [Mistry 2016: 1].
While indigenous groups’ use of prescribed fire early in the dry season to prevent destructive out-of-control fires is gaining broad recognition, that hasn’t necessarily translated into greater respect or autonomy for those communities. Instead, Indigenous people may be given auxiliary roles in fire management, or have their knowledge utilized but implemented by non-local organizations in structures that fail to benefit or empower the local communities themselves. While this may still achieve desired wildfire management results, it weakens intergenerational knowledge transfer and undermines the social and spiritual role of prescribed fire within communities.
Mistry et al. argue that “Indigenous fire management is effective in that it is an emergent property of a linked social-ecological system where Indigenous knowledge and culture, and associated livelihoods, are intimately interconnected with landscape management practices” [Mistry 2016: 4]. Precisely because prescribed fire matters to Indigenous communities as something more than a tool in the toolkit of managing wildfires, it is effective when carried out by those communities in reducing risk of destructive wildfires and supporting healthy and biodiverse ecosystems.
Importantly, the numerous uses of fire mean that burning is a relatively constant activity, particularly during the dry season, generally at low levels, thereby helping to prevent the build-up of flammable fuel and incidents of large-scale uncontrollable wildfires. Experimental studies of fire behaviour suggest that this patch mosaic burning not only reduces the occurrence of dangerous fires, but also increases spatial and temporal vegetation heterogeneity and biodiversity [Mistry 2016: 4].
These authors distinguish between Indigenous relationships to ecosystems and market-based approaches to ecosystem services valuation, which attempt to incentivize conservation through payment. While the goal of the market-based approach is to monitor and preserve functioning ecosystems, “their ideological foundations within a neoliberal agenda that promotes ‘selling nature to save it’ is in stark contradiction with Indigenous ontologies based on human–nonhuman–spiritual relationships” [Mistry 2016: 2].
Within Indigenous communities, fire plays a role in social bonding, intergenerational knowledge transfer, and agricultural practices. Mistry et al. argue that
savanna and forest ecosystems are being protected within Indigenous lands not because they are being ‘managed’ in a direct and active way, but as the indirect outcome of a healthy social–ecological system, i.e. the outcome of practices that maintain social and ecological integrity, or what can be termed ‘community owned solutions [Mistry 2016: 4].
But challenges, including loss of fire knowledge by younger generations within Indigenous groups because of outside pressures and encroachment, pose a threat to these fire management practices. For example, in Venezuela and Brazil,
young Wapishana and Makushi and some community leaders were more critical about the use of fire as they had more regular contact with state natural resource management officials and environmental organizations that promoted antifire discourses. As with the Krahô, changing Indigenous values to focus on fire prevention and suppression could have the effect of making the problem worse [Mistry 2016: 4].
That is, when prescribed burning is taken out of its original context and represented to younger generations of Indigenous people and land stewards as simply a well-incentivized tool, the Indigenous communities themselves are diminished, along with the robustness of their ecological knowledge that gets passed forward.
In spite of lingering antagonistic views in Brazil and Venezuela toward indigenous fire management, attitudes are changing.
Not only is there a move away from categorizing all fire as ‘bad’; there is also a recognition that Indigenous fire knowledge is a valid form of knowledge that could inform policy-making [Mistry et al. 2016: 6].
Mistry et al. suggest the best way to achieve both ecological and communal health might be through power-sharing arrangements. By empowering Indigenous communities, national governments could in turn work toward their fire management and biodiversity conservation goals. This might require evaluating ecological health in ways beyond just quantitative metrics, which reduce these complex systems down to a set of standardized numbers, as well as the recognition that the well-being of these ecosystems is tied to the Indigenous communities that inhabit them, according to the authors:
There needs to be enabling policies that focus on legitimizing and strengthening Indigenous fire management as a community owned solution. Critically, as community owned fire management is intricately linked with Indigenous survival strategies, so too must firefighting and prescribed burning be grounded in local social–ecological systems. We believe it is necessary to define long-term actions to support the integrated functioning and survival of Indigenous communities as a whole, rather than focusing on isolated issues (e.g. carbon retention) or benefits for some individuals (e.g. hiring Indigenous firefighters) [Mistry 2016: 8].
This systems approach may well be the key to successful long-term fire management. The authors offer this challenge:
What we want to do is not promote one over the other, but encourage decision-makers to engage with, and appreciate, Indigenous perspectives and worldviews on fire management. Community owned solutions acknowledge collectivity, spirituality, process orientation and locality, whereas many expert-led fire management interventions often result in promoting individualism, ethnocentrism, rationality, efficiency, commercialism and globalization. The question we raise is this: can the ‘community owned solutions’ approach be the mechanism through which Indigenous perspectives can be represented within fire management [Mistry et al. 2016: 8]?
Invasive grasses increase fire occurrence and frequency across US ecoregions, Fusco et al. 2019
It has long been suspected that the increasing abundance of invasive grass species may contribute to wildfires in the United States by adding abundant new fuels to ecosystems, increasing the range of conditions that lead to fire ignition, and enabling the development of larger, hotter fires. The new fire regimes (patterns of fire duration, intensity, and spread) that emerge can in turn destabilize wildlife and lead to local extinctions while expanding favorable habitat for the invasive species, for many of these grasses recover quickly after fires, providing renewed fuel and potentially increasing the frequency of fires.
The authors of this paper provide a comprehensive analysis on the impact of 12 non-native grasses on the occurrence (whether a fire occurred in a particular place), frequency (how many times a place burned), and size of wildfires. The research was conducted across 29 US ecoregions, including deserts, temperate forests, wetlands, woodlands, river valleys, shrublands, and coastal plains. Data were collected and combined from fire records and records of invasive grasses, and results from “invaded” regions and nearby “uninvaded” regions were compared. The authors also considered human activities and ecological factors related to fire.
One of the most notorious impacts of nonnative, invasive grasses is the alteration of fire regimes. Yet, most evidence of these impacts comes from local-scale studies, making it unclear whether they have broader implications for national and regional fire management. Our analysis of 12 invasive grasses documents regional-scale alteration of fire regimes for 8 species, which are already increasing fire occurrence by up to 230% and fire frequency by up to 150%. These impacts were demonstrated across US ecoregions and vegetation types, suggesting that many ecosystems are vulnerable to a novel grass-fire cycle. Managing existing grass invasions and preventing future introductions presents a key opportunity to remediate the ecological and economic consequences of invasive species and fire [Fusco 2019: 23594]
The results of this analysis showed that 8 of the 12 invasive grass species examined were associated with significantly higher fire occurrence and fire frequency, and that fire occurrence more than doubled for two of these species. Three of the species did not impact fire occurrence, and a decrease in fires was associated with one species (a wetlands grass species). The impact on fire size was variable, with two species associated with larger fires, three species associated with smaller fires.
Individually, climate change is expected to increase the potential for fire occurrence by 150% by the end of the century based on projected changes in temperature and precipitation. Here we show that 8 invasive grass species are already associated with increased rates of fire occurrence by 27 to 230%, and 6 invasive grass species are associated with increased mean fire frequency by 24 to 150%, compounding current and future fire risk across the United States. [Fusco et al. 2019: 23595]
The authors suggest that fire and invasive species managers work together to create integrated management plans; otherwise, the convergence of human activities, climate change, and invasive species will continue to promote wildfires across the United States.
Smokey the Beaver: beaver‐dammed riparian corridors stay green during wildfire throughout the western USA, Fairfax and Whittle 2020
This study examines the positive effects of beaver damming on the resistance of landscapes to wildfire damage. The authors find that in riparian corridors (areas along rivers), the presence of beavers and their dams can create refuges that withstand blazes that consume surrounding vegetation.
Beavers play an important role in wetland habitats and are known as ecosystem engineers for the way they can shape landscapes with their activities. Beaver dams slow down water moving across a landscape, holding it in place for longer and allowing water to infiltrate into the soil, which raises water tables.
The combination of building flow obstructions (dams), accumulating water (ponds), and spreading that water out in the landscape (channels) gives beavers the unique potential to modulate environmental extremes such as flood and drought. When it comes to water, beavers slow it, spread it, and store it.
Due to the fact that beaver channels and dams spread water out in the landscape and store it broadly in adjacent soils, the vegetation near beaver ponds doesn’t experience as much reduced water availability during drought. Drought-stricken vegetation burns more easily than lush, green vegetation, so it follows that the vegetation around beaver ponds would be more difficult to burn than vegetation around undammed creeks [Fairfax and Whittle 2020: 1].
Fairfax and Whittle observed the effects of beaver dams on preventing fire spread to the areas where they had built dams, examples of which are shown in satellite imagery below.
The authors quantify the effects of beaver activity in fireproofing areas by examining the Normalized Difference Vegetation Index (NDVI) observed in satellite imagery before, during, and after wildfire years in the American West. They found that while vegetation is able to reestablish itself a year after fire damage regardless of beaver activity due to its own resilience to fire, areas in beaver dammed zones maintained vegetation even during wildfires, demonstrating actual resistance to blazes, not just the ability to recover after damage. They note how vital this is for those ecosystems and the life within them.
These ribbons of fire-resistant riparian corridor may be particularly important for species that are unable to physically escape wildfire. They can provide temporary habitat for fish, amphibians, reptiles, small mammals, wild and domestic ungulates, and birds that are unable to outrun/outfly the spread of flames. While we found that beaver activity does play a significant role in maintaining vegetation greenness during wildfires, it does not appear to play a significant role in the ability for a riparian corridor to rebound in the year following fire. Riparian vegetation NDVI rebounded in the year following the fire regardless of proximity to beaver activity. Thus, we would describe beaver activity as creating refugia during wildfire, but not necessarily changing the long-term landscape outcomes [Fairfax and Whittle 2020: 7].
The survival of wildlife is crucial to these ecosystems, and beaver activity uniquely contributes to the creation of refuge areas that resist burning and can provide shelter for animals during these destructive events. The authors conclude,
As it stands today, wetland habitat is very limited and beavers can create and maintain wetland habitat that persists through flood, drought, and, as we have shown in this study, fire. This has immediate relevance to scientists and practitioners across North America and Eurasia, particularly in places with increasing wildfire risk and existing or planned beaver populations. Perhaps instead of relying solely on human engineering and management to create and maintain fire-resistant landscape patches, we could benefit from beavers’ ecosystem engineering to achieve the same goals at a lower cost [Fairfax and Whittle 2020: 7].
Planned Herbivory in the Management of Wildfire Fuels, Nader et al. 2007
Nader et al. survey herbicides, prescribed fire, mechanized treatments, hand cutting, and grazing animals as fire management techniques. Managing vegetation involves “changing the plant community to decrease the flame height when fire occurs,” favoring native species that may be more resilient to fire, and altering the landscape to create fuel breaks, which are patches across which it is hard for fire to jump [Nader 2007: 18].
Focusing their analysis on grazing and the contexts in which it is most useful, the authors note that there are many site and animal specific factors to take into account for successful implementation.
[Grazing]is a complex, dynamic tool with many plant and animal variables, and it requires sufficient knowledge of the critical control points to reach treatment objectives. Those control points involve the species of livestock grazed (cattle, sheep, goats, or a combination); the animals’ previous grazing experience (which can affect their preferences for certain plants); time of year as it relates to plant physiology (animal consumption is directed by the seasonal nutrient content); animal concentration or stocking density during grazing; grazing duration; plant secondary compounds; and animal physiological state [Nader 2007: 19].
Grazing has the advantage of keeping nutrients in the ecosystem, unlike mechanical methods that harvest vegetation to be sold as biomass chips (like wood chips). This means that when animals digest vegetation and excrete on the landscape, they participate in the local nutrient cycle. Animals also trample soil, which can crush fine fuel and mix it into the soil, where it cannot contribute to ignition, which reduces one contributing factor to persistent and destructive blazes. Animals do more than just remove extra vegetation – they can have many beneficial interactions within a given ecosystem.
Any grazing plan designed for fuel reduction needs to consider the grazing impacts on parameters other than just simply reduction. The effects of the grazing management should be studied for their impact on water quality, compaction, riparian vegetation, disease interaction with wildlife (bluetongue, pasturella), and weed transmission. The positive aspects of grazing over other treatments also should be weighed, including recycling of nutrients into the products of food and fiber [Nader 2007: 22].
By introducing grazing animals into a landscape or agricultural system, managers can affect biodiversity in complex ways. The authors mention that “Hadar et al. reported that light grazing increased plant diversity on treated sites. Thus, when proposing a stocking rate for treatment consumption, the environmental impact needs to be considered” [Nader 2007: 22].
Nader et al. conclude that “grazing is best used when addressing vegetation with stems of smaller diameters that make up the 1- and 10-hour fuels. These two fuel classes are important because they can greatly impact the rate of spread of a fire, as well as flame height” [Nader 2007: 19]. While they call for further research to validate anecdotal accounts supporting grazing and understand its best practice, they maintain that “prescribed grazing has the potential to be an ecologically and economically sustainable management tool for reduction of fuel loads” [Nader 2007: 20].
Landscape rehydration ‘better than dams’ in improving farm production, reducing fire risk, Major 2020
A project in Queensland, Australia has met with success in its efforts to rehydrate the landscape on the farmland property of Worona Station, improving biodiversity, water retention, and resistance to erosion and fire. Worona Station had been degraded and faced serious erosion issues, so Chris Le Feuvre, the owner, partnered with consultancy groups of NQ Dry Tropics and the Mulloon Institute in a project to rehydrate his land.
The project team has used a combination of planned grazing and small, low-tech dams to combat erosion problems. The grazing technique involves:
Splitting paddocks into small sizes and using large mobs of cattle grazing on rotation … grazing pasture more intensively while giving it longer to rest, [thereby] increasing carrying capacity.
Grazing in this way (which is evocative of Allan Savory’s Holistic Planned Grazing methodology) has resulted in increased pasture species diversity and boosted plant growth, allowing the Le Feuvre to double his herd size. Planned grazing has also reduced sediment runoff from the property. Sam Skeat, a grazing officer with NQ Dry Tropics, attests to the importance of grazing.
The plug-and-pond technique — also known as leaky weirs — involves small dam-like structures to lift the bed level of the water, which is then run onto the floodplain to grow pasture and recharge aquifers. While weirs have been strategically constructed, Mr. Skeat said grazing management was the most important tool to improve water retention in a landscape. ‘If you can use cattle as a tool to regenerate the grassland, you’ll get more infiltration, slow the flow, hold water up in the landscape and have you growing grass for longer’ [Major 2020].
Rehydrating landscapes can improve their resilience to extreme events, and improve their quality in the face of chronic problems like erosion. According to the Mulloon Institute Chairman Gary Nairn, the issue of degraded gullies and streams is a national concern. Gullies are created when parched land is unable to absorb rainwater, allowing it to run off. The sediment-filled runoff ends up in the ocean, polluting it.
Nairn sees land rehydration through planned grazing and related techniques as a better solution than building a massive, industrial-scale dam to retain water. The Australian government has been looking into building new large dams. Levels at Warragamba Dam, which supplies about 80 percent of Sydney’s water, have dropped to less than half capacity.
‘We’ve been able to demonstrate in Mulloon, if we repaired and rehydrated the catchment through to the Sydney water supply, you could store the equivalent of Warragamba Dam,’ he said [Major 2020].
Well-watered mulberry tree credited with saving home on NSW South Coast from summer bushfires, Aubrey 2020
A well-watered mulberry tree has been credited with averting the danger of destructive wildfires from destroying Brett Hawkins’ home during 2020’s unprecedented fire season in Australia. When massive fires raged through the bush through the summer, many homes were completely engulfed. However, Hawkins attested that when he returned to his home after evacuating,
‘I could see straight away the house was intact — the roof was intact, but everything else around it was burnt, with the exception of the mulberry tree.’ He described the stark scene greeting him upon arriving back home, ‘It was apocalyptic,’ Mr. Hawkins said. ‘There was not a tree left, ash on the ground and smouldering embers everywhere.’ But among the blackened trees, Mr. Hawkins found his mudbrick house and mulberry tree in full leaf [Aubrey 2020].
In the season’s drought, he had been rationing water, but sparing some to keep his tree hydrated and healthy, which may have been a contributing factor in its resistance. According to the article, “Mr. Hawkins believed that by heavily watering the tree, combined with luck regarding which direction the fires came, the full heat of the bushfires was shifted.” [Aubrey 2020]
Another important feature is the mulberry’s lack of dried leaves and brush at its base that might pose a danger of igniting. Other species, like eucalyptus, with oily leaves that could dry out, or pine trees whose long branches catch dry leaves, are less ideal.
A tree expert from the Fenner School of Environment at the Australian National University analyzed some of the possible factors leading to the survival of this mulberry tree and what it might teach those wishing to fortify the fire resilience of their homes and properties. “While there does not seem to be a clear answer on what to plant to ‘fire-proof’ your house, Professor Brack said a well-watered tree, with a clear trunk and no loose, dry leaves or branches is a good start” [Aubrey 2020].
All over the world, from Australia to Europe to North and South America, wildfires have waged destruction on natural landscapes and human settlements alike. The devastation of these disasters is heartbreaking, and the images of catastrophe – walls of flame, scorched wildlife, a world gone red – are unforgettable. There is no more potent image of the climate crisis than the towering infernos and eerie, hellish, smoke-filled skies that we’ve seen in this past year.
The question is how we best confront the issue, keeping people safe and ecosystems intact as much as possible. Given that the hotter, drier conditions that climate change causes are exacerbating wildfire seasons in both duration and destructiveness, it is urgent that we better understand fire’s natural role in ecosystems and the conditions that cause fires to become ultra-destructive. Wildfire is a complicated global problem that requires locally-informed responses adapted to local ecosystems.
“Fuel load” reduction is a major solution, and this may be part of the answer in certain cases.However, to the extent that hauling out vegetation from a fire-prone site further degrades that ecosystem, this practice may only exacerbate the problem in the long run. Fortunately, many alternative and complementary practices to diminish wildfire ferocity are known. This includes, for example, indigenous prescribed burning, promoting ecosystem health to reduce dryness and drought, favoring native species, and enacting land use policies that discourage development in fire-prone areas.
In discussion of fire and its risks, it is important to first note that not all fire is problematic, and the ideal healthy ecosystem would not necessarily be one without any fire. For example, species in fire-evolved landscapes depend on cyclical fires, and burning can also increase landscape heterogeneity and biodiversity.
In discussion of fire and its risks, it is important to first note that not all fire is problematic, and the ideal healthy ecosystem would not necessarily be one without any fire.
Moreover, a world without fire is impossible, and focusing on total fire suppressionis misguided. As environmental historian Stephen Pyne warns, “Removing fire from landscapes that have co-evolved or co-existed with it can be as ruinous as putting fire into landscapes that have no history of it” [Pyne 2020]. In fact, we can fight fire with fire by planning controlled burns, part of the strategy of managing the vegetative fuel available to wildfires.
Since grasses, shrubs, and trees become fuel for a fire, much scientific literature on wildfire examines how to reduce this fuel so that when wildfire does occur, it is less destructive and less far-reaching. Invasive plants are a particularly problematic form of fuel, because they haven’t necessarily evolved in fire-shaped landscapes. Fires can quickly gain in intensity when consuming them, and invasive grasses alter the cyclical availability of fuel and increase the frequency of fires [Fusco 2020].
In contrast, native plants in fire-prone ecosystems often evolve with fire playing an ecological role in their habitat, leading them to be more resistant and hardy in destructive blazes. Thus there is a clear advantage to favoring native species over invasive ones, especially since native species also perform ecological functions necessary for the health of the ecosystem and its inhabitants.
It is important to understand that more plant life does not necessarily translate to more fire fuel. The quality of landscape and details such as height, density, fuel bed depth and fuel moisture all matter for fire spread [Nader 2007]. Furthermore, not all vegetation is equally vulnerable. This is not just a matter of the differences between species, but also how landscape differs in its spacing, density, and topological features, making some landscapes more vulnerable to fire spread than others.
Often, our problems with fire are also problems of water. How well hydrated a landscape is can make a crucial difference to its vulnerability to wildfire. In fact, the capacity of land to retain water is linked to vegetation, and specifically to biodiverse vegetated areas. A healthy and hydrated tree or patch of forest may withstand fire that drier and more brittle vegetation does not. Healthy trees may even protect properties from fire, as anecdotes attest [Aubrey 2020].
Plant roots, along with the mycorrhiza and microbes that make up a symbiotic web of linked organisms within healthy soil, create a porous soil structure that water infiltrates when it rains; the soil thus acts as a sort of natural sponge. Unlike dehydrated and degraded soil, which do not absorb and retain water effectively, living soil soaks up precipitation; this reduces runoff and erosion during heavy deluges. Water infiltrating healthy soil hydrates organic matter, is retained in topsoil pores, or makes its way to the water table below ground, which can be thought of as a bank for water. When dry seasons or droughts arrive, this bank provides the moisture needed to keep vegetation healthy–and, when rains arrive again, this bank refills.
Plant presence helps build healthy soil, and it also contributes to the small water cycle (the circulation of water evaporating from land and falling in the form of precipitation over the same environment). For example, scientists have found that many plants release microbes which are borne up on evaporated water droplets and catalyze cloud formation and precipitation in the atmosphere, a phenomenon known as bioprecipitation [Morris 2014]. Plants don’t just need rain – they help create it too. Well-hydrated, fire-resistant plants are part of the key to retaining moisture in landscapes and sapping wildfire of its power.
Beavers also contribute to water retention on the landscapes they inhabit. They play a major ecological role by creating and preserving wetlands, so much so that they are sometimes called ecosystem architects or engineers.The hydrating effects of beaver activities have even been found to create areas that resist fire even while neighboring landscapes burn. Some experts suggest that “perhaps instead of relying solely on human engineering and management to create and maintain fire-resistant landscape patches, we could benefit from beavers’ ecosystem engineering to achieve the same goals at a lower cost” [Fairfax and Whittle 2020: 7].
Beavers are not the only type of animal life that can help in fire management. Ruminants like goats, cows and sheep can control vegetation, while also improving soil health and promoting water retention in well-managed grazing systems. The movement of hoofed grazing animals across grasslands breaks up soil, making it easier for water to infiltrate, while the animal manure provides natural fertilizer.
Another benefit to controlling vegetation through animal grazing is the retention of biomass on the landscape in the form of nourishing manure, rather than clearing it away via logging or burning, for example [Nader 2007]. Grazing also presents opportunities for diversified economic activity within agricultural or silvopasture (the practice of integrating trees, forage, and the grazing of domesticated animals in a mutually beneficial way) systems. In case studies, managed grazing has made farm operations more productive and profitable while at the same time promoting ecological health [Major 2020]. Grazing animals also can increase ecosystem biodiversity[2].
Finally, there is the role of humans and where we choose to live. Part of the reason fires cause such harm to human settlements is because we build houses in fire-prone areas. The wildland-urban interface (WUI) is the site of a large percentage of fires, including particularly destructive fires, and increasing human encroachment of wild areas means that more people are brought into closer range of wildfires, while also negatively impacting ecosystems and their biodiversity. Of course, one bold strategy to reduce fire risk is to change our land use [Syphard 2013].
Part of the reason fires cause such harm to human settlements is because we build houses in fire-prone areas.
As modern human settlements have expanded, we have designed our fire management strategies to focus almost exclusively on suppression in most places, enforcing a dominant paradigm that fire is always bad and should be eliminated from landscapes. Unfortunately, such policy allows the very buildup of vegetation that can fuel increasingly destructive wildfires.
However, alternative relationships to fire have been practiced across human history, such as the controlled burning used by Indigenous communities across the world, from California to Australia. Recently, the depth of these communities’ ecological knowledge is beginning to gain the respect it deserves and to be considered and implemented in mainstream fire practices. In certain circumstances, it is now accepted that controlled burns can help ensure healthy ecosystems by decreasing the destructiveness and frequency of wildfires.
Although the use of prescribed fire is one way to manage vegetation and shape a varied landscape, any strategy that combines fire management and ecological stewardship will be full of site-specific complexity. Thus, it is critical to understand how Indigenous stewardship has been carried out over generations on given landscapes and to factor this knowledge into strategies to combat wildfire and ensure ecological health.
Much of the world’s remaining biodiversity resides in land inhabited by Indigenous groups, whose fire and land management practices come from a deep cultural and spiritual context. That is, “Indigenous fire management is effective in that it is an emergent property of a linked social-ecological system where Indigenous knowledge and culture, and associated livelihoods, are intimately interconnected with landscape management practices” [Mistry 2016: 4].
A path forward into a fire-resilient age might be led by Indigenous groups implementing local, community-owned solutions. Society at large would benefit from supporting and learning from the local communities who have generational knowledge of local ecosystems and fire management. In addition, land stewardship using practices that favor native species, biodiversity, living soils, and aim to retain moisture on the land should be applied on a wide scale. The articles that follow offer detail and insight into these approaches.
Wildfire article summaries
Our burning planet: why we must learn to live with fire, Pyne 2020
Steven J. Pyne is an emeritus professor at Arizona State University and the author of several books on fire history and policy. He wrote this opinion piece as a protest against the prevention and suppression of wildfires in our land management process. He argues that revising our perception of fire and accepting its presence in ecosystems is critical to our ongoing relationship with our planet.
He describes “a paradox at the core of Earth’s unraveling firescapes,” that “we have too many bad fires — fires that kill people, burn towns, and trash valued landscapes. We have too few good ones — fires that enhance ecological integrity and hold fires within their historic ranges” [Pyne 2020]. Operating under a paradigm of total fire suppression leads us astray in managing landscapes, while we so readily accept fire in the form of fossil fuel combustion in so much of our lives. Pyne sees these behaviors as evidence that our relationship with fire is out of whack.
He stresses the importance of distinguishing between burning in living ecosystems and burning the fossils of life from past ages.
The critical contrast lies in a deeper dialectic than burned and unburned landscapes. It is a dialectic between burning living biomass and burning fossil biomass. We are taking stuff out of the geologic past, burning it in the present with all kinds of little understood consequences, and passing the effluent into the geologic future.
…
Fires in living landscapes come with ecological checks and balances. Fires in lithic landscapes have no boundaries save those humans impose on themselves [Pyne 2020].
Pyne associates three paradoxes with our current fire policies. First, abandoning a traditional lore of “light burning” has removed good fires and left us with only bad and harmful ones. When controlled burns are not practiced regularly to manage landscapes, vegetation can build up and fuel the intensity and spread of uncontrolled blazes that spark.
Second and surprisingly: “The Earth does not have more fire today than before fossil fuels emerged as a primary source of energy: It has significantly less” [Pyne 2020]. That is, the amount of land burned in fires has actually decreased, while the presence of intense “feral flames” has increased. The decrease in the scope of fire is largely due to the move away from fire’s use in agriculture and its replacement with modern techniques, including machinery powered by combustion. As Pyne describes,
Farmers had relied on fire to fertilize, fumigate, and alter microclimates. Fire did all this in one catalytic process that self-propagated. But with the transition to fossil biomass, modern agriculture found surrogates with artificial fertilizers, pesticides, and herbicides, and it now had the fossil-fuel-powered machines to distribute them. Production became more efficient; transport, more dense. As agriculture joins a modern economy, working flames recede [Pyne 2020].
What we are left with is intense, destructive wildfire, rather than helpful working fire. Pyne points out that we now only see one half of fire’s possibilities, since the working fire shaping landscapes and agricultural systems is notably absent. He says: “Landscape fire fades; what fire persists tends to be outbreaks of feral fire. We see those oft-disastrous flames. We don’t see the lost fires or the sublimated fires in machines that removed them” [Pyne 2020].
The third paradox is that as we reduce our use of fossil fuels going forward (as one sort of fire), we will have a greater need “to manage fire in living landscapes.” So Pyne calls for us “to reinstate the right kind of fire, and … adapt to fire’s presence and let it do the work for us.” He pushes for the recognition that transitioning away from our reliance on fossil fuel burning is an important but incomplete step in balancing ecosystem health. Fires in living systems have an important role to play, and according to Pyne, “the need is not just to reduce fuels to help contain wildfires; those missing fires did biological work for which no single surrogate exists” [Pyne 2020]. He asserts that we will need to reintroduce fire as a staple tool on our landscapes.
Fires in living systems have an important role to play, and according to Pyne, “the need is not just to reduce fuels to help contain wildfires; those missing fires did biological work for which no single surrogate exists.” He asserts that we will need to reintroduce fire as a staple tool on our landscapes.
Pyne concludes with a call for the overhaul of our conceptual and policy treatment of fire.
Anthropogenic fire needs more room to maneuver – more geographic space, more legal space, more political space, more conceptual space. … Equally, society needs to rethink liability law to reduce the risks incurred by fire officers doing a necessary job …; adapt air quality regulations …; and tweak National Environmental Policy Act review processes … [to] accommodate the realities of restored fire at a landscape scale. … Communities in the fire equivalents of floodplains need hardening [Pyne 2020].
He proposes that fire restoration jobs can replace those lost from forestry and fire suppression. He admits that our understanding of fire biology requires more research, and that our greatest need is for “a working fire culture … that ensures fire’s proper place in the landscape” by renewing “our ancient alliance” with fire and making it “an indispensable friend.”
Fire Myths, Hanson 2018
In this podcast interview, Dr. Chad Hanson, an ecologist and fire researcher, shares his perspective on the 2018 wildfires in the American West and some myths that have circulated about fire management in their wake.
First, there is a perception that wildfires in forested regions are so devastating that they reverse the ‘carbon sink’ effect of forests, releasing the carbon of the burned biomass back into the atmosphere. Forests still sequester large amounts of carbon, even if they experience wildfires, because most of the forest remains intact even through blazes. Models that fault wildfires for turning forests into net carbon emitters rest on the assumption that all of the carbon that would usually be stored in a forest is combusted during a fire, but this is far from the reality, in which just a small fraction of a forest’s biomass is consumed. As Hanson says,
In fact, even in the most intensely burned patches where a fire kills all the trees (which in reality, even in the biggest fires, it’s only a small portion, a minor portion of the overall fire)… But even in those areas, only about two or three percent of the above ground biomass is actually consumed, in other words, ends up as carbon. The trees are still standing there [Hanson 2018].
Second, the intensity of fires has not been universally increasing in recent years. We are experiencing a lot of geographically large fires, but these are not necessarily high intensity fires. The percentage of high intensity fire today is similar to historical precedent, and overall, there is much less fire in our landscapes now than in Earth’s geological history. Further, even in the highest intensity fires, forests are never decimated past the point of no return. Trees and vegetation are reestablished after fires, and the ash left behind is dense with nutrients, promoting new growth. Even dead trees, which have long been thought to be responsible for contributing to high intensity fires, are actually not shown to drive fire intensity, according to Hanson.
This series of myths – that forest fires are raging with high intensities, that they burn up so much biomass as to make forests ineffective sources of carbon sequestration, and that the only way to manage forests to avoid these outcomes is to thin out the trees – hinder our understanding of forest management and allow false and harmful solutions to propagate. As a result of these perceptions, proponents of logging have pushed to expand logging operations, purportedly as a fire management strategy. However, according to Hanson, logging is actually linked to greater fire intensity. He explains that small materials, like twigs, are more flammable than trunks.
Tree trunks are not combustible. They really just don’t burn. Again, outer bark can burn, but the trees themselves don’t burn. What logging does is it removes noncombustible material essentially from the forest and leaves behind very combustible kindling, like slash debris – the branches and small twigs and things like that that are not possible to get up off the forest floor after the tree trunks are removed and that’s very combustible.
The other thing that logging does is that it reduces the cooling shade of the forest canopy. By removing a lot of trees, you have more sunlight reaching the forest floor, and what that does is it creates hotter and drier conditions and that means everything on the forest floor gets more dried out, more potentially combustible, and logging also spreads invasive weeds like cheatgrass, which is very, very flammable. Cheatgrass loves a lot of sunlight and so you get a lot of that after intensive logging.
And the last [problem with logging] is a little bit more technical, but basically when you have a lot more trees, it cuts down on the wind speeds that drive fires. It has a buffering effect in a sense. And when a lot of the trees are removed, that buffering effect is reduced or eliminated and fire spreads through those forests faster [Hanson 2018].
This three-fold effect of logging makes forests more vulnerable, and it is important to dispel the concept that removing trees is the best way to keep people safe from fires. Hanson criticizes the opportunism of using these fire myths to advance an agenda of logging. He cautions that when fire science and policy emerge from the U.S. Forest Service, which manages national forests and gains a good deal of revenue from logging, there is a perverse incentive to keep practicing logging as fire management. He calls for clearer and more public communication from scientists to dispel fire myths and share recent findings that have been shifting so much of what we know about fire science.
Hanson says that the best strategy to ensure the protection of homes from wildfire is to focus on the homes themselves. This can be done by using fire resistant building materials, fire-proofing roofs, erecting rain gutter guards to prevent the accumulation of small fuels like pine needles, pruning the vegetation in a 100-foot radius of a house, and removing small shrubs and branches of mature trees, while leaving those trees standing. Fire management interventions at this level are shown to be far more effective at preventing damage than attempts to control the fuel load of fires within forests.
Hanson points out the need to decouple fires that occur in remote forest ecosystems and those that rage through human settlements and urban communities, because thinning out vegetation in attempts to suppress the former do not actually protect against the latter. In fact, thinning forests undermines the ecological processes that fire serves in forest systems. When discussing fires that have devastated homes and lives, he says
I mean, where’s the forest in Malibu? There’s no forest. These are chaparral ecosystems, most of the fires that are burned homes and lives have been lost are not in forest. In fact, they’re mostly nowhere near forests. They’re in grasslands, chaparral shrub habitat, oak woodlands. But the areas that are in forest, where we’ve had tragic loss of homes and lives, these are mostly areas where we’ve had intensive logging, and it’s like I mentioned earlier, you know, more logging is typically associated with more intense fire at a faster rate of spread. [Hanson 2018]
He advocates for a greater focus on fire prevention around homes and communities themselves, in what is known as ‘defensible space.’ He points out that such measures are a great source of jobs, as well as an effective intervention curbing the destructiveness of fires. With a shift in focus from forest thinning to fire-proofing, and better understanding and communication of fire science, we can let go of some of the fire myths that have been dictating policy and failing to meet public needs.
More logging is typically associated with more intense fire at a faster rate of spread [Hanson 2018].
Land use planning and wildfire: development policies influence future probability of housing loss, Syphard et al. 2013
Wildfire is a challenge that threatens human settlement at an increasing scale, but planning and development does not always address this threat. In fact, policy around land use is in large part responsible for the destruction of homes and property and the threat to human life that occurs in wildland-urban interfaces (WUIs). While there is much literature on how to suppress fires, mitigate their damage, or manage for less destructive fire seasons, a more far-reaching strategy would be to stop building in fire prone areas. Land use decisions can be improved to lessen the risk of infrastructure loss and foster healthy ecosystem function.
Land use planning is an alternative that represents a further shift in thinking, beyond the preparation of communities to withstand an inevitable fire, to preventing new residential structures from being exposed to fire in the first place. The reason homes are vulnerable to fires at the wildland-urban interface is a function of its very definition: “where homes meet or intermingle with wildland vegetation”. In other words, the location and pattern of homes influence their fire risk, and past land-use decision-making has allowed homes to be constructed in highly flammable areas. [Syphard 2013: 1-2]
In many areas, including in California, we have come to expect fire, but have not necessarily learned to live with it. The authors of this study analyzed what types of human development carried out in the next several years might contribute to or avert the risk of fire damage. They examined the South Coast Ecoregion of San Diego County, which they describe as:
topographically diverse with high levels of biodiversity, and urban development has been the primary cause of natural habitat loss and species extinction. Owing to the Mediterranean climate, with mild, wet winters and long summer droughts, the native shrublands dominating the landscape are extremely fire-prone [Syphard et al. 2013: 2].
This study acknowledges the responsibility humans have in shaping the landscape and its biodiversity, and in contributing to fire activity by building into wild areas and expanding WUI. They sought to understand how patterns of development and housing density might influence future fire spread and intensity. They found that
structures in areas with low- to intermediate- housing density were most likely to burn, potentially due to intermingling with wildland vegetation or difficulty of firefighter access. Fire frequency also tends to be highest at low to intermediate housing density, at least in regions where humans are the primary cause of ignitions [Syphard 2013: 2].
Though it is impossible to reverse the effects of policies that have shaped the fire landscape we have today, understanding the way human behavior contributes to our own risk of harm from wildfire can help us plan intelligently going forward. The authors conclude that
With projections of substantial global change in climate and human development, we recommend that land use planning should be considered as an important component to fire risk management, potentially to become as successful as the prevention of building on flood plains. History has shown us that preventing fires is impossible in areas where large wildfires are a natural ecological process. As Roger Kennedy put it, “the problem isn’t fires; the problem is people in the wrong places [Syphard 2013: 10-11].
Community owned solutions for fire management in tropical ecosystems: case studies from Indigenous communities of South America, Mistry et al. 2016
Indigenous groups across the world have developed ecological knowledge linked to the places they inhabit, including prescribed fire practices used to maintain healthy ecosystems. Mistry et al. examine the challenges Indigenous communities in South America face in managing the landscape through fire and preserving such knowledge across generations in sometimes hostile political climates. However, there is growing recognition that Indigenous people have a vital role to play in combating climate change and supporting biodiversity and healthy ecosystems.
Emerging research shows the fundamental role of Indigenous land-use practices for controlling deforestation and reducing CO2 emissions—analysis of satellite imagery suggests that Indigenous lands have reduced rates of deforestation and habitat conversion, and lower greenhouse gas (GHG) emissions, compared with surrounding areas [Mistry 2016: 1].
While indigenous groups’ use of prescribed fire early in the dry season to prevent destructive out-of-control fires is gaining broad recognition, that hasn’t necessarily translated into greater respect or autonomy for those communities. Instead, Indigenous people may be given auxiliary roles in fire management, or have their knowledge utilized but implemented by non-local organizations in structures that fail to benefit or empower the local communities themselves. While this may still achieve desired wildfire management results, it weakens intergenerational knowledge transfer and undermines the social and spiritual role of prescribed fire within communities.
Mistry et al. argue that “Indigenous fire management is effective in that it is an emergent property of a linked social-ecological system where Indigenous knowledge and culture, and associated livelihoods, are intimately interconnected with landscape management practices” [Mistry 2016: 4]. Precisely because prescribed fire matters to Indigenous communities as something more than a tool in the toolkit of managing wildfires, it is effective when carried out by those communities in reducing risk of destructive wildfires and supporting healthy and biodiverse ecosystems.
Importantly, the numerous uses of fire mean that burning is a relatively constant activity, particularly during the dry season, generally at low levels, thereby helping to prevent the build-up of flammable fuel and incidents of large-scale uncontrollable wildfires. Experimental studies of fire behaviour suggest that this patch mosaic burning not only reduces the occurrence of dangerous fires, but also increases spatial and temporal vegetation heterogeneity and biodiversity [Mistry 2016: 4].
These authors distinguish between Indigenous relationships to ecosystems and market-based approaches to ecosystem services valuation, which attempt to incentivize conservation through payment. While the goal of the market-based approach is to monitor and preserve functioning ecosystems, “their ideological foundations within a neoliberal agenda that promotes ‘selling nature to save it’ is in stark contradiction with Indigenous ontologies based on human–nonhuman–spiritual relationships” [Mistry 2016: 2].
Within Indigenous communities, fire plays a role in social bonding, intergenerational knowledge transfer, and agricultural practices. Mistry et al. argue that
savanna and forest ecosystems are being protected within Indigenous lands not because they are being ‘managed’ in a direct and active way, but as the indirect outcome of a healthy social–ecological system, i.e. the outcome of practices that maintain social and ecological integrity, or what can be termed ‘community owned solutions [Mistry 2016: 4].
But challenges, including loss of fire knowledge by younger generations within Indigenous groups because of outside pressures and encroachment, pose a threat to these fire management practices. For example, in Venezuela and Brazil,
young Wapishana and Makushi and some community leaders were more critical about the use of fire as they had more regular contact with state natural resource management officials and environmental organizations that promoted antifire discourses. As with the Krahô, changing Indigenous values to focus on fire prevention and suppression could have the effect of making the problem worse [Mistry 2016: 4].
That is, when prescribed burning is taken out of its original context and represented to younger generations of Indigenous people and land stewards as simply a well-incentivized tool, the Indigenous communities themselves are diminished, along with the robustness of their ecological knowledge that gets passed forward.
In spite of lingering antagonistic views in Brazil and Venezuela toward indigenous fire management, attitudes are changing.
Not only is there a move away from categorizing all fire as ‘bad’; there is also a recognition that Indigenous fire knowledge is a valid form of knowledge that could inform policy-making [Mistry et al. 2016: 6].
Mistry et al. suggest the best way to achieve both ecological and communal health might be through power-sharing arrangements. By empowering Indigenous communities, national governments could in turn work toward their fire management and biodiversity conservation goals. This might require evaluating ecological health in ways beyond just quantitative metrics, which reduce these complex systems down to a set of standardized numbers, as well as the recognition that the well-being of these ecosystems is tied to the Indigenous communities that inhabit them, according to the authors:
There needs to be enabling policies that focus on legitimizing and strengthening Indigenous fire management as a community owned solution. Critically, as community owned fire management is intricately linked with Indigenous survival strategies, so too must firefighting and prescribed burning be grounded in local social–ecological systems. We believe it is necessary to define long-term actions to support the integrated functioning and survival of Indigenous communities as a whole, rather than focusing on isolated issues (e.g. carbon retention) or benefits for some individuals (e.g. hiring Indigenous firefighters) [Mistry 2016: 8].
This systems approach may well be the key to successful long-term fire management. The authors offer this challenge:
What we want to do is not promote one over the other, but encourage decision-makers to engage with, and appreciate, Indigenous perspectives and worldviews on fire management. Community owned solutions acknowledge collectivity, spirituality, process orientation and locality, whereas many expert-led fire management interventions often result in promoting individualism, ethnocentrism, rationality, efficiency, commercialism and globalization. The question we raise is this: can the ‘community owned solutions’ approach be the mechanism through which Indigenous perspectives can be represented within fire management [Mistry et al. 2016: 8]?
Invasive grasses increase fire occurrence and frequency across US ecoregions, Fusco et al. 2019
It has long been suspected that the increasing abundance of invasive grass species may contribute to wildfires in the United States by adding abundant new fuels to ecosystems, increasing the range of conditions that lead to fire ignition, and enabling the development of larger, hotter fires. The new fire regimes (patterns of fire duration, intensity, and spread) that emerge can in turn destabilize wildlife and lead to local extinctions while expanding favorable habitat for the invasive species, for many of these grasses recover quickly after fires, providing renewed fuel and potentially increasing the frequency of fires.
The authors of this paper provide a comprehensive analysis on the impact of 12 non-native grasses on the occurrence (whether a fire occurred in a particular place), frequency (how many times a place burned), and size of wildfires. The research was conducted across 29 US ecoregions, including deserts, temperate forests, wetlands, woodlands, river valleys, shrublands, and coastal plains. Data were collected and combined from fire records and records of invasive grasses, and results from “invaded” regions and nearby “uninvaded” regions were compared. The authors also considered human activities and ecological factors related to fire.
One of the most notorious impacts of nonnative, invasive grasses is the alteration of fire regimes. Yet, most evidence of these impacts comes from local-scale studies, making it unclear whether they have broader implications for national and regional fire management. Our analysis of 12 invasive grasses documents regional-scale alteration of fire regimes for 8 species, which are already increasing fire occurrence by up to 230% and fire frequency by up to 150%. These impacts were demonstrated across US ecoregions and vegetation types, suggesting that many ecosystems are vulnerable to a novel grass-fire cycle. Managing existing grass invasions and preventing future introductions presents a key opportunity to remediate the ecological and economic consequences of invasive species and fire [Fusco 2019: 23594]
The results of this analysis showed that 8 of the 12 invasive grass species examined were associated with significantly higher fire occurrence and fire frequency, and that fire occurrence more than doubled for two of these species. Three of the species did not impact fire occurrence, and a decrease in fires was associated with one species (a wetlands grass species). The impact on fire size was variable, with two species associated with larger fires, three species associated with smaller fires.
Individually, climate change is expected to increase the potential for fire occurrence by 150% by the end of the century based on projected changes in temperature and precipitation. Here we show that 8 invasive grass species are already associated with increased rates of fire occurrence by 27 to 230%, and 6 invasive grass species are associated with increased mean fire frequency by 24 to 150%, compounding current and future fire risk across the United States. [Fusco et al. 2019: 23595]
The authors suggest that fire and invasive species managers work together to create integrated management plans; otherwise, the convergence of human activities, climate change, and invasive species will continue to promote wildfires across the United States.
Smokey the Beaver: beaver‐dammed riparian corridors stay green during wildfire throughout the western USA, Fairfax and Whittle 2020
This study examines the positive effects of beaver damming on the resistance of landscapes to wildfire damage. The authors find that in riparian corridors (areas along rivers), the presence of beavers and their dams can create refuges that withstand blazes that consume surrounding vegetation.
Beavers play an important role in wetland habitats and are known as ecosystem engineers for the way they can shape landscapes with their activities. Beaver dams slow down water moving across a landscape, holding it in place for longer and allowing water to infiltrate into the soil, which raises water tables.
The combination of building flow obstructions (dams), accumulating water (ponds), and spreading that water out in the landscape (channels) gives beavers the unique potential to modulate environmental extremes such as flood and drought. When it comes to water, beavers slow it, spread it, and store it.
Due to the fact that beaver channels and dams spread water out in the landscape and store it broadly in adjacent soils, the vegetation near beaver ponds doesn’t experience as much reduced water availability during drought. Drought-stricken vegetation burns more easily than lush, green vegetation, so it follows that the vegetation around beaver ponds would be more difficult to burn than vegetation around undammed creeks [Fairfax and Whittle 2020: 1].
Fairfax and Whittle observed the effects of beaver dams on preventing fire spread to the areas where they had built dams, examples of which are shown in satellite imagery below.
The authors quantify the effects of beaver activity in fireproofing areas by examining the Normalized Difference Vegetation Index (NDVI) observed in satellite imagery before, during, and after wildfire years in the American West. They found that while vegetation is able to reestablish itself a year after fire damage regardless of beaver activity due to its own resilience to fire, areas in beaver dammed zones maintained vegetation even during wildfires, demonstrating actual resistance to blazes, not just the ability to recover after damage. They note how vital this is for those ecosystems and the life within them.
These ribbons of fire-resistant riparian corridor may be particularly important for species that are unable to physically escape wildfire. They can provide temporary habitat for fish, amphibians, reptiles, small mammals, wild and domestic ungulates, and birds that are unable to outrun/outfly the spread of flames. While we found that beaver activity does play a significant role in maintaining vegetation greenness during wildfires, it does not appear to play a significant role in the ability for a riparian corridor to rebound in the year following fire. Riparian vegetation NDVI rebounded in the year following the fire regardless of proximity to beaver activity. Thus, we would describe beaver activity as creating refugia during wildfire, but not necessarily changing the long-term landscape outcomes [Fairfax and Whittle 2020: 7].
The survival of wildlife is crucial to these ecosystems, and beaver activity uniquely contributes to the creation of refuge areas that resist burning and can provide shelter for animals during these destructive events. The authors conclude,
As it stands today, wetland habitat is very limited and beavers can create and maintain wetland habitat that persists through flood, drought, and, as we have shown in this study, fire. This has immediate relevance to scientists and practitioners across North America and Eurasia, particularly in places with increasing wildfire risk and existing or planned beaver populations. Perhaps instead of relying solely on human engineering and management to create and maintain fire-resistant landscape patches, we could benefit from beavers’ ecosystem engineering to achieve the same goals at a lower cost [Fairfax and Whittle 2020: 7].
Planned Herbivory in the Management of Wildfire Fuels, Nader et al. 2007
Nader et al. survey herbicides, prescribed fire, mechanized treatments, hand cutting, and grazing animals as fire management techniques. Managing vegetation involves “changing the plant community to decrease the flame height when fire occurs,” favoring native species that may be more resilient to fire, and altering the landscape to create fuel breaks, which are patches across which it is hard for fire to jump [Nader 2007: 18].
Focusing their analysis on grazing and the contexts in which it is most useful, the authors note that there are many site and animal specific factors to take into account for successful implementation.
[Grazing]is a complex, dynamic tool with many plant and animal variables, and it requires sufficient knowledge of the critical control points to reach treatment objectives. Those control points involve the species of livestock grazed (cattle, sheep, goats, or a combination); the animals’ previous grazing experience (which can affect their preferences for certain plants); time of year as it relates to plant physiology (animal consumption is directed by the seasonal nutrient content); animal concentration or stocking density during grazing; grazing duration; plant secondary compounds; and animal physiological state [Nader 2007: 19].
Grazing has the advantage of keeping nutrients in the ecosystem, unlike mechanical methods that harvest vegetation to be sold as biomass chips (like wood chips). This means that when animals digest vegetation and excrete on the landscape, they participate in the local nutrient cycle. Animals also trample soil, which can crush fine fuel and mix it into the soil, where it cannot contribute to ignition, which reduces one contributing factor to persistent and destructive blazes. Animals do more than just remove extra vegetation – they can have many beneficial interactions within a given ecosystem.
Any grazing plan designed for fuel reduction needs to consider the grazing impacts on parameters other than just simply reduction. The effects of the grazing management should be studied for their impact on water quality, compaction, riparian vegetation, disease interaction with wildlife (bluetongue, pasturella), and weed transmission. The positive aspects of grazing over other treatments also should be weighed, including recycling of nutrients into the products of food and fiber [Nader 2007: 22].
By introducing grazing animals into a landscape or agricultural system, managers can affect biodiversity in complex ways. The authors mention that “Hadar et al. reported that light grazing increased plant diversity on treated sites. Thus, when proposing a stocking rate for treatment consumption, the environmental impact needs to be considered” [Nader 2007: 22].
Nader et al. conclude that “grazing is best used when addressing vegetation with stems of smaller diameters that make up the 1- and 10-hour fuels. These two fuel classes are important because they can greatly impact the rate of spread of a fire, as well as flame height” [Nader 2007: 19]. While they call for further research to validate anecdotal accounts supporting grazing and understand its best practice, they maintain that “prescribed grazing has the potential to be an ecologically and economically sustainable management tool for reduction of fuel loads” [Nader 2007: 20].
Landscape rehydration ‘better than dams’ in improving farm production, reducing fire risk, Major 2020
A project in Queensland, Australia has met with success in its efforts to rehydrate the landscape on the farmland property of Worona Station, improving biodiversity, water retention, and resistance to erosion and fire. Worona Station had been degraded and faced serious erosion issues, so Chris Le Feuvre, the owner, partnered with consultancy groups of NQ Dry Tropics and the Mulloon Institute in a project to rehydrate his land.
The project team has used a combination of planned grazing and small, low-tech dams to combat erosion problems. The grazing technique involves:
Splitting paddocks into small sizes and using large mobs of cattle grazing on rotation … grazing pasture more intensively while giving it longer to rest, [thereby] increasing carrying capacity.
Grazing in this way (which is evocative of Allan Savory’s Holistic Planned Grazing methodology) has resulted in increased pasture species diversity and boosted plant growth, allowing the Le Feuvre to double his herd size. Planned grazing has also reduced sediment runoff from the property. Sam Skeat, a grazing officer with NQ Dry Tropics, attests to the importance of grazing.
The plug-and-pond technique — also known as leaky weirs — involves small dam-like structures to lift the bed level of the water, which is then run onto the floodplain to grow pasture and recharge aquifers. While weirs have been strategically constructed, Mr. Skeat said grazing management was the most important tool to improve water retention in a landscape. ‘If you can use cattle as a tool to regenerate the grassland, you’ll get more infiltration, slow the flow, hold water up in the landscape and have you growing grass for longer’ [Major 2020].
Rehydrating landscapes can improve their resilience to extreme events, and improve their quality in the face of chronic problems like erosion. According to the Mulloon Institute Chairman Gary Nairn, the issue of degraded gullies and streams is a national concern. Gullies are created when parched land is unable to absorb rainwater, allowing it to run off. The sediment-filled runoff ends up in the ocean, polluting it.
Nairn sees land rehydration through planned grazing and related techniques as a better solution than building a massive, industrial-scale dam to retain water. The Australian government has been looking into building new large dams. Levels at Warragamba Dam, which supplies about 80 percent of Sydney’s water, have dropped to less than half capacity.
‘We’ve been able to demonstrate in Mulloon, if we repaired and rehydrated the catchment through to the Sydney water supply, you could store the equivalent of Warragamba Dam,’ he said [Major 2020].
Well-watered mulberry tree credited with saving home on NSW South Coast from summer bushfires, Aubrey 2020
A well-watered mulberry tree has been credited with averting the danger of destructive wildfires from destroying Brett Hawkins’ home during 2020’s unprecedented fire season in Australia. When massive fires raged through the bush through the summer, many homes were completely engulfed. However, Hawkins attested that when he returned to his home after evacuating,
‘I could see straight away the house was intact — the roof was intact, but everything else around it was burnt, with the exception of the mulberry tree.’ He described the stark scene greeting him upon arriving back home, ‘It was apocalyptic,’ Mr. Hawkins said. ‘There was not a tree left, ash on the ground and smouldering embers everywhere.’ But among the blackened trees, Mr. Hawkins found his mudbrick house and mulberry tree in full leaf [Aubrey 2020].
In the season’s drought, he had been rationing water, but sparing some to keep his tree hydrated and healthy, which may have been a contributing factor in its resistance. According to the article, “Mr. Hawkins believed that by heavily watering the tree, combined with luck regarding which direction the fires came, the full heat of the bushfires was shifted.” [Aubrey 2020]
Another important feature is the mulberry’s lack of dried leaves and brush at its base that might pose a danger of igniting. Other species, like eucalyptus, with oily leaves that could dry out, or pine trees whose long branches catch dry leaves, are less ideal.
A tree expert from the Fenner School of Environment at the Australian National University analyzed some of the possible factors leading to the survival of this mulberry tree and what it might teach those wishing to fortify the fire resilience of their homes and properties. “While there does not seem to be a clear answer on what to plant to ‘fire-proof’ your house, Professor Brack said a well-watered tree, with a clear trunk and no loose, dry leaves or branches is a good start” [Aubrey 2020].
Covid19 has been an additional stressor on many Native American communities already burdened by deprivations from centuries of ongoing injustice. According to Julie Garreau, project coordinator of Cheyenne River Youth Project, which operates a 2.5-acre youth garden in South Dakota, gardens are a source of both food and healing. “Gardens represent so much more,” she said. “Food, yes, but a belief in our future. Gardens represent resiliency, strength, wellness, culture.” During the pandemic, the Youth Project delivered garden produce and other foods to the homes of Cheyenne River Sioux Reservation children.
Another youth-focused gardening organization is Dream of Wild Health. Based in Minneapolis/St. Paul, MN, this Native-led organization operates a 30-acre biodiverse suburban farm that supplies food, learning experiences, and the chance to reconnect with nature. Kids learn cooking and seed saving, and student interns called Garden Warriors help grow food. Due to Covid19, workshops moved online, with the organization delivering ingredients to kids’ homes and then leading them in an online cooking class.
“Working in a garden develops your relationship to the land,” says Aubrey Skye, a Hunkpapa Lakota gardener who for many years ran a gardening program on Standing Rock Reservation on the border of North and South Dakota. “Our ancestors understood that. Look at the old pictures. It’s etched on their faces. When you understand it as well, a sense of scarcity and insecurity transforms into a feeling of abundance and control—something we all need these days.”
Some tips from the gardeners mentioned in this article:
Start small if you’re a beginner (in a few pots or a raised bed).
Favor companion planting. (“Look at nature, and figure out combinations that mimic it,” recommends Traditional Native American Farmers Association Director Clayton Brascoupé.)
Embellish your garden with colorful native flowers to attract and nourish pollinators.
Use rocks to keep crops cozy and supported; rocks act as heat sink and can protect seedlings from early frost.
Reuse discarded materials – you’ll get for free while building a network in the collection process: mulch with used cardboard and paper; create drip irrigation from soda pop bottles pierced with a needle at the neck, fill with water then bury the neck in the soil close to the plant.
Make compacted soil soft and plant friendly using dandelions, a supposed weed with nutritional value, whose taproot breaks up hardened soil enabling earthworm activity.
Include healing herbs, especially native varieties.
Save the seeds of the plants that thrive best and are favorites, which not only enables future food supply, but also, as Aubrey Skye says, preserves history like little time capsules.
In South Korea, knowledge of ancient farming techniques adapted to various harsh conditions, along with a sense of urgency about the need to adapt to even harsher conditions as the global climate system deteriorates, is bringing about the blossoming of an environmentally friendly agriculture movement.
Farmers draw on traditional knowledge of “nitrogen-fixing plants, soil bacteria, micro-organisms, and the relations between all of them to optimize yields by increasing soil fertility, boost crop health and biomass for livestock grazing, and reduce weed and pest infestations.” These practices are combined with intercropping (planting multiple crops together in a field) and crop rotation (constantly changing crops over time in a field) in a developing agricultural ethic that favors biodiversity and soil health.
Interestingly, the role of soil microorganisms is understood and valued in a way that intersects a fermentation-based food culture.
Traditional Korean knowledge of soil nutrients and food fermentation techniques is also used by some farmers to create natural fertilizer and pesticide. This is done by culturing and proliferating indigenous microorganisms – fungi, bacteria and yeast – to enhance the soil’s fertility without the need for livestock waste.
Such practices are supported both by national policy aiming to facilitate transition to organic and environmentally friendly methods, and by community-led organic farming movements. From participating in national climate strikes to demanding protections of native seeds to facilitating organic food commerce, consumer coops are doing their part to help make South Korea a global model for sustainable farming.
Similarly, both government and grassroots groups have established initiatives to recruit youth into agricultural careers.
The South Korean Ministry of Agriculture, Food and Rural Affairs has set up a Back-to-Earth Promotion Project, Youth Farmer Fostering Policy, and the Farmland Banking Project, aiming to promote and fund startups and businesses in the agricultural sector and in farming villages. …
Grassroots initiatives that are part of a similar movement can be seen in the Milmeori Farm School in Yeoju county and the Geumsan Gandhi School in Geumsan county. These are boarding school programs that bring youth from cities to experience the countryside, learn Korean organic farming, and cook plant-rich dishes from their harvests.
In Mingo County, West Virginia, the soil on a flat expanse of what had been a mountaintop is compacted, composed mainly of blasted rocks, and lacks organic matter, due to several years of coal mining. The ground is harder than anticipated; even the soil scientists say they are not sure how long it will take to bring the soil back to life. Besides, the ground does not retain water very well as it was engineered to drain water into the valley. Furthermore, there is the problem of aggressive invaders (autumn olive, multiflora rose, and tall fescue), making it difficult to penetrate the terrain.
As Ben Gilmer, president of Refresh Appalachia, which helps convert post-mine lands into agriculture and forestry enterprises, says, “it’s a long-term science project.” Refresh Appalachia provides job training and encourages farming systems that form a loop between the animals and plants, where one nourishes the other, cutting down on feed and fertilizer, providing food and land management, and helping ensure food sovereignty in an economically depressed region. Refresh farms raise poultry, goats, pigs, and honey bees, along with fruits, nuts, vegetables, and herbs.
Appalachia is a temperate region with heavy rainfall, not a barren moonscape. Each site being restored “just needs some care and management appropriate to their characteristics,” says Carl Zipper, Virginia Tech crop and soil science professor specializing in mine-land restoration.
The workers previously responsible for blowing up are now trying to put back together that which was blown up. Many are working on associate degrees in conjunction with job training in sustainable agriculture and related fields. “I’m living the dream,” Refresh member Wilburn Jude exclaimed. Former miner Chris Farley is excited to be part of the first group to attempt to farm these lands. Everyone was eager for the arrival of a mulcher to remove and chew the invasive shrubs into the wood chip. The clearing would then be planted with over 2,000 berry, pawpaw, and hazelnut seedlings.
In continuation of the “blessed unrest” section of previous issues of the Compendium, the following sketches illustrate how people everywhere are seeing that humanity depends on nature for both our physical and spiritual wellbeing and our survival. As this awareness takes hold, people act to protect and restore not only the land, but also our relationship to it. As the stories below show, growing food in an eco-friendly way does that. Adopting Paul Hawken’s terminology and characterization of “blessed unrest” as a spontaneous, decentralized global social movement, we here present a diverse series of vignettes of everyday heroes. May such stories light the fire for new heroes to perpetually emerge in defense of all life on Earth.
The hopeful work of turning Appalachia’s mountaintop coal mines into farms
In Mingo County, West Virginia, the soil on a flat expanse of what had been a mountaintop is compacted, composed mainly of blasted rocks, and lacks organic matter, due to several years of coal mining. The ground is harder than anticipated; even the soil scientists say they are not sure how long it will take to bring the soil back to life. Besides, the ground does not retain water very well as it was engineered to drain water into the valley. Furthermore, there is the problem of aggressive invaders (autumn olive, multiflora rose, and tall fescue), making it difficult to penetrate the terrain.
As Ben Gilmer, president of Refresh Appalachia, which helps convert post-mine lands into agriculture and forestry enterprises, says, “it’s a long-term science project.” Refresh Appalachia provides job training and encourages farming systems that form a loop between the animals and plants, where one nourishes the other, cutting down on feed and fertilizer, providing food and land management, and helping ensure food sovereignty in an economically depressed region. Refresh farms raise poultry, goats, pigs, and honey bees, along with fruits, nuts, vegetables, and herbs.
Appalachia is a temperate region with heavy rainfall, not a barren moonscape. Each site being restored “just needs some care and management appropriate to their characteristics,” says Carl Zipper, Virginia Tech crop and soil science professor specializing in mine-land restoration.
The workers previously responsible for blowing up are now trying to put back together that which was blown up. Many are working on associate degrees in conjunction with job training in sustainable agriculture and related fields. “I’m living the dream,” Refresh member Wilburn Jude exclaimed. Former miner Chris Farley is excited to be part of the first group to attempt to farm these lands. Everyone was eager for the arrival of a mulcher to remove and chew the invasive shrubs into the wood chip. The clearing would then be planted with over 2,000 berry, pawpaw, and hazelnut seedlings.
In South Korea, centuries of farming point to the future for sustainable agriculture
In South Korea, knowledge of ancient farming techniques adapted to various harsh conditions, along with a sense of urgency about the need to adapt to even harsher conditions as the global climate system deteriorates, is bringing about the blossoming of an environmentally friendly agriculture movement.
Farmers draw on traditional knowledge of “nitrogen-fixing plants, soil bacteria, micro-organisms, and the relations between all of them to optimize yields by increasing soil fertility, boost crop health and biomass for livestock grazing, and reduce weed and pest infestations.” These practices are combined with intercropping (planting multiple crops together in a field) and crop rotation (constantly changing crops over time in a field) in a developing agricultural ethic that favors biodiversity and soil health.
Interestingly, the role of soil microorganisms is understood and valued in a way that intersects a fermentation-based food culture.
Traditional Korean knowledge of soil nutrients and food fermentation techniques is also used by some farmers to create natural fertilizer and pesticide. This is done by culturing and proliferating indigenous microorganisms – fungi, bacteria and yeast – to enhance the soil’s fertility without the need for livestock waste.
Such practices are supported both by national policy aiming to facilitate transition to organic and environmentally friendly methods, and by community-led organic farming movements. From participating in national climate strikes to demanding protections of native seeds to facilitating organic food commerce, consumer coops are doing their part to help make South Korea a global model for sustainable farming.
Similarly, both government and grassroots groups have established initiatives to recruit youth into agricultural careers.
The South Korean Ministry of Agriculture, Food and Rural Affairs has set up a Back-to-Earth Promotion Project, Youth Farmer Fostering Policy, and the Farmland Banking Project, aiming to promote and fund startups and businesses in the agricultural sector and in farming villages. …
Grassroots initiatives that are part of a similar movement can be seen in the Milmeori Farm School in Yeoju county and the Geumsan Gandhi School in Geumsan county. These are boarding school programs that bring youth from cities to experience the countryside, learn Korean organic farming, and cook plant-rich dishes from their harvests.
Covid19 has been an additional stressor on many Native American communities already burdened by deprivations from centuries of ongoing injustice. According to Julie Garreau, project coordinator of Cheyenne River Youth Project, which operates a 2.5-acre youth garden in South Dakota, gardens are a source of both food and healing. “Gardens represent so much more,” she said. “Food, yes, but a belief in our future. Gardens represent resiliency, strength, wellness, culture.” During the pandemic, the Youth Project delivered garden produce and other foods to the homes of Cheyenne River Sioux Reservation children.
Another youth-focused gardening organization is Dream of Wild Health. Based in Minneapolis/St. Paul, MN, this Native-led organization operates a 30-acre biodiverse suburban farm that supplies food, learning experiences, and the chance to reconnect with nature. Kids learn cooking and seed saving, and student interns called Garden Warriors help grow food. Due to Covid19, workshops moved online, with the organization delivering ingredients to kids’ homes and then leading them in an online cooking class.
“Working in a garden develops your relationship to the land,” says Aubrey Skye, a Hunkpapa Lakota gardener who for many years ran a gardening program on Standing Rock Reservation on the border of North and South Dakota. “Our ancestors understood that. Look at the old pictures. It’s etched on their faces. When you understand it as well, a sense of scarcity and insecurity transforms into a feeling of abundance and control—something we all need these days.”
Some tips from the gardeners mentioned in this article:
Start small if you’re a beginner (in a few pots or a raised bed).
Favor companion planting. (“Look at nature, and figure out combinations that mimic it,” recommends Traditional Native American Farmers Association Director Clayton Brascoupé.)
Embellish your garden with colorful native flowers to attract and nourish pollinators.
Use rocks to keep crops cozy and supported; rocks act as heat sink and can protect seedlings from early frost.
Reuse discarded materials – you’ll get for free while building a network in the collection process: mulch with used cardboard and paper; create drip irrigation from soda pop bottles pierced with a needle at the neck, fill with water then bury the neck in the soil close to the plant.
Make compacted soil soft and plant friendly using dandelions, a supposed weed with nutritional value, whose taproot breaks up hardened soil enabling earthworm activity.
Include healing herbs, especially native varieties.
Save the seeds of the plants that thrive best and are favorites, which not only enables future food supply, but also, as Aubrey Skye says, preserves history like little time capsules.
While several studies have shown that biodiversity within a trophic level (among plants, for example) increases ecosystem function (such as productivity), this study examines the effects of increased plant diversity on multi-trophic networks (encompassing plants, soil microorganisms, and above- and belowground invertebrates). The authors compared monoculture plots (with one plant species) to plots containing 60 plant species, and found that:
higher plant diversity leads to more energy stored, greater energy flow and higher community-energy-use efficiency across the entire trophic network. These effects of biodiversity on energy dynamics were not restricted to only plants but were also expressed by other trophic groups and, to a similar degree, in aboveground and belowground parts of the ecosystem, even though plants are by far the dominating group in the system [Buzhdygan 2020: 1].
“More energy stored” means there is more standing biomass in the system, including plants, plant litter, microorganisms, insects and other invertebrates – in short, more life.
Compared to monoculture plots, high-diversity plots also had 50% greater energy flow, which implies “that the overall amount of resources consumed and recycled by the community increased with greater plant diversity” [Buzhdygan 2020: 2].
A community with “higher energy-use efficiency” has lower “maintenance costs,” referring to the amount of energy expended (through respiration) “to support the energetic demands of the living biomass stored in the system” [Buzhdygan 2020: 2]. In other words, organisms in an ecological community with high energy-use efficiency collectively work less hard to sustain themselves compared to, collectively, the organisms in a community with low energy-use efficiency. Biodiversity increases energy-use efficiency by increasing the quantity and variety of resources available to consumers.
Plant communities with a high plant diversity are typically more productive than low-diversity communities and, therefore, provide a larger quantity and variety of resources to consumers. This increase in resource availability can reduce competition and increase energy flow to consumers. A larger variety of resources can also attract a higher number of specialized species, supporting trophic complementarity across the network and resulting in a reduction of community maintenance costs [Buzhdygan 2020: 4].
In this way, higher energy-use efficiency boosts ecosystem function.
Higher energy use efficiency at high plant species richness may be an additional mechanism that contributes to the resilience of ecosystems because communities with low maintenance costs have a higher potential to compensate for energy loss during disturbance. … Moreover, lower community maintenance costs may imply a reduced ‘leakiness’ of ecosystems at high biodiversity. Indeed, evidence is mounting that high-biodiversity ecosystems lose less soil nitrogen, store more carbon in the soil and have more efficient soil microbial communities [Buzhdygan 2020: 7].
Inversely,
the reduced community-energy-use efficiency and standing stock biomass in species-poor ecosystems indicates that more carbon is released into the atmosphere; this implies potential feedback effects of the ongoing global biodiversity loss on carbon sequestration and climate change [Buzhdygan 2020: 8].
Biodiversity increases multitrophic energy use efficiency, flow and storage in grasslands, Buzhdygan 2020
While several studies have shown that biodiversity within a trophic level (among plants, for example) increases ecosystem function (such as productivity), this study examines the effects of increased plant diversity on multi-trophic networks (encompassing plants, soil microorganisms, and above- and belowground invertebrates). The authors compared monoculture plots (with one plant species) to plots containing 60 plant species, and found that:
higher plant diversity leads to more energy stored, greater energy flow and higher community-energy-use efficiency across the entire trophic network. These effects of biodiversity on energy dynamics were not restricted to only plants but were also expressed by other trophic groups and, to a similar degree, in aboveground and belowground parts of the ecosystem, even though plants are by far the dominating group in the system [Buzhdygan 2020: 1].
“More energy stored” means there is more standing biomass in the system, including plants, plant litter, microorganisms, insects and other invertebrates – in short, more life.
Compared to monoculture plots, high-diversity plots also had 50% greater energy flow, which implies “that the overall amount of resources consumed and recycled by the community increased with greater plant diversity” [Buzhdygan 2020: 2].
A community with “higher energy-use efficiency” has lower “maintenance costs,” referring to the amount of energy expended (through respiration) “to support the energetic demands of the living biomass stored in the system” [Buzhdygan 2020: 2]. In other words, organisms in an ecological community with high energy-use efficiency collectively work less hard to sustain themselves compared to, collectively, the organisms in a community with low energy-use efficiency. Biodiversity increases energy-use efficiency by increasing the quantity and variety of resources available to consumers.
Plant communities with a high plant diversity are typically more productive than low-diversity communities and, therefore, provide a larger quantity and variety of resources to consumers. This increase in resource availability can reduce competition and increase energy flow to consumers. A larger variety of resources can also attract a higher number of specialized species, supporting trophic complementarity across the network and resulting in a reduction of community maintenance costs [Buzhdygan 2020: 4].
In this way, higher energy-use efficiency boosts ecosystem function.
Higher energy use efficiency at high plant species richness may be an additional mechanism that contributes to the resilience of ecosystems because communities with low maintenance costs have a higher potential to compensate for energy loss during disturbance. … Moreover, lower community maintenance costs may imply a reduced ‘leakiness’ of ecosystems at high biodiversity. Indeed, evidence is mounting that high-biodiversity ecosystems lose less soil nitrogen, store more carbon in the soil and have more efficient soil microbial communities [Buzhdygan 2020: 7].
Inversely,
the reduced community-energy-use efficiency and standing stock biomass in species-poor ecosystems indicates that more carbon is released into the atmosphere; this implies potential feedback effects of the ongoing global biodiversity loss on carbon sequestration and climate change [Buzhdygan 2020: 8].
This article analyzes the policy context for forest ecosystem restoration, arguing that it is heavily shaped by the way we define a forest. The use of a forest definition lacking ecological considerations severely undermines conservation and restoration initiatives.
We live in an era of unprecedented environmental change, motivating equally unprecedented global actions to protect and restore forest ecosystems. These efforts could fail to achieve their ambitious goals if they are not informed by clear and appropriate concepts and definitions of forests [Chazdon 2016: 1].
There are multiple definitions of a forest. Early European and internationally adopted definitions tended to define forests according to their usage for timber. FAO’s 1948 definition created for assessing wood harvesting potential of the world’s forests is still in use today. Yet new definitions have since been created that emphasize conservation, carbon sequestration and biodiversity values of forests.
However, national and global forest assessments tend to use narrow technical definitions that ignore ecological values of forested land.
In many cases, forest assessments do not distinguish between land covered by natural and planted forests. Thus, if natural forests are cleared and replaced with plantations, no net loss of forest cover is reported [Chazdon 2016: 6].
In other words, areas that should not be considered forest in ecological terms are counted as forest – an obfuscation with disastrous environmental outcomes. Similarly, ecologically important yet small patches of trees that are not counted in forest inventories and lack legal protection are at risk of being lost.
Areas classified as ‘‘non-forests’’ are as important to forest definitions as are forests. More than 43 % of agricultural land globally is in agroforestry systems with 10 % tree cover. In Rwanda and Brazil, forest inventories using a 0.5-ha threshold ignore substantial areas of small forest fragments, agroforests, and woodlots, leading to underestimates of actual tree cover. Small patches of trees and even isolated remnant trees can hold high ecological and conservation value, and can play an important role in enhancing landscape connectivity, local biodiversity, and local livelihoods [Chazdon 2016: 7].
Information from participatory local monitoring and remote sensing technology that distinguishes “among successional stages of forests, selectively logged forests, and single-species plantations” [Chazdon 2016: 10] is needed.
Access to this information will allow countries and international agencies to track changes in natural forest cover, and to monitor processes of restoration, rehabilitation, and afforestation within a landscape context and, consequently, make informed policy decisions. We are on the frontier of developing new ways of monitoring and assessing land cover that will provide robust indicators of the quality and origins of tree cover and enable new ways of viewing and defining forests and reforests. To see beyond the overly simplified categories of forest loss, forest degradation, and forest gain, we need to develop and apply more adapted and nuanced definitions that will deepen our understanding of the drivers and outcomes of land-use change and forest dynamics within landscapes [Chazdon 2016: 10].
This study tested the Miyawaki method of rapid natural forest regeneration (which has been shown to work in Japan and elsewhere)in the arid Mediterranean. In this area, millennia of human civilization have resulted in degraded soils and reduced and changed forest cover, traditional reforestation efforts have often failed, and desertification is a looming threat. The Miyawaki method speeds up the process of ecological succession by densely planting a multilayer forest made up of a wide diversity of indigenous species.
In a natural forest cycle, as Clements (1916) described, annual plants on barren land are succeeded by perennial grass, sun-tolerant shrubs, light-demanding, fast-growing trees, and finally natural forests; each step may require decades, and the climax vegetation could be formed after two centuries or more. Currently, most forest reforestation programs adopt a scheme of planting one or more early successional species; after successful establishment, they are gradually replaced by intermediate species (either naturally or by planting), until late successional species arise. This pattern tries to simulate natural processes of ecological succession, from pioneer species to climax vegetation. However, it requires several silvicultural practices and normally takes a long time [Shirone 2011:82].
In the Miyawaki method, by contrast, one plants all at once the many plant species normally present in a native forest community, thus bypassing the earliest stages of ecological succession. Other tree-planting methods favor fast-growing non-native tree species, while omitting understory species – in other words, creating a simple plantation rather than a forest community that functions ecologically and can evolve and sustain itself.
Multilayer forests, and natural biocoenosis [ecological community] is possible, and well-developed ecosystems can be quickly established because of the simultaneous use of intermediate and late successional species in plantations. The Miyawaki method involves surveying the potential natural vegetation of the area to be reforested and recovering topsoil to a depth of 20– 30 cm by mixing the soil and a compost from organic materials, such as fallen leaves, mowed grass, etc. In this way, the time of the natural process of soil evolution, established by the vegetational succession itself, is reduced [Shirone 2011:82].
The authors of this study found that compared to traditional reforestation, there was “a more rapid development of trees on the Miyawaki plots, in particular, early-successional species [especially maritime pine]. The benefits over previous methods are remarkable and comparable with those obtained by Miyawaki in Asia and South America.” The Miyawaki method favoring denser plantings works even in arid climates, in spite of traditional views favoring sparse plantings in arid places, although the optimal density for the Mediterranean still needs testing, according to the authors.
In fact, low plant density has been traditionally retained as appropriate in arid and semiarid environments in order to avoid competition for water resources between plants, but it is now evident that cooperative processes, e.g., mutual shading, prevail over competitive processes. High plant density also reduces the impact of acorn predators, thus encouraging oak regeneration, i.e., the main late-successional forest species in Mediterranean environments. In addition, excellent plant stock remains fundamental for planting success in harsh environments [Shirone 2011:91].
This article assesses the ecological value of small woodlands relative to larger ones. The authors conclude that:
…smaller woodlands potentially deliver multiple services at higher performance levels on a per area basis than larger woodlands of a similar age, even if the larger woodlands harbor a higher biodiversity [Valdes 2020: 12].
Because of their high edge-to-core ratio, smaller woodlots get more sunlight and more nutrient input from surrounding farmland, resulting in denser vegetation cover and higher biomass production at edges.
This altered functioning in turn increases the delivery potential of some services, such as game production potential, due to an increased quantity of food available for game, and topsoil carbon storage, due to the faster incorporation of organic matter in the soil. Tick-borne disease risk is, however, lower, likely due to decreased larval densities in the unfavorable (e.g. hotter and drier) microclimatic conditions at the edge [Valdes 2020: 12].
While smaller woodlands were more apt to deliver “multiple services at higher levels of performance per area than larger woodlands of a similar age,” the greater biodiversity of larger woodlands increased certain individual ecosystem services.
The supply potential of several individual ecosystem services indirectly increased in larger and more ancient woodlands because it was dependent on higher levels of biodiversity. For example, abundance of usable plants and game production potential might have increased due to a positive correlation with vascular plant diversity, while pest control potential probably increased due to bottom-up effects through the trophic chain. On the contrary, tick-borne disease risk, topsoil carbon storage and stemwood volume were unrelated to multidiversity, probably because they depended on particular environmental conditions or on the presence and abundance of specific species rather than on species richness per se.
Finally, it should be noted that we focused on the service delivery potential on a per area basis and that the total amount of services provided by large patches might still be larger than that of small patches. Our findings should therefore not be interpreted as a trade-off between large, biodiverse patches versus small patches that have a higher potential to deliver services, but rather as an observation that small woodlands in agricultural landscapes have the potential to deliver a high flow of services relative to their size [Valdes 2020: 12].
This study tested biodiversity effects on ecosystem function in the process of reviving severely degraded and contaminated land, and found that “increasing plant diversity greatly enhanced the reclamation of these lands” [Jia 2020: 1].
Prior to implementing the reclamation experiment, the degraded mine wasteland investigated in this study was heavily impacted by past mining activities and was devoid of vegetation for more than a decade and the soil lacked structure, contained high levels of toxic metals and low levels of nutrients. … our results showed that higher plant species richness enhanced land reclamation across all standard measures of reclamation success and specifically resulted in higher vegetation coverage, biomass yield and stability for all 3 years [of the experiment] [Jia 2020: 6].
Furthermore, higher biodiversity plots had higher levels of organic carbon in the soil, higher soil microorganism abundance, lower fungal pathogens, and lower heavy metal concentrations in plant tissue.
The most striking impact of plant diversity on soil was on the microbial communities. Both soil fungal and bacterial OTUs [operational taxonomic units[8]] increased significantly with plant species richness. More importantly, we found that higher plant species richness significantly increased the relative abundance of soil cellulolytic bacteria that degrade cellulose and are thus essential components of nutrient cycling [Jia 2020: 7].
The concept of “proforestation” presented here means letting existing forests continue to grow and reach their full ecological potential. Due to intensive management practices, most existing forests sequester carbon at only half (or less) of their potential rate. In addition to storing (embodying) more carbon than their smaller counterparts, large trees also sequester carbon at a faster rate. For example, “Each year a single tree that is 100 cm in diameter adds the equivalent biomass of an entire 10-20 cm diameter tree, further underscoring the role of large trees” [Moomaw 2019: 4]. Imagine, reader, that every year you planted a whole new medium-sized tree – that’s essentially what large trees are doing.
“Each year a single tree that is 100 cm in diameter adds the equivalent biomass of an entire 10-20 cm diameter tree, further underscoring the role of large trees” [Moomaw 2019: 4].
Much of Maine’s forests have been harvested continuously for 200 years and have a carbon density less than one-third of the forests of Southern Vermont and New Hampshire, Northwestern Connecticut and Western Massachusetts – a region that has not been significantly harvested over the past 75-150 years. …
Ecosystem services accrue as forests age for centuries. Far from plateauing in terms of carbon sequestration (or added wood) at a relatively young age as was long believed, older forests (e.g., >200 years of age without intervention) contain a variety of habitats, typically continue to sequester additional carbon for many decades or even centuries, and sequester significantly more carbon than younger and managed stands [Moomaw 2019: 5].
Because existing forests are already sequestering carbon, and will continue at an increasing rate as tree size grows, the author argues that proforestation is a more effective immediate solution than either reforestation (planting new trees where they had been cleared for agriculture, etc.) or afforestation (planting trees in new places), though these other two approaches are important for longer term ecosystem function. Moomaw et al. argue that the urgency of removing CO2makes it imperative to keep existing forests growing.
Globally, existing forests only store approximately half of their potential due to past and present management, and many existing forests are capable of immediate and even more extensive growth for many decades. During the timeframe while seedlings planted for afforestation and reforestation are growing (yet will never achieve the carbon density of an intact forest), proforestation is a safe, highly effective, immediate natural solution that does not rely on uncertain discounted future benefits inherent in other options [Moomaw 2019: 7].
Furthermore, existing, older forests are critical habitats for threatened wildlife, even small intact woods.
Forest bird guilds also benefit from small intact forests in urban landscapes relative to unprotected matrix forests. Several bird species in the U.S. that are globally threatened – including the wood thrush, cerulean warbler, marbled murrelet, and spotted owl are, in part, dependent on intact, older forests with large trees [Moomaw 2019: 5].
In sum, proforestation provides the most effective solution to dual global crises – climate change and biodiversity loss. It is the only practical, rapid, economical, and effective means for atmospheric CDR [carbon dioxide removal] among the multiple options that have been proposed because it removes more atmospheric carbon dioxide in the immediate future and continues to sequester it long-term. Proforestation will increase the diversity of many groups of organisms and provide numerous additional and important ecosystem services. While multiple strategies will be needed to address global environmental crises, proforestation is a very low-cost option for increasing carbon sequestration that does not require additional land beyond what is already forested and provides new forest related jobs and opportunities along with a wide array of quantifiable ecosystem services, including human health [Moomaw 2019: 8].
This perspective piece argues against scientific or public adoption of the term “rewilding,” which the authors view as being generally synonymous with the classical and better-understood concept of ecological restoration. Definitions of restoration are sufficient to encompass practices espoused in rewilding.
Early definitions of restoration describe the practice as “the process of repairing damage caused by humans to the diversity and dynamics of indigenous ecosystems” (Jackson et al., 1995). Although at the time of this definition, restoration science was still developing, it was clear that it had established itself under the broad banner of repairing damaged ecosystems. … More recently, restoration has been defined as “any activity whose aim it is to ultimately achieve ecosystem recovery, insofar as possible and relative to an appropriate local native model (termed here a reference ecosystem), regardless of the period of time required to achieve the recovery outcome” (McDonald et al., 2016) [Hayward 2019: 257].
The term rewilding, which has evolved over time, “was arguably conceived to promote the original authors’ view of conservation via cores [habitats], corridors, and carnivores” [Hayward 2019: 256]. In this early context,
‘rewilding’ referred to conservation and management interventions that focused on reintroducing keystone predators and ensuring that they had sufficient interconnected space to live. The authors emphasized within their original work that rewilding was “one essential element in most efforts to restore fully functioning ecosystems” (Soulé and Noss, 1998). As such, it is clear that rewilding was originally aimed to be a term that referred to one component of ecological restoration [Hayward 2019: 256].
Since then, rewilding has come to refer to practices involving “translocating substitute species to fill vacant ecological niches left by extinct species” [Hayward 2019: 257] or reintroducing locally extinct species, or simply allowing natural succession to occur on abandoned land. Given multiple definitions, all of which relate to the idea of restoration, the term rewilding is seen here as superfluous and confusing.
Given the lack of clear differences between rewilding and restoration in both definition and practice, we see little need for these competing terms within scientific discourse [Hayward 2019: 257].
However, the authors suggest two positive contributions from the rewilding discourse. Because of its overall focus on large fauna, rewilding has captured public imagination and interest in conservation, while also helping to shift a potential vegetation bias among restoration practitioners/scientists toward equal emphasis on the ecosystem role of animals.
Therefore, rather than adopting a new term with copious definitions that lack clarity, this debate can be used as an opportunity to adaptively improve current restoration practice by incorporating a more equal focus between flora and fauna [Hayward 2019: 258].
Because of its overall focus on large fauna, rewilding has captured public imagination and interest in conservation, while also helping to shift a potential vegetation bias among restoration practitioners/scientists toward equal emphasis on the ecosystem role of animals.
This commentary distinguishes between restoration and rewilding of ecosystems, explaining that the latter aims at ecological adaptation to novel local environmental conditions wrought by global climate change. By contrast, restoration, as defined here, aims to recreate and maintain an historical state or condition of an ecosystem, regardless of current environmental conditions.
Although the two words are often conflated,
Rewilding is thus conceptually different from restoring. It is an adaptive approach to conserving ecological functionality under changing environmental conditions, to which historical benchmarks are less relevant than to restoring. It inherently acknowledges and promotes unpredictability, while placing the emphasis on function over species composition [du Toit & Pettorelli 2019: 2].
The authors assert that rewilding is better suited to preserving biodiversity and ecosystem function under present and future conditions.
It is difficult to imagine how conserving biodiversity and ecosystem services could be possible in predicted future scenarios without rewilding. Simply stated, anthropogenic environmental forcing makes ecosystem restoration a diminishing option [du Toit & Pettorelli 2019: 4].
Rewilding is gaining traction as an approach to conservation. However, many different perspectives about which species and ecological processes to focus rewilding efforts on and how deeply to intervene in systems has created some confusion and contention within the field. Furthermore, the most ambitious and extreme rewilding proposals (for example, recreating communities that went extinct millennia ago) have often attracted more attention, while the more pragmatic and immediate solutions in the field are overlooked.
This article attempts to clarify the concept. The authors emphasize that rewilding is a process-oriented approach to biodiversity conservation “focused on preserving and restoring the structural and functional complexity of degraded ecosystems” [Fernandez 2017: 276].
Rewilding pursues the goal of restoring wild species interactions and their regulation of key ecosystem processes including nutrient and energy flows, vegetation succession and disturbances, drawing specific attention to the key roles of large-bodied species that are especially sensitive to the human appropriation of landscapes [Fernandez 2017: 277].
The authors suggest further research is needed. They note that while the negative effects on ecosystems of the loss of biodiversity and keystone species is well documented, ecosystem responses to species reintroduction and other rewilding efforts are not as well studied. To guide future research, the authors
propose an unequivocally process-oriented formulation of the “rewilding hypothesis” as a general guidance: that the large-scale restoration of apex consumers and large herbivores promotes self-regulation in community assemblages, and increases the complexity of ecological processes in ecosystems [Fernandez 2016: 277].
Also needed is policy and management practice support, particularly in terms of protecting the areas and species in question. Proactive policies could ensure that gains made toward ecological restoration are not undermined by damaging human activity.
Policies and practices should be developed in order to enforce the idea that rewilding is about reducing the human control on ecosystem processes. It must begin to include varied objectives to alleviate pressures on wildlife populations such as a full legal protection of large predators based on their unique ecological roles and not just depending on their conservation status; the eradication of predator control programs; or the elimination of game management practices such as wildlife fencing, introduction of alien game populations, supplementary feeding and others that profoundly alter the natural regulation and the genetic structure of large herbivore populations [Fernandez 2016: 278].
A growing body of literature emphasizes the need for novel, process-oriented approaches to restoring ecosystems in our rapidly changing world. Dynamic and process-oriented approaches focus on the adaptive capacity of ecosystems and the restoration of ecosystem processes promoting biodiversity, rather than aiming to maintain or restore particular ecosystem states characterized by predefined species compositions or particular bundles of ecosystem services [Perino 2019: 1].
In contrast to other types of restoration efforts aiming to recreate the composition and appearance of an historical ecological community, rewilding focuses on ecosystem function and recognizes the dynamic and unpredictable nature of ecosystems. This article highlights three key ecological processes that rewilding aims to activate: trophic complexity, natural disturbances and dispersal.
Rewilding aims to restore these three ecological processes to foster complex and self-organizing ecosystems that require minimum human management in the long term [Perino 2019: 2].
Trophic complexity implies the presence of large vertebrates, including herbivores that modify the landscape through grazing or dam building and predators that control the herbivore populations. These keystone species can promote biodiversity in the landscapes they inhabit.
Stochastic (random) natural disturbance (such as fire or flooding) can increase ecosystem heterogeneity and complexity, allowing less competitive species to survive. Rewilding involves discontinuing both controlled anthropogenic disturbances and suppression of natural disturbances.
Rewilding actions aim to release ecosystems from continued and controlled anthropogenic disturbances to allow for natural variability and sources of stochasticity. Mowing of grassland can be reduced or replaced by natural grazing. Dams can be removed or their management modified to restore natural flood regimes. Logging can be replaced by allowing natural fire and pest regimes [Perino 2019: 4].
Dispersal– rewilding aims to remove anthropogenic barriers that limit the movement of plants and animals and thus the dispersal of their genetic material and potential for recolonization after a disturbance event. The creation of ecological corridors is an example of a rewilding activity that enhances dispersal.
The interaction of these ecological processes boosts the functioning of each. For example, the presence of larger animals facilitates seed dispersal throughout the system. High levels of dispersal, in turn, can facilitate ecosystem recovery following a disturbance.
Rewilding projects can be passive (allowing abandoned agricultural fields to recover on their own) or active (species reintroductions, for example), and are most effective when conducted in a manner that engages the local community in the process.
This meta-analysis of 400 studies compared passive and active ecosystem repair outcomes in terms of the speed and completeness of recovery, and found little difference between the two approaches.
Active restoration did not result in faster or more complete recovery than simply ending the disturbances ecosystems face [Jones 2018: 1].
Passive recovery simply means ending the anthropogenic disturbance that was causing the degradation, while active restoration here includes anything from fertilizer application to recontouring/dredging to planting a desired species mix.
The authors speculate that the lack of different outcomes between the two approaches could be due to restoration managers correctly choosing to actively restore the ecosystems that “are not recovering on their own and require active restoration to improve recovery outcomes relative to passively recovering systems” [Jones 2018: 6]. Also, the actively managed sites in the study had, on average, less time to recover than the passively managed sites. Finally, the authors suggest there may be right and wrong ways to actively restore ecosystems, and “recommend that restoration strategies be tailored more closely to overcome the specific barriers to recovery in individual sites” [Jones 2018: 6].
Assuming active and passive restoration achieve comparable outcomes in many cases, then passive restoration deserves serious consideration, given limited resources available for the vast amount of ecosystem repair required in the world today.
Letting ecosystems repair themselves in many cases may be the most effective restoration strategy – a counterintuitive yet critical finding that could help society allocate restoration funds more efficiently in the future [Jones 2018: 6].
The study also consistently found that across systems, ecosystems didn’t fully recover, at least not within the timeframe of the studies.
Our results expand those findings to a broader range of ecosystems and geographies, and, together with previous work, suggest the majority of ecosystems have not yet recovered fully following disturbance and may not in the future. Thus, restoration should not be considered a substitute for conservation, which is a key strategy to ensure sustained support of biodiversity and delivery of ecosystem services in the future [Jones 2018: 4].
This meta-analysis comparing active restoration to natural ecosystem regeneration found the latter to be more effective. The authors conclude that “lower-cost natural regeneration surpasses active restoration in achieving tropical forest restoration success for biodiversity and vegetation structure[7]” [Crouzeilles 2017: 4]. This conclusion runs counter to conventional wisdom that active restoration is preferable despite being more expensive.
Natural forest regeneration is the spontaneous recovery of native tree species that colonize and establish in abandoned fields or natural disturbances; this process can also be assisted through human interventions such as fencing to control livestock grazing, weed control, and fire protection. In contrast, active restoration requires planting of nursery-grown seedlings, direct seeding, and/or the manipulation of disturbance regimes (for example, thinning and burning) to speed up the recovery process, often at a high cost to establish structural features of the vegetation (hereafter termed vegetation structure), reassemble local species composition, and/or catalyze ecological succession [Crouzeilles 2017: 1].
However, “restoration success for biodiversity and vegetation structure was significantly lower in both natural regeneration and active restoration than in reference systems” [Crouzeilles 2017: 2], underscoring the importance of conserving existing intact ecosystems.
“Restoration success for biodiversity and vegetation structure was significantly lower in both natural regeneration and active restoration than in reference systems” [Crouzeilles 2017: 2], underscoring the importance of conserving existing intact ecosystems.
Part of the explanation for the lower success of active restoration compared to natural regeneration is that the composition and/or diversity of species chosen for planting in active restoration may be inappropriate, while the species that colonize abandoned land are likely to be diverse and locally adapted.
Natural regeneration is initiated through the colonization of opportunistic and locally adapted species, resulting in a stochastic dynamic process of forest restoration that ultimately leads to higher diversity of native, locally adapted plant species than in tree planting schemes (that is, active restoration). Active restoration also can create a highly diverse habitat through human introduction of up to 6000 seedlings/ha, but tree species used in plantings often lack the full range of functional traits found in natural regrowth forests. In addition, most tropical forest plantings for restoration or forest plantations use relatively few species, that is, these plantations may not be planted primarily for biodiversity outcomes. Thus, the higher plant biodiversity in naturally regenerated systems creates more habitats and resources, which provide additional sources of food, shelter, nesting, and breeding sites, to support higher animal biodiversity [Crouzeilles 2017: 2].
Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests, Crouzeilles et al. 2017
This meta-analysis comparing active restoration to natural ecosystem regeneration found the latter to be more effective. The authors conclude that “lower-cost natural regeneration surpasses active restoration in achieving tropical forest restoration success for biodiversity and vegetation structure[7]” [Crouzeilles 2017: 4]. This conclusion runs counter to conventional wisdom that active restoration is preferable despite being more expensive.
Natural forest regeneration is the spontaneous recovery of native tree species that colonize and establish in abandoned fields or natural disturbances; this process can also be assisted through human interventions such as fencing to control livestock grazing, weed control, and fire protection. In contrast, active restoration requires planting of nursery-grown seedlings, direct seeding, and/or the manipulation of disturbance regimes (for example, thinning and burning) to speed up the recovery process, often at a high cost to establish structural features of the vegetation (hereafter termed vegetation structure), reassemble local species composition, and/or catalyze ecological succession [Crouzeilles 2017: 1].
However, “restoration success for biodiversity and vegetation structure was significantly lower in both natural regeneration and active restoration than in reference systems” [Crouzeilles 2017: 2], underscoring the importance of conserving existing intact ecosystems.
“Restoration success for biodiversity and vegetation structure was significantly lower in both natural regeneration and active restoration than in reference systems” [Crouzeilles 2017: 2], underscoring the importance of conserving existing intact ecosystems.
Part of the explanation for the lower success of active restoration compared to natural regeneration is that the composition and/or diversity of species chosen for planting in active restoration may be inappropriate, while the species that colonize abandoned land are likely to be diverse and locally adapted.
Natural regeneration is initiated through the colonization of opportunistic and locally adapted species, resulting in a stochastic dynamic process of forest restoration that ultimately leads to higher diversity of native, locally adapted plant species than in tree planting schemes (that is, active restoration). Active restoration also can create a highly diverse habitat through human introduction of up to 6000 seedlings/ha, but tree species used in plantings often lack the full range of functional traits found in natural regrowth forests. In addition, most tropical forest plantings for restoration or forest plantations use relatively few species, that is, these plantations may not be planted primarily for biodiversity outcomes. Thus, the higher plant biodiversity in naturally regenerated systems creates more habitats and resources, which provide additional sources of food, shelter, nesting, and breeding sites, to support higher animal biodiversity [Crouzeilles 2017: 2].
Restoration and repair of Earth’s damaged ecosystems, Jones et al. 2018
This meta-analysis of 400 studies compared passive and active ecosystem repair outcomes in terms of the speed and completeness of recovery, and found little difference between the two approaches.
Active restoration did not result in faster or more complete recovery than simply ending the disturbances ecosystems face [Jones 2018: 1].
Passive recovery simply means ending the anthropogenic disturbance that was causing the degradation, while active restoration here includes anything from fertilizer application to recontouring/dredging to planting a desired species mix.
The authors speculate that the lack of different outcomes between the two approaches could be due to restoration managers correctly choosing to actively restore the ecosystems that “are not recovering on their own and require active restoration to improve recovery outcomes relative to passively recovering systems” [Jones 2018: 6]. Also, the actively managed sites in the study had, on average, less time to recover than the passively managed sites. Finally, the authors suggest there may be right and wrong ways to actively restore ecosystems, and “recommend that restoration strategies be tailored more closely to overcome the specific barriers to recovery in individual sites” [Jones 2018: 6].
Assuming active and passive restoration achieve comparable outcomes in many cases, then passive restoration deserves serious consideration, given limited resources available for the vast amount of ecosystem repair required in the world today.
Letting ecosystems repair themselves in many cases may be the most effective restoration strategy – a counterintuitive yet critical finding that could help society allocate restoration funds more efficiently in the future [Jones 2018: 6].
The study also consistently found that across systems, ecosystems didn’t fully recover, at least not within the timeframe of the studies.
Our results expand those findings to a broader range of ecosystems and geographies, and, together with previous work, suggest the majority of ecosystems have not yet recovered fully following disturbance and may not in the future. Thus, restoration should not be considered a substitute for conservation, which is a key strategy to ensure sustained support of biodiversity and delivery of ecosystem services in the future [Jones 2018: 4].
Rewilding complex ecosystems, Perino et al. 2019
A growing body of literature emphasizes the need for novel, process-oriented approaches to restoring ecosystems in our rapidly changing world. Dynamic and process-oriented approaches focus on the adaptive capacity of ecosystems and the restoration of ecosystem processes promoting biodiversity, rather than aiming to maintain or restore particular ecosystem states characterized by predefined species compositions or particular bundles of ecosystem services [Perino 2019: 1].
In contrast to other types of restoration efforts aiming to recreate the composition and appearance of an historical ecological community, rewilding focuses on ecosystem function and recognizes the dynamic and unpredictable nature of ecosystems. This article highlights three key ecological processes that rewilding aims to activate: trophic complexity, natural disturbances and dispersal.
Rewilding aims to restore these three ecological processes to foster complex and self-organizing ecosystems that require minimum human management in the long term [Perino 2019: 2].
Trophic complexity implies the presence of large vertebrates, including herbivores that modify the landscape through grazing or dam building and predators that control the herbivore populations. These keystone species can promote biodiversity in the landscapes they inhabit.
Stochastic (random) natural disturbance (such as fire or flooding) can increase ecosystem heterogeneity and complexity, allowing less competitive species to survive. Rewilding involves discontinuing both controlled anthropogenic disturbances and suppression of natural disturbances.
Rewilding actions aim to release ecosystems from continued and controlled anthropogenic disturbances to allow for natural variability and sources of stochasticity. Mowing of grassland can be reduced or replaced by natural grazing. Dams can be removed or their management modified to restore natural flood regimes. Logging can be replaced by allowing natural fire and pest regimes [Perino 2019: 4].
Dispersal– rewilding aims to remove anthropogenic barriers that limit the movement of plants and animals and thus the dispersal of their genetic material and potential for recolonization after a disturbance event. The creation of ecological corridors is an example of a rewilding activity that enhances dispersal.
The interaction of these ecological processes boosts the functioning of each. For example, the presence of larger animals facilitates seed dispersal throughout the system. High levels of dispersal, in turn, can facilitate ecosystem recovery following a disturbance.
Rewilding projects can be passive (allowing abandoned agricultural fields to recover on their own) or active (species reintroductions, for example), and are most effective when conducted in a manner that engages the local community in the process.
Rewilding: a call for boosting ecological complexity in conservation, Fernández et al. 2017
Rewilding is gaining traction as an approach to conservation. However, many different perspectives about which species and ecological processes to focus rewilding efforts on and how deeply to intervene in systems has created some confusion and contention within the field. Furthermore, the most ambitious and extreme rewilding proposals (for example, recreating communities that went extinct millennia ago) have often attracted more attention, while the more pragmatic and immediate solutions in the field are overlooked.
This article attempts to clarify the concept. The authors emphasize that rewilding is a process-oriented approach to biodiversity conservation “focused on preserving and restoring the structural and functional complexity of degraded ecosystems” [Fernandez 2017: 276].
Rewilding pursues the goal of restoring wild species interactions and their regulation of key ecosystem processes including nutrient and energy flows, vegetation succession and disturbances, drawing specific attention to the key roles of large-bodied species that are especially sensitive to the human appropriation of landscapes [Fernandez 2017: 277].
The authors suggest further research is needed. They note that while the negative effects on ecosystems of the loss of biodiversity and keystone species is well documented, ecosystem responses to species reintroduction and other rewilding efforts are not as well studied. To guide future research, the authors
propose an unequivocally process-oriented formulation of the “rewilding hypothesis” as a general guidance: that the large-scale restoration of apex consumers and large herbivores promotes self-regulation in community assemblages, and increases the complexity of ecological processes in ecosystems [Fernandez 2016: 277].
Also needed is policy and management practice support, particularly in terms of protecting the areas and species in question. Proactive policies could ensure that gains made toward ecological restoration are not undermined by damaging human activity.
Policies and practices should be developed in order to enforce the idea that rewilding is about reducing the human control on ecosystem processes. It must begin to include varied objectives to alleviate pressures on wildlife populations such as a full legal protection of large predators based on their unique ecological roles and not just depending on their conservation status; the eradication of predator control programs; or the elimination of game management practices such as wildlife fencing, introduction of alien game populations, supplementary feeding and others that profoundly alter the natural regulation and the genetic structure of large herbivore populations [Fernandez 2016: 278].
The differences between rewilding and restoring an ecologically degraded landscape, du Toit & Pettorelli 2019
This commentary distinguishes between restoration and rewilding of ecosystems, explaining that the latter aims at ecological adaptation to novel local environmental conditions wrought by global climate change. By contrast, restoration, as defined here, aims to recreate and maintain an historical state or condition of an ecosystem, regardless of current environmental conditions.
Although the two words are often conflated,
Rewilding is thus conceptually different from restoring. It is an adaptive approach to conserving ecological functionality under changing environmental conditions, to which historical benchmarks are less relevant than to restoring. It inherently acknowledges and promotes unpredictability, while placing the emphasis on function over species composition [du Toit & Pettorelli 2019: 2].
The authors assert that rewilding is better suited to preserving biodiversity and ecosystem function under present and future conditions.
It is difficult to imagine how conserving biodiversity and ecosystem services could be possible in predicted future scenarios without rewilding. Simply stated, anthropogenic environmental forcing makes ecosystem restoration a diminishing option [du Toit & Pettorelli 2019: 4].
Reintroducing rewilding to restoration – rejecting the search for novelty, Hayward et al. 2019
This perspective piece argues against scientific or public adoption of the term “rewilding,” which the authors view as being generally synonymous with the classical and better-understood concept of ecological restoration. Definitions of restoration are sufficient to encompass practices espoused in rewilding.
Early definitions of restoration describe the practice as “the process of repairing damage caused by humans to the diversity and dynamics of indigenous ecosystems” (Jackson et al., 1995). Although at the time of this definition, restoration science was still developing, it was clear that it had established itself under the broad banner of repairing damaged ecosystems. … More recently, restoration has been defined as “any activity whose aim it is to ultimately achieve ecosystem recovery, insofar as possible and relative to an appropriate local native model (termed here a reference ecosystem), regardless of the period of time required to achieve the recovery outcome” (McDonald et al., 2016) [Hayward 2019: 257].
The term rewilding, which has evolved over time, “was arguably conceived to promote the original authors’ view of conservation via cores [habitats], corridors, and carnivores” [Hayward 2019: 256]. In this early context,
‘rewilding’ referred to conservation and management interventions that focused on reintroducing keystone predators and ensuring that they had sufficient interconnected space to live. The authors emphasized within their original work that rewilding was “one essential element in most efforts to restore fully functioning ecosystems” (Soulé and Noss, 1998). As such, it is clear that rewilding was originally aimed to be a term that referred to one component of ecological restoration [Hayward 2019: 256].
Since then, rewilding has come to refer to practices involving “translocating substitute species to fill vacant ecological niches left by extinct species” [Hayward 2019: 257] or reintroducing locally extinct species, or simply allowing natural succession to occur on abandoned land. Given multiple definitions, all of which relate to the idea of restoration, the term rewilding is seen here as superfluous and confusing.
Given the lack of clear differences between rewilding and restoration in both definition and practice, we see little need for these competing terms within scientific discourse [Hayward 2019: 257].
However, the authors suggest two positive contributions from the rewilding discourse. Because of its overall focus on large fauna, rewilding has captured public imagination and interest in conservation, while also helping to shift a potential vegetation bias among restoration practitioners/scientists toward equal emphasis on the ecosystem role of animals.
Therefore, rather than adopting a new term with copious definitions that lack clarity, this debate can be used as an opportunity to adaptively improve current restoration practice by incorporating a more equal focus between flora and fauna [Hayward 2019: 258].
Because of its overall focus on large fauna, rewilding has captured public imagination and interest in conservation, while also helping to shift a potential vegetation bias among restoration practitioners/scientists toward equal emphasis on the ecosystem role of animals.
Intact forests in the United States: proforestation mitigates climate change and serves the greatest good, Moomaw 2019
The concept of “proforestation” presented here means letting existing forests continue to grow and reach their full ecological potential. Due to intensive management practices, most existing forests sequester carbon at only half (or less) of their potential rate. In addition to storing (embodying) more carbon than their smaller counterparts, large trees also sequester carbon at a faster rate. For example, “Each year a single tree that is 100 cm in diameter adds the equivalent biomass of an entire 10-20 cm diameter tree, further underscoring the role of large trees” [Moomaw 2019: 4]. Imagine, reader, that every year you planted a whole new medium-sized tree – that’s essentially what large trees are doing.
“Each year a single tree that is 100 cm in diameter adds the equivalent biomass of an entire 10-20 cm diameter tree, further underscoring the role of large trees” [Moomaw 2019: 4].
Much of Maine’s forests have been harvested continuously for 200 years and have a carbon density less than one-third of the forests of Southern Vermont and New Hampshire, Northwestern Connecticut and Western Massachusetts – a region that has not been significantly harvested over the past 75-150 years. …
Ecosystem services accrue as forests age for centuries. Far from plateauing in terms of carbon sequestration (or added wood) at a relatively young age as was long believed, older forests (e.g., >200 years of age without intervention) contain a variety of habitats, typically continue to sequester additional carbon for many decades or even centuries, and sequester significantly more carbon than younger and managed stands [Moomaw 2019: 5].
Because existing forests are already sequestering carbon, and will continue at an increasing rate as tree size grows, the author argues that proforestation is a more effective immediate solution than either reforestation (planting new trees where they had been cleared for agriculture, etc.) or afforestation (planting trees in new places), though these other two approaches are important for longer term ecosystem function. Moomaw et al. argue that the urgency of removing CO2makes it imperative to keep existing forests growing.
Globally, existing forests only store approximately half of their potential due to past and present management, and many existing forests are capable of immediate and even more extensive growth for many decades. During the timeframe while seedlings planted for afforestation and reforestation are growing (yet will never achieve the carbon density of an intact forest), proforestation is a safe, highly effective, immediate natural solution that does not rely on uncertain discounted future benefits inherent in other options [Moomaw 2019: 7].
Furthermore, existing, older forests are critical habitats for threatened wildlife, even small intact woods.
Forest bird guilds also benefit from small intact forests in urban landscapes relative to unprotected matrix forests. Several bird species in the U.S. that are globally threatened – including the wood thrush, cerulean warbler, marbled murrelet, and spotted owl are, in part, dependent on intact, older forests with large trees [Moomaw 2019: 5].
In sum, proforestation provides the most effective solution to dual global crises – climate change and biodiversity loss. It is the only practical, rapid, economical, and effective means for atmospheric CDR [carbon dioxide removal] among the multiple options that have been proposed because it removes more atmospheric carbon dioxide in the immediate future and continues to sequester it long-term. Proforestation will increase the diversity of many groups of organisms and provide numerous additional and important ecosystem services. While multiple strategies will be needed to address global environmental crises, proforestation is a very low-cost option for increasing carbon sequestration that does not require additional land beyond what is already forested and provides new forest related jobs and opportunities along with a wide array of quantifiable ecosystem services, including human health [Moomaw 2019: 8].
Plant diversity enhances the reclamation of degraded lands by stimulating plant-soil feedbacks, Jia et al. 2020
This study tested biodiversity effects on ecosystem function in the process of reviving severely degraded and contaminated land, and found that “increasing plant diversity greatly enhanced the reclamation of these lands” [Jia 2020: 1].
Prior to implementing the reclamation experiment, the degraded mine wasteland investigated in this study was heavily impacted by past mining activities and was devoid of vegetation for more than a decade and the soil lacked structure, contained high levels of toxic metals and low levels of nutrients. … our results showed that higher plant species richness enhanced land reclamation across all standard measures of reclamation success and specifically resulted in higher vegetation coverage, biomass yield and stability for all 3 years [of the experiment] [Jia 2020: 6].
Furthermore, higher biodiversity plots had higher levels of organic carbon in the soil, higher soil microorganism abundance, lower fungal pathogens, and lower heavy metal concentrations in plant tissue.
The most striking impact of plant diversity on soil was on the microbial communities. Both soil fungal and bacterial OTUs [operational taxonomic units[8]] increased significantly with plant species richness. More importantly, we found that higher plant species richness significantly increased the relative abundance of soil cellulolytic bacteria that degrade cellulose and are thus essential components of nutrient cycling [Jia 2020: 7].
High ecosystem service delivery potential of small woodlands in agricultural landscapes, Valdes 2020
This article assesses the ecological value of small woodlands relative to larger ones. The authors conclude that:
…smaller woodlands potentially deliver multiple services at higher performance levels on a per area basis than larger woodlands of a similar age, even if the larger woodlands harbor a higher biodiversity [Valdes 2020: 12].
Because of their high edge-to-core ratio, smaller woodlots get more sunlight and more nutrient input from surrounding farmland, resulting in denser vegetation cover and higher biomass production at edges.
This altered functioning in turn increases the delivery potential of some services, such as game production potential, due to an increased quantity of food available for game, and topsoil carbon storage, due to the faster incorporation of organic matter in the soil. Tick-borne disease risk is, however, lower, likely due to decreased larval densities in the unfavorable (e.g. hotter and drier) microclimatic conditions at the edge [Valdes 2020: 12].
While smaller woodlands were more apt to deliver “multiple services at higher levels of performance per area than larger woodlands of a similar age,” the greater biodiversity of larger woodlands increased certain individual ecosystem services.
The supply potential of several individual ecosystem services indirectly increased in larger and more ancient woodlands because it was dependent on higher levels of biodiversity. For example, abundance of usable plants and game production potential might have increased due to a positive correlation with vascular plant diversity, while pest control potential probably increased due to bottom-up effects through the trophic chain. On the contrary, tick-borne disease risk, topsoil carbon storage and stemwood volume were unrelated to multidiversity, probably because they depended on particular environmental conditions or on the presence and abundance of specific species rather than on species richness per se.
Finally, it should be noted that we focused on the service delivery potential on a per area basis and that the total amount of services provided by large patches might still be larger than that of small patches. Our findings should therefore not be interpreted as a trade-off between large, biodiverse patches versus small patches that have a higher potential to deliver services, but rather as an observation that small woodlands in agricultural landscapes have the potential to deliver a high flow of services relative to their size [Valdes 2020: 12].
Effectiveness of the Miyawaki method in Mediterranean forest restoration programs, Shirone, Salis & Vessela 2011
This study tested the Miyawaki method of rapid natural forest regeneration (which has been shown to work in Japan and elsewhere)in the arid Mediterranean. In this area, millennia of human civilization have resulted in degraded soils and reduced and changed forest cover, traditional reforestation efforts have often failed, and desertification is a looming threat. The Miyawaki method speeds up the process of ecological succession by densely planting a multilayer forest made up of a wide diversity of indigenous species.
In a natural forest cycle, as Clements (1916) described, annual plants on barren land are succeeded by perennial grass, sun-tolerant shrubs, light-demanding, fast-growing trees, and finally natural forests; each step may require decades, and the climax vegetation could be formed after two centuries or more. Currently, most forest reforestation programs adopt a scheme of planting one or more early successional species; after successful establishment, they are gradually replaced by intermediate species (either naturally or by planting), until late successional species arise. This pattern tries to simulate natural processes of ecological succession, from pioneer species to climax vegetation. However, it requires several silvicultural practices and normally takes a long time [Shirone 2011:82].
In the Miyawaki method, by contrast, one plants all at once the many plant species normally present in a native forest community, thus bypassing the earliest stages of ecological succession. Other tree-planting methods favor fast-growing non-native tree species, while omitting understory species – in other words, creating a simple plantation rather than a forest community that functions ecologically and can evolve and sustain itself.
Multilayer forests, and natural biocoenosis [ecological community] is possible, and well-developed ecosystems can be quickly established because of the simultaneous use of intermediate and late successional species in plantations. The Miyawaki method involves surveying the potential natural vegetation of the area to be reforested and recovering topsoil to a depth of 20– 30 cm by mixing the soil and a compost from organic materials, such as fallen leaves, mowed grass, etc. In this way, the time of the natural process of soil evolution, established by the vegetational succession itself, is reduced [Shirone 2011:82].
The authors of this study found that compared to traditional reforestation, there was “a more rapid development of trees on the Miyawaki plots, in particular, early-successional species [especially maritime pine]. The benefits over previous methods are remarkable and comparable with those obtained by Miyawaki in Asia and South America.” The Miyawaki method favoring denser plantings works even in arid climates, in spite of traditional views favoring sparse plantings in arid places, although the optimal density for the Mediterranean still needs testing, according to the authors.
In fact, low plant density has been traditionally retained as appropriate in arid and semiarid environments in order to avoid competition for water resources between plants, but it is now evident that cooperative processes, e.g., mutual shading, prevail over competitive processes. High plant density also reduces the impact of acorn predators, thus encouraging oak regeneration, i.e., the main late-successional forest species in Mediterranean environments. In addition, excellent plant stock remains fundamental for planting success in harsh environments [Shirone 2011:91].
When is a forest a forest? Forest concepts and definitions in the era of forest and landscape restoration, Chazdon et al. 2016
This article analyzes the policy context for forest ecosystem restoration, arguing that it is heavily shaped by the way we define a forest. The use of a forest definition lacking ecological considerations severely undermines conservation and restoration initiatives.
We live in an era of unprecedented environmental change, motivating equally unprecedented global actions to protect and restore forest ecosystems. These efforts could fail to achieve their ambitious goals if they are not informed by clear and appropriate concepts and definitions of forests [Chazdon 2016: 1].
There are multiple definitions of a forest. Early European and internationally adopted definitions tended to define forests according to their usage for timber. FAO’s 1948 definition created for assessing wood harvesting potential of the world’s forests is still in use today. Yet new definitions have since been created that emphasize conservation, carbon sequestration and biodiversity values of forests.
However, national and global forest assessments tend to use narrow technical definitions that ignore ecological values of forested land.
In many cases, forest assessments do not distinguish between land covered by natural and planted forests. Thus, if natural forests are cleared and replaced with plantations, no net loss of forest cover is reported [Chazdon 2016: 6].
In other words, areas that should not be considered forest in ecological terms are counted as forest – an obfuscation with disastrous environmental outcomes. Similarly, ecologically important yet small patches of trees that are not counted in forest inventories and lack legal protection are at risk of being lost.
Areas classified as ‘‘non-forests’’ are as important to forest definitions as are forests. More than 43 % of agricultural land globally is in agroforestry systems with 10 % tree cover. In Rwanda and Brazil, forest inventories using a 0.5-ha threshold ignore substantial areas of small forest fragments, agroforests, and woodlots, leading to underestimates of actual tree cover. Small patches of trees and even isolated remnant trees can hold high ecological and conservation value, and can play an important role in enhancing landscape connectivity, local biodiversity, and local livelihoods [Chazdon 2016: 7].
Information from participatory local monitoring and remote sensing technology that distinguishes “among successional stages of forests, selectively logged forests, and single-species plantations” [Chazdon 2016: 10] is needed.
Access to this information will allow countries and international agencies to track changes in natural forest cover, and to monitor processes of restoration, rehabilitation, and afforestation within a landscape context and, consequently, make informed policy decisions. We are on the frontier of developing new ways of monitoring and assessing land cover that will provide robust indicators of the quality and origins of tree cover and enable new ways of viewing and defining forests and reforests. To see beyond the overly simplified categories of forest loss, forest degradation, and forest gain, we need to develop and apply more adapted and nuanced definitions that will deepen our understanding of the drivers and outcomes of land-use change and forest dynamics within landscapes [Chazdon 2016: 10].
The UN’s Decade of Ecosystem Restoration declaration aims to “prevent, halt and reverse the degradation of ecosystems worldwide,” stating that “there has never been a more urgent need to restore damaged ecosystems than now” [UNEP/FAO Factsheet 2020].
Estimates of global land degradation range from 25% to 75% of Earth’s land surface. The uncertainty is due to different ideas about what counts as degraded land and different methodologies for quantifying it (expert opinion, satellite data, or modeling, for example). Generally, degradation is defined as “a reduction in productivity of the land or soil due to human activity” [Gibbs & Salmon 2015: 13].
Another global assessment [IPBES 2019] states that 75% of the land surface “is significantly altered, 66 percent of the ocean area is experiencing increasing cumulative impacts, and over 85 percent of wetlands (area) has been lost” [IPBES 2019: 11]. Meanwhile, wilderness remains on just 23% of Earth’s land surface [Watson 2016]. Wilderness areas are defined as
biologically and ecologically largely intact landscapes that are mostly free of human disturbance. These areas do not exclude people, as many are in fact critical to certain communities, including indigenous peoples. Rather, they have lower levels of impacts from the kinds of human uses that result in significant biophysical disturbance to natural habitats, such as large-scale land conversion, industrial activity, or infrastructure development [Watson 2016: 1].
Degraded lands have reduced ecosystem function, upon which humans and other beings depend for clean water and air, shelter, food, and habitable local and global climate systems. In fact, half of Earth’s surface (including ocean and land) should be maintained or restored to intact ecosystems, in addition to cutting fossil fuel emissions, for any chance of keeping global warming from surpassing 1.5C above pre-industrial levels and averting catastrophic climate change [Dinerstein 2019].
This means existing wilderness needs to be protected and degraded lands regenerated. Currently less than 15% of land surface is formally protected and 2% of oceans [Dinerstein 2019]. Furthermore, intact ecosystems need to be connected by wildlife corridors to allow for migration and dispersal of diverse species. Quantifying a few of the direct human benefits from large-scale restoration, the UN estimates that:
Restoration of 350 million hectares of degraded land between now and 2030 could generate USD 9 trillion in ecosystem services and take an additional 13-26 gigatons of greenhouse gases out of the atmosphere [UNEP 2019].
So how does ecosystem restoration happen? There are critical social, political and cultural responses to this question that are beyond the scope of this review – except to stress that all hands are needed on deck. As the UN says:
This incredible challenge can only be met if everyone – including member states, local governments, partners from the private sector, academia and civil society – come together to find viable, lasting solutions [UNEP/FAO Factsheet 2020: 1].
Here we present a handful of the solutions as discussed in a growing body of ecological restoration literature, such as the articles summarized in the following section. There are two overarching approaches – active versus passive restoration. The latter aims simply to remove the anthropogenic disturbance causing the degradation and allow abandoned or no-longer-disturbed land to regenerate on its own. Sometimes simple removal of the disturbance (such as a dam) involves plenty of human energy, and may not seem passive at all. By contrast, though, active restoration actively facilitates land regeneration. Activities may include installing hollow logs, wood piles or other habitat features; planting; dredging; prescribed burning; reintroducing key species; or controlling invasive species, for example.
A somewhat intermediate approach of planting “tree islands” to restore tropical forests is another option [Holl 2020]. Instead of planting rows of trees throughout a given plot, clusters of trees are planted on just a fraction of the area, costing just a fraction of the price of a plantation-style restoration effort. Tree islands simulate the patchiness of natural forest recovery, while speeding up the process, and rely on animals to disperse tree seeds. In a study in Costa Rica of the tree island restoration method, cover of trees and shrubs had increased from 20% to over 90% over 15 years [Holl 2020].
Deciding which approach is best may depend on the situation – the degree of degradation needing to be reversed or the proximity of adjacent ecosystems as a seed source to kickstart colonization. Funding may also be a factor as active restoration projects typically cost more than do passive ones. Interestingly, in spite of being less expensive and often less work, passive regeneration projects are comparable to or more effective than active restoration in terms of achieving outcomes, according to two meta-studies [Jones 2018, Crouzeilles 2017]. Desired outcomes for restoration projects relate to measures of biodiversity, density and height of vegetation, amount of biomass produced, and speed of recovery, for example.
Letting ecosystems repair themselves in many cases may be the most effective restoration strategy – a counterintuitive yet critical finding that could help society allocate restoration funds more efficiently in the future [Jones 2018: 6].
The most effective strategy for a given situation is not always taken, and projects sometimes fail or only partially succeed. One reason for underperformance is simply that ecosystems are complex and restoring them requires “significant time, resources, and knowledge,” which may not always be available [Gann 2019: 14]. Necessary follow up after activities are completed may never happen, such as when trees are planted and then abandoned in the first few critical years of establishment. Another potential roadblock to ecological recovery is competing goals for a project, which may be skewed toward economic concerns.
For instance, the Bonn Challenge to restore 330 million hectares of deforested and degraded land by 2030 has been criticized for counting monoculture timber plantations in progress toward reforestation goals [Lewis & Wheeler 2019]. Such woodlots offer little in the way of wildlife habitat and store an estimated 40 times less carbon than do natural forests [Lewis & Wheeler 2019]. Part of the problem of allowing such practices to count as restoration lies in the definition of “forest” used in setting goals and making policy; definitions may lack consideration of biodiversity or other essential elements of natural forests. The UN Food and Agriculture Organization’s definition in use today was originally created to facilitate timber inventories [Chazdon 2016].
Early afforestation efforts in arid and semi-arid northern China reveals how restoration can go wrong when natural ecosystem characteristics are ignored [Cao 2008]. Much of the area, whose natural state is grasslands, had become desertified by the middle of the 20th Century due to agriculture, overgrazing and monoculture timber plantations. Large-scale afforestation began in 1978, when modern restoration science was not yet well established. Plantings involved fast-growing water-use inefficient tree species not well adapted to arid environments, which only exacerbated dry conditions, rather than native grassland shrubs.
The natural vegetation of much of the region was desert steppe vegetation or dryland shrub communities, which have a much higher water-use efficiency than most tree communities and which have evolved to use soil water sustainably under these environmental conditions [Cao 2008: 1828].
In his analysis of the project, Shixiong Cao [2008] recommends a change of practice.
In terms of revegetation strategies, planners must understand that different environments will support different vegetation communities and that forests are not a suitable choice in all areas. To successfully revegetate an area, planners must determine which vegetation types a given environment can naturally sustain and target restoration activities at creating such communities. For example, stable communities of natural desert steppe and grassland vegetation, and possibly even lichen species in more severely degraded environments, can develop in arid and semiarid areas as a result of natural processes, thereby increasing vegetation cover beyond the levels that could be sustained for trees, and can thereby provide better protection for the soil [Cao 2008: 1830].
The concept of rewilding, which entered ecological restoration discourse a couple of decades ago, has sparked confusion and controversy due to its multiple definitions [Hayward 2019]. However, the concept has also perhaps served to reinforce ecological principles within the larger restoration movement. Rewilding is an approach to restoring ecosystems that “aims to restore self-sustaining and complex ecosystems with interlinked ecological processes that promote and support one another while minimizing or gradually reducing human interventions” [Perino 2019: 1].
For example, rewilding has often focused on the roles of top predators in ecosystem processes, and proposed their reintroduction as a key tactic. The addition of large animals to a system enhances trophic complexity and dispersal. Along with natural disturbances (like natural fires), these are key ecosystem processes rewilding aims to activate.
Rewilding aims to restore these three ecological processes [trophic complexity, dispersal and natural disturbances] to foster complex and self-organizing ecosystems that require minimum human management in the long term [Perino 2019: 2].
Establishing ecological corridors that connect larger intact ecosystems can facilitate migration and dispersal of plants and animals to colonize new areas. The value of ecological corridors highlights the role of small-scale, local restoration and conservation projects in rebuilding landscape-level ecological integrity. Protecting and expanding hedges, river systems and roadsides contributes to the success of the larger wilderness areas they connect. Similarly, even small woodlots in agricultural landscapes can have unexpectedly high ecosystem functionality [Valdes 2019]. The relevance of small projects to overall ecological wellbeing means that almost anybody anywhere has a role to play.
Lastly, the importance of conserving existing ecosystems cannot be overstated, and restoration projects should never serve to justify destruction elsewhere – mature intact ecosystems are irreplaceable. Rather, ecosystem restoration should be viewed as a strategy that “seeks to advance conservation by rebuilding nature” [Young & Schwartz 2019: 1]. Many restoration projects achieve only partial recovery during relevant time periods, meaning they may not ever become as fully functional as mature intact ecosystems during our lifetimes [Jones 2018]. Existing mature forests and other ecosystems are major carbon sinks in terms of the large amount of carbon they contain and in terms of the superior sequestration rates of older larger trees [Moomaw 2019]. Intact forests are also home to about two thirds of all species on Earth [Dinerstein 2019].
Both conservation and restoration are essential – they are an inseparable two-part solution to a dire global ecological crisis.
Lastly, the importance of conserving existing ecosystems cannot be overstated, and restoration projects should never serve to justify destruction elsewhere – mature intact ecosystems are irreplaceable. Rather, ecosystem restoration should be viewed as a strategy that “seeks to advance conservation by rebuilding nature” [Young & Schwartz 2019: 1].
Approaches to ecosystem restoration article summaries
Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests, Crouzeilles et al. 2017
This meta-analysis comparing active restoration to natural ecosystem regeneration found the latter to be more effective. The authors conclude that “lower-cost natural regeneration surpasses active restoration in achieving tropical forest restoration success for biodiversity and vegetation structure[7]” [Crouzeilles 2017: 4]. This conclusion runs counter to conventional wisdom that active restoration is preferable despite being more expensive.
Natural forest regeneration is the spontaneous recovery of native tree species that colonize and establish in abandoned fields or natural disturbances; this process can also be assisted through human interventions such as fencing to control livestock grazing, weed control, and fire protection. In contrast, active restoration requires planting of nursery-grown seedlings, direct seeding, and/or the manipulation of disturbance regimes (for example, thinning and burning) to speed up the recovery process, often at a high cost to establish structural features of the vegetation (hereafter termed vegetation structure), reassemble local species composition, and/or catalyze ecological succession [Crouzeilles 2017: 1].
However, “restoration success for biodiversity and vegetation structure was significantly lower in both natural regeneration and active restoration than in reference systems” [Crouzeilles 2017: 2], underscoring the importance of conserving existing intact ecosystems.
“Restoration success for biodiversity and vegetation structure was significantly lower in both natural regeneration and active restoration than in reference systems” [Crouzeilles 2017: 2], underscoring the importance of conserving existing intact ecosystems.
Part of the explanation for the lower success of active restoration compared to natural regeneration is that the composition and/or diversity of species chosen for planting in active restoration may be inappropriate, while the species that colonize abandoned land are likely to be diverse and locally adapted.
Natural regeneration is initiated through the colonization of opportunistic and locally adapted species, resulting in a stochastic dynamic process of forest restoration that ultimately leads to higher diversity of native, locally adapted plant species than in tree planting schemes (that is, active restoration). Active restoration also can create a highly diverse habitat through human introduction of up to 6000 seedlings/ha, but tree species used in plantings often lack the full range of functional traits found in natural regrowth forests. In addition, most tropical forest plantings for restoration or forest plantations use relatively few species, that is, these plantations may not be planted primarily for biodiversity outcomes. Thus, the higher plant biodiversity in naturally regenerated systems creates more habitats and resources, which provide additional sources of food, shelter, nesting, and breeding sites, to support higher animal biodiversity [Crouzeilles 2017: 2].
Restoration and repair of Earth’s damaged ecosystems, Jones et al. 2018
This meta-analysis of 400 studies compared passive and active ecosystem repair outcomes in terms of the speed and completeness of recovery, and found little difference between the two approaches.
Active restoration did not result in faster or more complete recovery than simply ending the disturbances ecosystems face [Jones 2018: 1].
Passive recovery simply means ending the anthropogenic disturbance that was causing the degradation, while active restoration here includes anything from fertilizer application to recontouring/dredging to planting a desired species mix.
The authors speculate that the lack of different outcomes between the two approaches could be due to restoration managers correctly choosing to actively restore the ecosystems that “are not recovering on their own and require active restoration to improve recovery outcomes relative to passively recovering systems” [Jones 2018: 6]. Also, the actively managed sites in the study had, on average, less time to recover than the passively managed sites. Finally, the authors suggest there may be right and wrong ways to actively restore ecosystems, and “recommend that restoration strategies be tailored more closely to overcome the specific barriers to recovery in individual sites” [Jones 2018: 6].
Assuming active and passive restoration achieve comparable outcomes in many cases, then passive restoration deserves serious consideration, given limited resources available for the vast amount of ecosystem repair required in the world today.
Letting ecosystems repair themselves in many cases may be the most effective restoration strategy – a counterintuitive yet critical finding that could help society allocate restoration funds more efficiently in the future [Jones 2018: 6].
The study also consistently found that across systems, ecosystems didn’t fully recover, at least not within the timeframe of the studies.
Our results expand those findings to a broader range of ecosystems and geographies, and, together with previous work, suggest the majority of ecosystems have not yet recovered fully following disturbance and may not in the future. Thus, restoration should not be considered a substitute for conservation, which is a key strategy to ensure sustained support of biodiversity and delivery of ecosystem services in the future [Jones 2018: 4].
Rewilding complex ecosystems, Perino et al. 2019
A growing body of literature emphasizes the need for novel, process-oriented approaches to restoring ecosystems in our rapidly changing world. Dynamic and process-oriented approaches focus on the adaptive capacity of ecosystems and the restoration of ecosystem processes promoting biodiversity, rather than aiming to maintain or restore particular ecosystem states characterized by predefined species compositions or particular bundles of ecosystem services [Perino 2019: 1].
In contrast to other types of restoration efforts aiming to recreate the composition and appearance of an historical ecological community, rewilding focuses on ecosystem function and recognizes the dynamic and unpredictable nature of ecosystems. This article highlights three key ecological processes that rewilding aims to activate: trophic complexity, natural disturbances and dispersal.
Rewilding aims to restore these three ecological processes to foster complex and self-organizing ecosystems that require minimum human management in the long term [Perino 2019: 2].
Trophic complexity implies the presence of large vertebrates, including herbivores that modify the landscape through grazing or dam building and predators that control the herbivore populations. These keystone species can promote biodiversity in the landscapes they inhabit.
Stochastic (random) natural disturbance (such as fire or flooding) can increase ecosystem heterogeneity and complexity, allowing less competitive species to survive. Rewilding involves discontinuing both controlled anthropogenic disturbances and suppression of natural disturbances.
Rewilding actions aim to release ecosystems from continued and controlled anthropogenic disturbances to allow for natural variability and sources of stochasticity. Mowing of grassland can be reduced or replaced by natural grazing. Dams can be removed or their management modified to restore natural flood regimes. Logging can be replaced by allowing natural fire and pest regimes [Perino 2019: 4].
Dispersal– rewilding aims to remove anthropogenic barriers that limit the movement of plants and animals and thus the dispersal of their genetic material and potential for recolonization after a disturbance event. The creation of ecological corridors is an example of a rewilding activity that enhances dispersal.
The interaction of these ecological processes boosts the functioning of each. For example, the presence of larger animals facilitates seed dispersal throughout the system. High levels of dispersal, in turn, can facilitate ecosystem recovery following a disturbance.
Rewilding projects can be passive (allowing abandoned agricultural fields to recover on their own) or active (species reintroductions, for example), and are most effective when conducted in a manner that engages the local community in the process.
Rewilding: a call for boosting ecological complexity in conservation, Fernández et al. 2017
Rewilding is gaining traction as an approach to conservation. However, many different perspectives about which species and ecological processes to focus rewilding efforts on and how deeply to intervene in systems has created some confusion and contention within the field. Furthermore, the most ambitious and extreme rewilding proposals (for example, recreating communities that went extinct millennia ago) have often attracted more attention, while the more pragmatic and immediate solutions in the field are overlooked.
This article attempts to clarify the concept. The authors emphasize that rewilding is a process-oriented approach to biodiversity conservation “focused on preserving and restoring the structural and functional complexity of degraded ecosystems” [Fernandez 2017: 276].
Rewilding pursues the goal of restoring wild species interactions and their regulation of key ecosystem processes including nutrient and energy flows, vegetation succession and disturbances, drawing specific attention to the key roles of large-bodied species that are especially sensitive to the human appropriation of landscapes [Fernandez 2017: 277].
The authors suggest further research is needed. They note that while the negative effects on ecosystems of the loss of biodiversity and keystone species is well documented, ecosystem responses to species reintroduction and other rewilding efforts are not as well studied. To guide future research, the authors
propose an unequivocally process-oriented formulation of the “rewilding hypothesis” as a general guidance: that the large-scale restoration of apex consumers and large herbivores promotes self-regulation in community assemblages, and increases the complexity of ecological processes in ecosystems [Fernandez 2016: 277].
Also needed is policy and management practice support, particularly in terms of protecting the areas and species in question. Proactive policies could ensure that gains made toward ecological restoration are not undermined by damaging human activity.
Policies and practices should be developed in order to enforce the idea that rewilding is about reducing the human control on ecosystem processes. It must begin to include varied objectives to alleviate pressures on wildlife populations such as a full legal protection of large predators based on their unique ecological roles and not just depending on their conservation status; the eradication of predator control programs; or the elimination of game management practices such as wildlife fencing, introduction of alien game populations, supplementary feeding and others that profoundly alter the natural regulation and the genetic structure of large herbivore populations [Fernandez 2016: 278].
The differences between rewilding and restoring an ecologically degraded landscape, du Toit & Pettorelli 2019
This commentary distinguishes between restoration and rewilding of ecosystems, explaining that the latter aims at ecological adaptation to novel local environmental conditions wrought by global climate change. By contrast, restoration, as defined here, aims to recreate and maintain an historical state or condition of an ecosystem, regardless of current environmental conditions.
Although the two words are often conflated,
Rewilding is thus conceptually different from restoring. It is an adaptive approach to conserving ecological functionality under changing environmental conditions, to which historical benchmarks are less relevant than to restoring. It inherently acknowledges and promotes unpredictability, while placing the emphasis on function over species composition [du Toit & Pettorelli 2019: 2].
The authors assert that rewilding is better suited to preserving biodiversity and ecosystem function under present and future conditions.
It is difficult to imagine how conserving biodiversity and ecosystem services could be possible in predicted future scenarios without rewilding. Simply stated, anthropogenic environmental forcing makes ecosystem restoration a diminishing option [du Toit & Pettorelli 2019: 4].
Reintroducing rewilding to restoration – rejecting the search for novelty, Hayward et al. 2019
This perspective piece argues against scientific or public adoption of the term “rewilding,” which the authors view as being generally synonymous with the classical and better-understood concept of ecological restoration. Definitions of restoration are sufficient to encompass practices espoused in rewilding.
Early definitions of restoration describe the practice as “the process of repairing damage caused by humans to the diversity and dynamics of indigenous ecosystems” (Jackson et al., 1995). Although at the time of this definition, restoration science was still developing, it was clear that it had established itself under the broad banner of repairing damaged ecosystems. … More recently, restoration has been defined as “any activity whose aim it is to ultimately achieve ecosystem recovery, insofar as possible and relative to an appropriate local native model (termed here a reference ecosystem), regardless of the period of time required to achieve the recovery outcome” (McDonald et al., 2016) [Hayward 2019: 257].
The term rewilding, which has evolved over time, “was arguably conceived to promote the original authors’ view of conservation via cores [habitats], corridors, and carnivores” [Hayward 2019: 256]. In this early context,
‘rewilding’ referred to conservation and management interventions that focused on reintroducing keystone predators and ensuring that they had sufficient interconnected space to live. The authors emphasized within their original work that rewilding was “one essential element in most efforts to restore fully functioning ecosystems” (Soulé and Noss, 1998). As such, it is clear that rewilding was originally aimed to be a term that referred to one component of ecological restoration [Hayward 2019: 256].
Since then, rewilding has come to refer to practices involving “translocating substitute species to fill vacant ecological niches left by extinct species” [Hayward 2019: 257] or reintroducing locally extinct species, or simply allowing natural succession to occur on abandoned land. Given multiple definitions, all of which relate to the idea of restoration, the term rewilding is seen here as superfluous and confusing.
Given the lack of clear differences between rewilding and restoration in both definition and practice, we see little need for these competing terms within scientific discourse [Hayward 2019: 257].
However, the authors suggest two positive contributions from the rewilding discourse. Because of its overall focus on large fauna, rewilding has captured public imagination and interest in conservation, while also helping to shift a potential vegetation bias among restoration practitioners/scientists toward equal emphasis on the ecosystem role of animals.
Therefore, rather than adopting a new term with copious definitions that lack clarity, this debate can be used as an opportunity to adaptively improve current restoration practice by incorporating a more equal focus between flora and fauna [Hayward 2019: 258].
Because of its overall focus on large fauna, rewilding has captured public imagination and interest in conservation, while also helping to shift a potential vegetation bias among restoration practitioners/scientists toward equal emphasis on the ecosystem role of animals.
Intact forests in the United States: proforestation mitigates climate change and serves the greatest good, Moomaw 2019
The concept of “proforestation” presented here means letting existing forests continue to grow and reach their full ecological potential. Due to intensive management practices, most existing forests sequester carbon at only half (or less) of their potential rate. In addition to storing (embodying) more carbon than their smaller counterparts, large trees also sequester carbon at a faster rate. For example, “Each year a single tree that is 100 cm in diameter adds the equivalent biomass of an entire 10-20 cm diameter tree, further underscoring the role of large trees” [Moomaw 2019: 4]. Imagine, reader, that every year you planted a whole new medium-sized tree – that’s essentially what large trees are doing.
“Each year a single tree that is 100 cm in diameter adds the equivalent biomass of an entire 10-20 cm diameter tree, further underscoring the role of large trees” [Moomaw 2019: 4].
Much of Maine’s forests have been harvested continuously for 200 years and have a carbon density less than one-third of the forests of Southern Vermont and New Hampshire, Northwestern Connecticut and Western Massachusetts – a region that has not been significantly harvested over the past 75-150 years. …
Ecosystem services accrue as forests age for centuries. Far from plateauing in terms of carbon sequestration (or added wood) at a relatively young age as was long believed, older forests (e.g., >200 years of age without intervention) contain a variety of habitats, typically continue to sequester additional carbon for many decades or even centuries, and sequester significantly more carbon than younger and managed stands [Moomaw 2019: 5].
Because existing forests are already sequestering carbon, and will continue at an increasing rate as tree size grows, the author argues that proforestation is a more effective immediate solution than either reforestation (planting new trees where they had been cleared for agriculture, etc.) or afforestation (planting trees in new places), though these other two approaches are important for longer term ecosystem function. Moomaw et al. argue that the urgency of removing CO2makes it imperative to keep existing forests growing.
Globally, existing forests only store approximately half of their potential due to past and present management, and many existing forests are capable of immediate and even more extensive growth for many decades. During the timeframe while seedlings planted for afforestation and reforestation are growing (yet will never achieve the carbon density of an intact forest), proforestation is a safe, highly effective, immediate natural solution that does not rely on uncertain discounted future benefits inherent in other options [Moomaw 2019: 7].
Furthermore, existing, older forests are critical habitats for threatened wildlife, even small intact woods.
Forest bird guilds also benefit from small intact forests in urban landscapes relative to unprotected matrix forests. Several bird species in the U.S. that are globally threatened – including the wood thrush, cerulean warbler, marbled murrelet, and spotted owl are, in part, dependent on intact, older forests with large trees [Moomaw 2019: 5].
In sum, proforestation provides the most effective solution to dual global crises – climate change and biodiversity loss. It is the only practical, rapid, economical, and effective means for atmospheric CDR [carbon dioxide removal] among the multiple options that have been proposed because it removes more atmospheric carbon dioxide in the immediate future and continues to sequester it long-term. Proforestation will increase the diversity of many groups of organisms and provide numerous additional and important ecosystem services. While multiple strategies will be needed to address global environmental crises, proforestation is a very low-cost option for increasing carbon sequestration that does not require additional land beyond what is already forested and provides new forest related jobs and opportunities along with a wide array of quantifiable ecosystem services, including human health [Moomaw 2019: 8].
Plant diversity enhances the reclamation of degraded lands by stimulating plant-soil feedbacks, Jia et al. 2020
This study tested biodiversity effects on ecosystem function in the process of reviving severely degraded and contaminated land, and found that “increasing plant diversity greatly enhanced the reclamation of these lands” [Jia 2020: 1].
Prior to implementing the reclamation experiment, the degraded mine wasteland investigated in this study was heavily impacted by past mining activities and was devoid of vegetation for more than a decade and the soil lacked structure, contained high levels of toxic metals and low levels of nutrients. … our results showed that higher plant species richness enhanced land reclamation across all standard measures of reclamation success and specifically resulted in higher vegetation coverage, biomass yield and stability for all 3 years [of the experiment] [Jia 2020: 6].
Furthermore, higher biodiversity plots had higher levels of organic carbon in the soil, higher soil microorganism abundance, lower fungal pathogens, and lower heavy metal concentrations in plant tissue.
The most striking impact of plant diversity on soil was on the microbial communities. Both soil fungal and bacterial OTUs [operational taxonomic units[8]] increased significantly with plant species richness. More importantly, we found that higher plant species richness significantly increased the relative abundance of soil cellulolytic bacteria that degrade cellulose and are thus essential components of nutrient cycling [Jia 2020: 7].
High ecosystem service delivery potential of small woodlands in agricultural landscapes, Valdes 2020
This article assesses the ecological value of small woodlands relative to larger ones. The authors conclude that:
…smaller woodlands potentially deliver multiple services at higher performance levels on a per area basis than larger woodlands of a similar age, even if the larger woodlands harbor a higher biodiversity [Valdes 2020: 12].
Because of their high edge-to-core ratio, smaller woodlots get more sunlight and more nutrient input from surrounding farmland, resulting in denser vegetation cover and higher biomass production at edges.
This altered functioning in turn increases the delivery potential of some services, such as game production potential, due to an increased quantity of food available for game, and topsoil carbon storage, due to the faster incorporation of organic matter in the soil. Tick-borne disease risk is, however, lower, likely due to decreased larval densities in the unfavorable (e.g. hotter and drier) microclimatic conditions at the edge [Valdes 2020: 12].
While smaller woodlands were more apt to deliver “multiple services at higher levels of performance per area than larger woodlands of a similar age,” the greater biodiversity of larger woodlands increased certain individual ecosystem services.
The supply potential of several individual ecosystem services indirectly increased in larger and more ancient woodlands because it was dependent on higher levels of biodiversity. For example, abundance of usable plants and game production potential might have increased due to a positive correlation with vascular plant diversity, while pest control potential probably increased due to bottom-up effects through the trophic chain. On the contrary, tick-borne disease risk, topsoil carbon storage and stemwood volume were unrelated to multidiversity, probably because they depended on particular environmental conditions or on the presence and abundance of specific species rather than on species richness per se.
Finally, it should be noted that we focused on the service delivery potential on a per area basis and that the total amount of services provided by large patches might still be larger than that of small patches. Our findings should therefore not be interpreted as a trade-off between large, biodiverse patches versus small patches that have a higher potential to deliver services, but rather as an observation that small woodlands in agricultural landscapes have the potential to deliver a high flow of services relative to their size [Valdes 2020: 12].
Effectiveness of the Miyawaki method in Mediterranean forest restoration programs, Shirone, Salis & Vessela 2011
This study tested the Miyawaki method of rapid natural forest regeneration (which has been shown to work in Japan and elsewhere)in the arid Mediterranean. In this area, millennia of human civilization have resulted in degraded soils and reduced and changed forest cover, traditional reforestation efforts have often failed, and desertification is a looming threat. The Miyawaki method speeds up the process of ecological succession by densely planting a multilayer forest made up of a wide diversity of indigenous species.
In a natural forest cycle, as Clements (1916) described, annual plants on barren land are succeeded by perennial grass, sun-tolerant shrubs, light-demanding, fast-growing trees, and finally natural forests; each step may require decades, and the climax vegetation could be formed after two centuries or more. Currently, most forest reforestation programs adopt a scheme of planting one or more early successional species; after successful establishment, they are gradually replaced by intermediate species (either naturally or by planting), until late successional species arise. This pattern tries to simulate natural processes of ecological succession, from pioneer species to climax vegetation. However, it requires several silvicultural practices and normally takes a long time [Shirone 2011:82].
In the Miyawaki method, by contrast, one plants all at once the many plant species normally present in a native forest community, thus bypassing the earliest stages of ecological succession. Other tree-planting methods favor fast-growing non-native tree species, while omitting understory species – in other words, creating a simple plantation rather than a forest community that functions ecologically and can evolve and sustain itself.
Multilayer forests, and natural biocoenosis [ecological community] is possible, and well-developed ecosystems can be quickly established because of the simultaneous use of intermediate and late successional species in plantations. The Miyawaki method involves surveying the potential natural vegetation of the area to be reforested and recovering topsoil to a depth of 20– 30 cm by mixing the soil and a compost from organic materials, such as fallen leaves, mowed grass, etc. In this way, the time of the natural process of soil evolution, established by the vegetational succession itself, is reduced [Shirone 2011:82].
The authors of this study found that compared to traditional reforestation, there was “a more rapid development of trees on the Miyawaki plots, in particular, early-successional species [especially maritime pine]. The benefits over previous methods are remarkable and comparable with those obtained by Miyawaki in Asia and South America.” The Miyawaki method favoring denser plantings works even in arid climates, in spite of traditional views favoring sparse plantings in arid places, although the optimal density for the Mediterranean still needs testing, according to the authors.
In fact, low plant density has been traditionally retained as appropriate in arid and semiarid environments in order to avoid competition for water resources between plants, but it is now evident that cooperative processes, e.g., mutual shading, prevail over competitive processes. High plant density also reduces the impact of acorn predators, thus encouraging oak regeneration, i.e., the main late-successional forest species in Mediterranean environments. In addition, excellent plant stock remains fundamental for planting success in harsh environments [Shirone 2011:91].
When is a forest a forest? Forest concepts and definitions in the era of forest and landscape restoration, Chazdon et al. 2016
This article analyzes the policy context for forest ecosystem restoration, arguing that it is heavily shaped by the way we define a forest. The use of a forest definition lacking ecological considerations severely undermines conservation and restoration initiatives.
We live in an era of unprecedented environmental change, motivating equally unprecedented global actions to protect and restore forest ecosystems. These efforts could fail to achieve their ambitious goals if they are not informed by clear and appropriate concepts and definitions of forests [Chazdon 2016: 1].
There are multiple definitions of a forest. Early European and internationally adopted definitions tended to define forests according to their usage for timber. FAO’s 1948 definition created for assessing wood harvesting potential of the world’s forests is still in use today. Yet new definitions have since been created that emphasize conservation, carbon sequestration and biodiversity values of forests.
However, national and global forest assessments tend to use narrow technical definitions that ignore ecological values of forested land.
In many cases, forest assessments do not distinguish between land covered by natural and planted forests. Thus, if natural forests are cleared and replaced with plantations, no net loss of forest cover is reported [Chazdon 2016: 6].
In other words, areas that should not be considered forest in ecological terms are counted as forest – an obfuscation with disastrous environmental outcomes. Similarly, ecologically important yet small patches of trees that are not counted in forest inventories and lack legal protection are at risk of being lost.
Areas classified as ‘‘non-forests’’ are as important to forest definitions as are forests. More than 43 % of agricultural land globally is in agroforestry systems with 10 % tree cover. In Rwanda and Brazil, forest inventories using a 0.5-ha threshold ignore substantial areas of small forest fragments, agroforests, and woodlots, leading to underestimates of actual tree cover. Small patches of trees and even isolated remnant trees can hold high ecological and conservation value, and can play an important role in enhancing landscape connectivity, local biodiversity, and local livelihoods [Chazdon 2016: 7].
Information from participatory local monitoring and remote sensing technology that distinguishes “among successional stages of forests, selectively logged forests, and single-species plantations” [Chazdon 2016: 10] is needed.
Access to this information will allow countries and international agencies to track changes in natural forest cover, and to monitor processes of restoration, rehabilitation, and afforestation within a landscape context and, consequently, make informed policy decisions. We are on the frontier of developing new ways of monitoring and assessing land cover that will provide robust indicators of the quality and origins of tree cover and enable new ways of viewing and defining forests and reforests. To see beyond the overly simplified categories of forest loss, forest degradation, and forest gain, we need to develop and apply more adapted and nuanced definitions that will deepen our understanding of the drivers and outcomes of land-use change and forest dynamics within landscapes [Chazdon 2016: 10].
Increasing agricultural production to feed >11 billion people by 2100 raises several challenges for effectively managing infectious disease. Of many factors examined in this article linking agricultural expansion to infectious disease, one is conversion of natural habitat to cropland or rangeland. Land conversion increases contact between wild animals, livestock and humans.
As natural ecosystems are converted to crop land or range land, interactions among humans, and domesticated and wild animals, could increase. … These interactions are crucial because 77% of livestock pathogens are capable of infecting multiple host species, including wildlife and humans, and based on published estimates from the 2000s, over half of all recognized human pathogens are currently or originally zoonotic, as are 60–76% of recent emerging infectious disease events [Rohr 2019: 451].
“As natural ecosystems are converted to crop land or range land, interactions among humans, and domesticated and wild animals, could increase” [Rohr 2019: 451].
Land conversion pushes humans and livestock up against wilderness areas, increasing contact between species with previously little to no contact. The jumping of a pathogen to a new host species is called “spillover.”
Spillover appears to be a function of the frequency, duration and intimacy of interactions between a reservoir and novel host population. For example, influenza is believed to have jumped from horses to humans soon after domesticating horses and then made additional jumps to humans from other domesticated animals, such as poultry and swine [Rohr 2019: 451].
Furthermore, agricultural intensification tends to involve greater concentrations of a single variety of a single species, increasing the risk that any new disease will spread quickly in the population.
A central tenet of epidemiology is that the incidence of many infectious diseases should increase proportionally with host density because of increased contact rates and thus transmission among hosts. Hence, increasing human and livestock densities could cause increases in infectious diseases unless investments in disease prevention are sufficient to prevent these increases [Rohr 2019: 451].
Industrial-scale confined livestock production is
vulnerable to devastating losses of animals to disease. For instance, in just the last 25 years, an influenza A virus (H5N1) and a foot-and-mouth outbreak led to the destruction of more than 1.2 million chickens and 6 million livestock in China and Great Britain, respectively, and a ‘mad cow disease’ epizootic led to the slaughter of 11 million cattle worldwide [Rohr 2019: 449].
Increased agricultural production tends to be accompanied by new irrigation infrastructure and increased pesticide, fertilizer and antibiotic use, all of which increase infectious disease risk. Dams (often created for irrigation schemes) increase risk of mosquito-borne disease. Antibiotic overuse for livestock fosters resistance among pathogens that can also infect humans. Greater pesticide use leads to resistance among disease vectors such as mosquitoes to insecticides, while also weakening immune systems among exposed humans and wildlife hosts, increasing infection rates/severity. Nutrient enrichment caused by fertilizer can also contribute to the spread of infectious disease, for example, through mosquitos or snail vectors.
Finally, the urbanization and globalization associated with agricultural intensification/expansion elongates food supply chains, which increases movement of people and goods over borders, spreading food-born illness, flu and other infections.
In short,
These analyses revealed that agricultural drivers were associated with 25% of all diseases and nearly 50% of zoonotic diseases that emerged in humans since 1940. These values are even higher if we include the use of antimicrobial agents as an agricultural driver of human disease emergence, given that agricultural uses of antibiotics outpace medical uses in the developed world nearly nine to one [Rohr 2019: 451].
The authors recommend numerous measures for improving agricultural production while limiting infectious disease, including reducing antibiotic use for livestock, conserving biodiversity, improving and diversifying livestock and crop genetic material, investing in urban agriculture, social investments, and inter-disciplinary research and collaboration.
This article introduces the concept of One Health, a public health framework adopted by the Centers for Disease Control in 2009, which recognizes the interdependence of humans, animals and our shared environment. The concept has gained traction as a way to address health problems arising from global environmental change.
Climate change, loss of biodiversity, habitat fragmentation and pollution, and subsequent degradation of natural environments threaten the range of ecosystem services that support all life on this planet [Sleeman 2019: 91].
…
It was the challenge of responding to these complex [environmental] problems that led to the emergence of the concept of One Health, which is defined by the Centers for Disease Control and Prevention (CDC) as the collaborative effort of multiple disciplines and sectors – working locally, nationally, regionally and globally – with the goal of achieving optimal health outcomes, recognizing the interconnection among people, animals, plants and our shared environment. This definition acknowledges that human, domestic animal and wildlife health are interconnected within the context of ecosystem/environmental health and provides a useful conceptual framework for the development of solutions to global health and environmental challenges. Given this interconnection, it follows that actions aimed primarily at improving the health of one part of the human-animal-environmental triad may have unanticipated consequences for the system as a whole if the harms they may cause to the other components are not considered. However, previous authors have noted that, despite the acknowledged interdependencies, few public or livestock health interventions include a consideration of biodiversity conservation or ecosystem/environmental health. Instead, health-promoting interventions focus largely on single-sector outcomes and, thus, may miss the opportunity to concurrently optimize outcomes in the other two sectors [Sleeman 2019: 92].
The authors suggest that despite its potential, the One Health approach does not as yet fully integrate wildlife and environmental health, instead favoring human health. Yet failure to optimize the health of all three realms can lead to unexpected and outcomes, ironically increasing risk to humans in some cases. Therefore, the authors propose the clarification of One Health values and goals, and integration of a systems approach and a harm reduction perspective into the One Health framework.
Systems biology provides methods to understand how interactions among [interrelated and interdependent] parts [livestock, humans and wildlife, for example] give rise to the function and behavior of that system [Sleeman 2019: 96].
A harm reduction perspective recognizes that solutions to complex problems require a broad societal response and that elimination of risk is not feasible for most issues. Consequently, this perspective promotes collaborative, multisectoral approaches whereby reducing harm, despite uncertainty regarding the outcome, is valued over inaction spurred by a desire for a perfect solution [Sleeman 2019: 94].
Habitat loss reduces biodiversity, which leads to infectious disease emergence. The way a habitat is fragmented (how many patches it is divided into, how those patches are shaped, and what the distance is between them) further affects the extent of disease emergence. Both the number of divisions of habitat into smaller patches and the irregularity of patch shapes tend to increase habitat perimeter, which in turn increases contact between disease agents and humans.
The hazard is greatest in places with greater pre-existing biodiversity, where there is a greater diversity of microbial pathogens and associated hosts. There is a double risk of developing wilderness areas in these places because there are more pathogens to begin with, and the resulting biodiversity loss tends to amplify disease transmission.
Human encroachment into species-rich habitats may simultaneously decrease biodiversity and increase exposure of people to novel microbes [Wilkinson 2018: 1].
Ebola virus disease outbreaks in West and Central Africa have been linked to spillover from potential disease reservoirs such as bats, apes, and duikers (an antelope-like animal). Spillover has been thought to be related to population density, vegetation cover, and human activities such as hunting, poaching, and bushmeat consumption. In this study, forest data from satellites coupled with disease outbreak records identify a nexus between forest fragmentation and Ebola.
The researchers identified 11 sites of the first human infection of Ebola from a wild species having occurred since 2004. Changes in forest cover between the year 2000 (baseline year) and the years of first infection for each of these outbreaks were determined using high-resolution satellite data on tree cover. All 11 centers of infection were found to be located in forested areas where the rate of forest fragmentation was greater than the regional average. Similarly, forest fragmentation decreased with increasing distance from the centers of infection.
All 11 centers of infection were found to be located in forested areas where the rate of forest fragmentation was greater than the regional average.
The centers of first infection … tend to occur in areas where on the outbreak year the average degree of forest fragmentation (e.g., within a 25 km, 50 km or 100 km distance from the infection center) was significantly higher than in the rest of the region [Rulli 2017: 2].
Furthermore, eight of the 11 centers of infection were located in fragmentation “hotspots,” meaning within a cluster of highly fragmented forest areas.
Bats are the commonly accepted host to filoviruses such as Ebola and tend to increase in population in fragmented habitats. The geographic distribution of potential bat hosts was consistent with the distribution of the zoonotic niche of Ebola. A decline in the population of insectivorous bats and an increase in the frugivorous (fruit-eating) bat species as a result of forest fragmentation was observed. Reshaping forest boundaries, habitat disruption, and biodiversity loss may enhance the likelihood of zoonotic infection by increasing the abundance of a particular species and thus the prevalence of that species’ pathogens.
The fact that spillover tends to occur in hotspots of forest fragmentation rather than in clearcut areas suggests that chances of human interactions with host wildlife are higher in areas where human encroachment leaves forest fragments that provide habitat for reservoir species [Rulli 2017: 5].
Pressure on land and its products is increasingly pushing people into forested areas. Given the danger of zoonotic disease outbreak, any evaluation of the costs, risk, and benefits of forest loss and fragmentation should include global health considerations.
This article very pragmatically addresses the question of whether biodiversity conservation could be an effective public health tool against infectious disease emergence and transmission.
Determining whether biodiversity conservation is an effective public health strategy requires answering four questions: (1) Is there a general, causal relationship between host biodiversity and disease risk? (2) If the link is causal and negative for most pathogens, does the increased diversity of pathogens with more diverse host communities result in net total increase or decrease in infectious disease burden? (3) Is the net benefit of biodiversity conservation greater than the net benefit of diversity-degrading processes (agricultural land-use change and wild animal harvesting)? (4) Are conservation interventions feasible and cost-effective compared to standard public health approaches (vaccines and treatments)?
Regarding the first question, experimental and observational research shows that increased biodiversity is associated with reduced disease burden.
Overall, the available data suggests that there is some correlational support in many zoonotic systems for a dilution effect, and that some species or species groups are more important than others in transmission [Kilpatrick 2017: 4].
The dilution effect hypothesis originated to explain the Lyme disease system. Greater numbers of hosts that are less “competent” (at spreading Lyme disease) – opossums, birds, raccoons and skunks – dilutes the transmission of Lyme bacteria to larval ticks by more competent hosts – white-footed mice, eastern chipmunks and shrews. Changes in community diversity affect, for example, host-vector encounter rates and host and vector abundances.
However,
much more research is needed to show that observed correlations are causal and to identify the mechanisms by which diversity is influencing disease risk [Kilpatrick 2017: 4].
The possibility of confounding factors in observational field studies is high because the same disturbances that change host diversity alters other aspects of transmission as well. For example, an ecosystem disturbance may, in addition to decreasing host diversity, also increase vector abundance, making it difficult to discern the proximate cause of increased disease rates. The authors note that the dilution effect may well cause decreased disease rates – more research is needed to determine this. But they caution that if the dilution effect turns out not to be the direct cause of decreased disease rates in any given pathogen system, then interventions to increase host diversity could be in vain with respect to that desired outcome.
Examples of potential conservation interventions to improve public health include preserving or restoring forest land, reintroducing top predators to control host populations, installing bat or owl boxes to increase predation of mosquitos (vectors) or rodents (hosts). Our still limited understanding of the mechanisms driving disease incidence patterns, however, make it difficult to predict outcomes for broad-scale land-use interventions, according to the authors. They argue instead that more targeted interventions aiming to reduce populations of key hosts in transmission may be more feasible public health tools than general land preservation. Even this, however, requires “deep understanding of both disease and population ecology.”
Further research to address this knowledge gap may be worth the investment, both for human wellbeing and for the planet. Exposure to nature has been shown to improve human mental and physical health and wellbeing, the authors note, regardless of biodiversity’s potential to reduce infectious disease. Furthermore,
If diverse communities can be shown to provide net benefits to human wellbeing, this could provide a powerful motivation for preserving Earth’s remaining biodiversity [Kilpatrick 2017: 7].
This status-quo-challenging editorial is written for the American Society of Clinical Pathology, a group seemingly unrelated to the Bio4Climate community. The authors suggest that medical training in pathology over-emphasizes oncology at the expense of an adequate coverage of infectious disease, even though “between 1940 and 2004, a total of 335 human infectious diseases ‘emerged,’ and 60% of these were zoonotic” [Granter 2016: 645]. Having explained biodiversity loss as a factor driving disease rates, the authors make a plea for diagnosticians to become aware of the human health implications of environmental destruction.
Knowledge and prowess with infectious diseases for diagnosticians must be incorporated back into training with a reimagined lens crafted from the information we have gained by studying our environment, its destruction, and the ultimate resulting human infections. As loss of habitat, habitat fragmentation, and consequent biodiversity loss continue unabated, tools and skills will need to be in the hands of all diagnosticians if we hope to minimize the effect of these infections as they continually emerge [Granter 2016: 645].
This paper provides a particularly clear explanation of how biodiversity loss increases human infection risk.
The relationship between loss of biodiversity and human disease was first illustrated by Lyme disease. Its cause, the Borrelia burgdorferi bacterium, has the opportunity to encounter numerous vertebrate hosts – in one study estimated to be at least 125 species – in a diverse and healthy ecosystem. The potential hosts vary tremendously in their ability to harbor and transmit the bacteria, that is, their “reservoir competence.” Studies estimate the white-footed mouse infects more than 90% of ticks that complete their blood meal. While a few other hosts, such as eastern chipmunks and short-tailed shrews, are moderately competent, most tick hosts are marginally competent or dead-end hosts that are highly unlikely to transmit the infection. Since the white-footed mouse tends to thrive in impoverished ecosystems lacking biodiversity, infected ticks and, consequently, risk of human infection show a strong negative relationship with biodiversity. Because a diverse ecosystem with a range of vertebrate hosts – including many incompetent and dead-end hosts – “dilutes” the representation of the white-footed mouse and reduces human infection risk, this phenomenon has been termed the dilution effect [Granter 2016: 644].
“Because a diverse ecosystem with a range of vertebrate hosts – including many incompetent and dead-end hosts – “dilutes” the representation of the white-footed mouse and reduces human infection risk, this phenomenon has been termed the dilution effect” [Granter 2016: 644].
Human activities are dramatically reducing biodiversity, and the frequency and severity of infectious disease outbreaks in human, wildlife, and domesticated species are increasing. These concurrent patterns have prompted suggestions that biodiversity and the spread of diseases may be causally linked. For example, the dilution effect hypothesis proposes that diverse host communities inhibit the abundance of parasites through several mechanisms, such as regulating populations of susceptible hosts or interfering with the transmission process. Thus, diverse communities may inhibit the proliferation of parasites, thereby promoting the stability of ecological communities and ecosystem services (e.g., nutrient cycling, carbon sequestration, and natural product production) [Civitello 2015: 8667].
This meta-analysis concludes that as a general rule across ecosystems, biodiversity inhibits parasitism. Previous studies had focused on particular host-parasite systems, and found that greater host diversity dilutes, or limits, the spread of disease. “Consequently, anthropogenic declines in biodiversity could increase human and wildlife diseases and decrease crop and forest production” [Civitello 2015: 8667].
This paper contextualizes reduced transmission of infectious disease as one of the many ecosystem services provided by biodiversity. Changes in biodiversity affect infectious disease transmission by changing the abundance of the host and/or vector; the loss of non-host species may increase the density of host species, increasing the encounter rates between pathogen and host.
Often, the species that remain when biodiversity is lost are those which are better pathogen hosts, while the lost species tend to be more resistant to infectious disease.
In several case studies, the species most likely to be lost from ecological communities as diversity declines are those most likely to reduce pathogen transmission [Keesing 2010: 648].
For example, the white-footed mouse, which are high-quality hosts both for the bacteria causing Lyme Disease and for the tick vectors, are abundant in both biodiverse systems and impoverished systems, while opossums, a poorer host for the Lyme bacterium that also kill/eat most ticks attempting to feed on them, do poorly in lower-biodiversity conditions.
Therefore, as biodiversity is lost, the host with a strong buffering effect – the opossum – disappears, while the host with a strong amplifying effect – the mouse – remains [Keesing 2010: 650].
There may be a causal link between a species’ susceptibility to biodiversity loss and its quality as a disease host. Among vertebrates,
resilience in the face of disturbances that cause biodiversity loss, such as habitat destruction and fragmentation, is facilitated by life history features such as high reproductive output and intrinsic rates of increase. Vertebrates with these features tend to invest minimally in some aspects of adaptive immunity; we hypothesize that this may make them more competent hosts for pathogens and vectors [Keesing 2010: 650].
Biodiversity also affects the emergence of infectious disease, such as the evolution of a new strain of pathogen in the same host (due to antibiotic resistance, for example), and the spillover to a new host species. Pathogen establishment in humans from other animal hosts is related to mammal species richness (a larger source pool), and land-use change (such as deforestation), which increases contact between humans and pathogen hosts. The pathogen then becomes an epidemic due to the new host species’ density (domesticated animals and humans).
The authors recommend preserving biodiversity by protecting natural habitat, while also preserving microbial diversity within organisms by limiting the use of antimicrobial agents. A diverse microbiome within an organism serves as a buffer against pathogens.
West Nile Virus is an infectious disease that arrived in New York City in 1999, and subsequently spread across the country to the west coast. It is transmitted to humans from passerine (perching) birds via mosquito vectors. This study tested the dilution effect hypothesis, which posits that greater diversity (of birds in this case) reduces the concentration of species that are the primary disease reservoirs (American robin, American crow, blue jay, western scrub jay, common grackle, house finch, and house sparrow), thus reducing vector contact with infected individuals, and ultimately transmission to humans. The study analyzed the incidence of human infection during 2003-2004, and found that biodiversity was indeed associated with reduced WNV infection rates among humans.
For all 3 years, the county-level human incidence of WNV disease was strongly, and significantly, negatively correlated with bird diversity within that county [Ostfeld 2009: 41].
Similar results are reported for studies of the dilution effect of biodiversity on Lyme disease risk. Furthermore, having collected data on the competence of various mammalian hosts to infect ticks with Lyme disease, as well as each host species’ average tick burden, the authors state that “we can project the number of ticks that will feed on them and the proportion of those ticks that will become infected” [Ostfeld 2009: 42].
We conclude from these studies that high vertebrate diversity is negatively correlated with human risk of exposure to Lyme disease. Furthermore, knowledge of the species composition of these communities, beyond simple measures of species richness or evenness, strongly enhances our ability to predict risk [Ostfeld 2009: 42].
In summary,
Evidence for a protective dilution effect of high diversity has been obtained for numerous infectious diseases of humans, wildlife, and plants. The weight of evidence suggests that protection against exposure to infectious diseases should be added to the list of utilitarian functions of biodiversity [Ostfeld 2009: 42].
“Evidence for a protective dilution effect of high diversity has been obtained for numerous infectious diseases of humans, wildlife, and plants. The weight of evidence suggests that protection against exposure to infectious diseases should be added to the list of utilitarian functions of biodiversity [Ostfeld 2009: 42].
This review article describes the potential mechanisms by which biodiversity affects disease risk. The authors explore the mechanisms at play in simple systems with only host and pathogen, as well as in more complex systems that include a vector species and/or multiple hosts. The reduction of disease risk by increased diversity is called the “dilution effect.” The opposite, termed the “amplification effect,” is when disease risk increases. “Both models and literature reviews suggest that high host diversity is more likely to decrease than increase disease risk” [Keesing 2006: 485].
The mechanisms by which diversity affects disease risk are as follows:
Encounter reduction: An additional species (such as a predator) suppresses the movement of host species or vector species, thereby reducing contact between susceptible hosts and infected hosts or vectors. (Alternatively, if the presence of a different species causes host species to clump together more among their own kind, then transmission could increase in an encounter augmentation.)
Transmission reduction: An additional species in a system (such as a prey) reduces host stress, boosting immune system response and lowering pathogen load. An added species could also modify host behavior in a way that reduces the duration of their encounters and thus limits transmission.
Vector or susceptible host [population] regulation: The addition of any species that reduces birth rates or increases death rates, limiting overall population, among hosts susceptible to the pathogen or among pathogen vectors. Transmission rates may be reduced, for example, with the addition of host species predators or with the addition of species that attract vectors (ticks, for instance), but then groom themselves in a way that kills many vector individuals.
Infected host mortality: An added species outcompetes infected hosts for resources or targets infected hosts for predation.
Recovery augmentation: The addition of a prey species as an added resource for host species could, for example, increase full recovery rates of host species, creating a dilution effect, or, by contrast, increase the longevity of sick hosts in an amplification effect.
When there are many hosts for a particular pathogen, some species transmit the disease more readily than others. Often, the species that most effectively spread the disease (the most competent reservoirs) are present in species-poor, degraded ecosystems, meaning that any additional host species is likely to dilute the presences of the more contagious species.
One key question in multi-host disease systems is whether the most competent reservoir is present in species-poor communities. If so, species added to these communities have, by definition, lower (if any) reservoir competence and thus have the potential to decrease disease risk. If the most competent reservoir is not present in species-poor communities, by contrast, then an increase in diversity could include the addition of the most competent reservoir itself, which is likely to result in an amplification of disease risk. Ostfeld & Keesing (2000b) considered evidence that the most competent reservoir for a variety of vector-borne zoonoses was typically present in species-poor communities [Keesing 2006: 495].
Humans are not the only species to suffer global pandemics. Planetwide, fungal disease ravages amphibians, just as honeybees are ravaged by varroasis. A herpes virus caused mass mortality of pilchard fish off the coast of Australia and New Zealand in 1995, and seals from Antarctica to the Caspian Sea have contracted canine distemper viruses, for which domestic dogs are also hosts.
The authors point to multiple anthropogenic environmental changes as the underlying causes of disease emergence among wildlife, livestock and humans.
Emerging infectious diseases (EIDs) are defined as diseases that have recently increased in incidence or geographic range, recently moved into new host populations, recently been discovered or are caused by newly-evolved pathogens [Daszak 2001: 103].
Two major known causes of disease emergence in wildlife are (1) “spillover” of livestock disease into wild populations; and (2) pathogen pollution, which stems from the global transport of domestic and wild animals, and contaminated products and materials. In addition, habitat destruction and fragmentation, and toxic pollution, are likely to contribute to disease emergence, although these factors hadn’t been as well studied (at least at the time of the writing in 2001).
The authors conclude with the following observation:
We have described a group of wildlife diseases that can be classified as emerging in the same way as human EIDs. These represent a link in the chain of emergence of human and domestic animal diseases, with pathogens, habitats and environmental changes shared between these populations. Parallels between causes of emergence across these groups of diseases demonstrates an important concept: that human environmental change may be the most significant driver of wildlife, domestic animal and human EIDs [Daszak 2001: 112].
Anthropogenic environmental change and the emergence of infectious diseases in wildlife, Daszak, Cunningham & Hyatt 2001
Humans are not the only species to suffer global pandemics. Planetwide, fungal disease ravages amphibians, just as honeybees are ravaged by varroasis. A herpes virus caused mass mortality of pilchard fish off the coast of Australia and New Zealand in 1995, and seals from Antarctica to the Caspian Sea have contracted canine distemper viruses, for which domestic dogs are also hosts.
The authors point to multiple anthropogenic environmental changes as the underlying causes of disease emergence among wildlife, livestock and humans.
Emerging infectious diseases (EIDs) are defined as diseases that have recently increased in incidence or geographic range, recently moved into new host populations, recently been discovered or are caused by newly-evolved pathogens [Daszak 2001: 103].
Two major known causes of disease emergence in wildlife are (1) “spillover” of livestock disease into wild populations; and (2) pathogen pollution, which stems from the global transport of domestic and wild animals, and contaminated products and materials. In addition, habitat destruction and fragmentation, and toxic pollution, are likely to contribute to disease emergence, although these factors hadn’t been as well studied (at least at the time of the writing in 2001).
The authors conclude with the following observation:
We have described a group of wildlife diseases that can be classified as emerging in the same way as human EIDs. These represent a link in the chain of emergence of human and domestic animal diseases, with pathogens, habitats and environmental changes shared between these populations. Parallels between causes of emergence across these groups of diseases demonstrates an important concept: that human environmental change may be the most significant driver of wildlife, domestic animal and human EIDs [Daszak 2001: 112].
Effects of species diversity on disease risk, Keesing, Holt & Ostfeld 2006
This review article describes the potential mechanisms by which biodiversity affects disease risk. The authors explore the mechanisms at play in simple systems with only host and pathogen, as well as in more complex systems that include a vector species and/or multiple hosts. The reduction of disease risk by increased diversity is called the “dilution effect.” The opposite, termed the “amplification effect,” is when disease risk increases. “Both models and literature reviews suggest that high host diversity is more likely to decrease than increase disease risk” [Keesing 2006: 485].
The mechanisms by which diversity affects disease risk are as follows:
Encounter reduction: An additional species (such as a predator) suppresses the movement of host species or vector species, thereby reducing contact between susceptible hosts and infected hosts or vectors. (Alternatively, if the presence of a different species causes host species to clump together more among their own kind, then transmission could increase in an encounter augmentation.)
Transmission reduction: An additional species in a system (such as a prey) reduces host stress, boosting immune system response and lowering pathogen load. An added species could also modify host behavior in a way that reduces the duration of their encounters and thus limits transmission.
Vector or susceptible host [population] regulation: The addition of any species that reduces birth rates or increases death rates, limiting overall population, among hosts susceptible to the pathogen or among pathogen vectors. Transmission rates may be reduced, for example, with the addition of host species predators or with the addition of species that attract vectors (ticks, for instance), but then groom themselves in a way that kills many vector individuals.
Infected host mortality: An added species outcompetes infected hosts for resources or targets infected hosts for predation.
Recovery augmentation: The addition of a prey species as an added resource for host species could, for example, increase full recovery rates of host species, creating a dilution effect, or, by contrast, increase the longevity of sick hosts in an amplification effect.
When there are many hosts for a particular pathogen, some species transmit the disease more readily than others. Often, the species that most effectively spread the disease (the most competent reservoirs) are present in species-poor, degraded ecosystems, meaning that any additional host species is likely to dilute the presences of the more contagious species.
One key question in multi-host disease systems is whether the most competent reservoir is present in species-poor communities. If so, species added to these communities have, by definition, lower (if any) reservoir competence and thus have the potential to decrease disease risk. If the most competent reservoir is not present in species-poor communities, by contrast, then an increase in diversity could include the addition of the most competent reservoir itself, which is likely to result in an amplification of disease risk. Ostfeld & Keesing (2000b) considered evidence that the most competent reservoir for a variety of vector-borne zoonoses was typically present in species-poor communities [Keesing 2006: 495].
Biodiversity loss and the rise of zoonotic pathogens, Ostfeld 2009
West Nile Virus is an infectious disease that arrived in New York City in 1999, and subsequently spread across the country to the west coast. It is transmitted to humans from passerine (perching) birds via mosquito vectors. This study tested the dilution effect hypothesis, which posits that greater diversity (of birds in this case) reduces the concentration of species that are the primary disease reservoirs (American robin, American crow, blue jay, western scrub jay, common grackle, house finch, and house sparrow), thus reducing vector contact with infected individuals, and ultimately transmission to humans. The study analyzed the incidence of human infection during 2003-2004, and found that biodiversity was indeed associated with reduced WNV infection rates among humans.
For all 3 years, the county-level human incidence of WNV disease was strongly, and significantly, negatively correlated with bird diversity within that county [Ostfeld 2009: 41].
Similar results are reported for studies of the dilution effect of biodiversity on Lyme disease risk. Furthermore, having collected data on the competence of various mammalian hosts to infect ticks with Lyme disease, as well as each host species’ average tick burden, the authors state that “we can project the number of ticks that will feed on them and the proportion of those ticks that will become infected” [Ostfeld 2009: 42].
We conclude from these studies that high vertebrate diversity is negatively correlated with human risk of exposure to Lyme disease. Furthermore, knowledge of the species composition of these communities, beyond simple measures of species richness or evenness, strongly enhances our ability to predict risk [Ostfeld 2009: 42].
In summary,
Evidence for a protective dilution effect of high diversity has been obtained for numerous infectious diseases of humans, wildlife, and plants. The weight of evidence suggests that protection against exposure to infectious diseases should be added to the list of utilitarian functions of biodiversity [Ostfeld 2009: 42].
“Evidence for a protective dilution effect of high diversity has been obtained for numerous infectious diseases of humans, wildlife, and plants. The weight of evidence suggests that protection against exposure to infectious diseases should be added to the list of utilitarian functions of biodiversity [Ostfeld 2009: 42].
Impacts of biodiversity on the emergence and transmission of infectious diseases, Keesing et al. 2010
This paper contextualizes reduced transmission of infectious disease as one of the many ecosystem services provided by biodiversity. Changes in biodiversity affect infectious disease transmission by changing the abundance of the host and/or vector; the loss of non-host species may increase the density of host species, increasing the encounter rates between pathogen and host.
Often, the species that remain when biodiversity is lost are those which are better pathogen hosts, while the lost species tend to be more resistant to infectious disease.
In several case studies, the species most likely to be lost from ecological communities as diversity declines are those most likely to reduce pathogen transmission [Keesing 2010: 648].
For example, the white-footed mouse, which are high-quality hosts both for the bacteria causing Lyme Disease and for the tick vectors, are abundant in both biodiverse systems and impoverished systems, while opossums, a poorer host for the Lyme bacterium that also kill/eat most ticks attempting to feed on them, do poorly in lower-biodiversity conditions.
Therefore, as biodiversity is lost, the host with a strong buffering effect – the opossum – disappears, while the host with a strong amplifying effect – the mouse – remains [Keesing 2010: 650].
There may be a causal link between a species’ susceptibility to biodiversity loss and its quality as a disease host. Among vertebrates,
resilience in the face of disturbances that cause biodiversity loss, such as habitat destruction and fragmentation, is facilitated by life history features such as high reproductive output and intrinsic rates of increase. Vertebrates with these features tend to invest minimally in some aspects of adaptive immunity; we hypothesize that this may make them more competent hosts for pathogens and vectors [Keesing 2010: 650].
Biodiversity also affects the emergence of infectious disease, such as the evolution of a new strain of pathogen in the same host (due to antibiotic resistance, for example), and the spillover to a new host species. Pathogen establishment in humans from other animal hosts is related to mammal species richness (a larger source pool), and land-use change (such as deforestation), which increases contact between humans and pathogen hosts. The pathogen then becomes an epidemic due to the new host species’ density (domesticated animals and humans).
The authors recommend preserving biodiversity by protecting natural habitat, while also preserving microbial diversity within organisms by limiting the use of antimicrobial agents. A diverse microbiome within an organism serves as a buffer against pathogens.
Biodiversity inhibits parasites: Broad evidence for the dilution effect, Civitello et al. 2015
Human activities are dramatically reducing biodiversity, and the frequency and severity of infectious disease outbreaks in human, wildlife, and domesticated species are increasing. These concurrent patterns have prompted suggestions that biodiversity and the spread of diseases may be causally linked. For example, the dilution effect hypothesis proposes that diverse host communities inhibit the abundance of parasites through several mechanisms, such as regulating populations of susceptible hosts or interfering with the transmission process. Thus, diverse communities may inhibit the proliferation of parasites, thereby promoting the stability of ecological communities and ecosystem services (e.g., nutrient cycling, carbon sequestration, and natural product production) [Civitello 2015: 8667].
This meta-analysis concludes that as a general rule across ecosystems, biodiversity inhibits parasitism. Previous studies had focused on particular host-parasite systems, and found that greater host diversity dilutes, or limits, the spread of disease. “Consequently, anthropogenic declines in biodiversity could increase human and wildlife diseases and decrease crop and forest production” [Civitello 2015: 8667].
Where the Wild Things Aren’t: Loss of Biodiversity, Emerging Infectious Diseases, and Implications for Diagnosticians, Granter 2016
This status-quo-challenging editorial is written for the American Society of Clinical Pathology, a group seemingly unrelated to the Bio4Climate community. The authors suggest that medical training in pathology over-emphasizes oncology at the expense of an adequate coverage of infectious disease, even though “between 1940 and 2004, a total of 335 human infectious diseases ‘emerged,’ and 60% of these were zoonotic” [Granter 2016: 645]. Having explained biodiversity loss as a factor driving disease rates, the authors make a plea for diagnosticians to become aware of the human health implications of environmental destruction.
Knowledge and prowess with infectious diseases for diagnosticians must be incorporated back into training with a reimagined lens crafted from the information we have gained by studying our environment, its destruction, and the ultimate resulting human infections. As loss of habitat, habitat fragmentation, and consequent biodiversity loss continue unabated, tools and skills will need to be in the hands of all diagnosticians if we hope to minimize the effect of these infections as they continually emerge [Granter 2016: 645].
This paper provides a particularly clear explanation of how biodiversity loss increases human infection risk.
The relationship between loss of biodiversity and human disease was first illustrated by Lyme disease. Its cause, the Borrelia burgdorferi bacterium, has the opportunity to encounter numerous vertebrate hosts – in one study estimated to be at least 125 species – in a diverse and healthy ecosystem. The potential hosts vary tremendously in their ability to harbor and transmit the bacteria, that is, their “reservoir competence.” Studies estimate the white-footed mouse infects more than 90% of ticks that complete their blood meal. While a few other hosts, such as eastern chipmunks and short-tailed shrews, are moderately competent, most tick hosts are marginally competent or dead-end hosts that are highly unlikely to transmit the infection. Since the white-footed mouse tends to thrive in impoverished ecosystems lacking biodiversity, infected ticks and, consequently, risk of human infection show a strong negative relationship with biodiversity. Because a diverse ecosystem with a range of vertebrate hosts – including many incompetent and dead-end hosts – “dilutes” the representation of the white-footed mouse and reduces human infection risk, this phenomenon has been termed the dilution effect [Granter 2016: 644].
“Because a diverse ecosystem with a range of vertebrate hosts – including many incompetent and dead-end hosts – “dilutes” the representation of the white-footed mouse and reduces human infection risk, this phenomenon has been termed the dilution effect” [Granter 2016: 644].
Conservation of biodiversity as a strategy for improving human health and wellbeing, Kilpatrick et al. 2017
This article very pragmatically addresses the question of whether biodiversity conservation could be an effective public health tool against infectious disease emergence and transmission.
Determining whether biodiversity conservation is an effective public health strategy requires answering four questions: (1) Is there a general, causal relationship between host biodiversity and disease risk? (2) If the link is causal and negative for most pathogens, does the increased diversity of pathogens with more diverse host communities result in net total increase or decrease in infectious disease burden? (3) Is the net benefit of biodiversity conservation greater than the net benefit of diversity-degrading processes (agricultural land-use change and wild animal harvesting)? (4) Are conservation interventions feasible and cost-effective compared to standard public health approaches (vaccines and treatments)?
Regarding the first question, experimental and observational research shows that increased biodiversity is associated with reduced disease burden.
Overall, the available data suggests that there is some correlational support in many zoonotic systems for a dilution effect, and that some species or species groups are more important than others in transmission [Kilpatrick 2017: 4].
The dilution effect hypothesis originated to explain the Lyme disease system. Greater numbers of hosts that are less “competent” (at spreading Lyme disease) – opossums, birds, raccoons and skunks – dilutes the transmission of Lyme bacteria to larval ticks by more competent hosts – white-footed mice, eastern chipmunks and shrews. Changes in community diversity affect, for example, host-vector encounter rates and host and vector abundances.
However,
much more research is needed to show that observed correlations are causal and to identify the mechanisms by which diversity is influencing disease risk [Kilpatrick 2017: 4].
The possibility of confounding factors in observational field studies is high because the same disturbances that change host diversity alters other aspects of transmission as well. For example, an ecosystem disturbance may, in addition to decreasing host diversity, also increase vector abundance, making it difficult to discern the proximate cause of increased disease rates. The authors note that the dilution effect may well cause decreased disease rates – more research is needed to determine this. But they caution that if the dilution effect turns out not to be the direct cause of decreased disease rates in any given pathogen system, then interventions to increase host diversity could be in vain with respect to that desired outcome.
Examples of potential conservation interventions to improve public health include preserving or restoring forest land, reintroducing top predators to control host populations, installing bat or owl boxes to increase predation of mosquitos (vectors) or rodents (hosts). Our still limited understanding of the mechanisms driving disease incidence patterns, however, make it difficult to predict outcomes for broad-scale land-use interventions, according to the authors. They argue instead that more targeted interventions aiming to reduce populations of key hosts in transmission may be more feasible public health tools than general land preservation. Even this, however, requires “deep understanding of both disease and population ecology.”
Further research to address this knowledge gap may be worth the investment, both for human wellbeing and for the planet. Exposure to nature has been shown to improve human mental and physical health and wellbeing, the authors note, regardless of biodiversity’s potential to reduce infectious disease. Furthermore,
If diverse communities can be shown to provide net benefits to human wellbeing, this could provide a powerful motivation for preserving Earth’s remaining biodiversity [Kilpatrick 2017: 7].
The nexus between forest fragmentation in Africa and Ebola virus disease outbreaks, Rulli et al. 2017
Ebola virus disease outbreaks in West and Central Africa have been linked to spillover from potential disease reservoirs such as bats, apes, and duikers (an antelope-like animal). Spillover has been thought to be related to population density, vegetation cover, and human activities such as hunting, poaching, and bushmeat consumption. In this study, forest data from satellites coupled with disease outbreak records identify a nexus between forest fragmentation and Ebola.
The researchers identified 11 sites of the first human infection of Ebola from a wild species having occurred since 2004. Changes in forest cover between the year 2000 (baseline year) and the years of first infection for each of these outbreaks were determined using high-resolution satellite data on tree cover. All 11 centers of infection were found to be located in forested areas where the rate of forest fragmentation was greater than the regional average. Similarly, forest fragmentation decreased with increasing distance from the centers of infection.
All 11 centers of infection were found to be located in forested areas where the rate of forest fragmentation was greater than the regional average.
The centers of first infection … tend to occur in areas where on the outbreak year the average degree of forest fragmentation (e.g., within a 25 km, 50 km or 100 km distance from the infection center) was significantly higher than in the rest of the region [Rulli 2017: 2].
Furthermore, eight of the 11 centers of infection were located in fragmentation “hotspots,” meaning within a cluster of highly fragmented forest areas.
Bats are the commonly accepted host to filoviruses such as Ebola and tend to increase in population in fragmented habitats. The geographic distribution of potential bat hosts was consistent with the distribution of the zoonotic niche of Ebola. A decline in the population of insectivorous bats and an increase in the frugivorous (fruit-eating) bat species as a result of forest fragmentation was observed. Reshaping forest boundaries, habitat disruption, and biodiversity loss may enhance the likelihood of zoonotic infection by increasing the abundance of a particular species and thus the prevalence of that species’ pathogens.
The fact that spillover tends to occur in hotspots of forest fragmentation rather than in clearcut areas suggests that chances of human interactions with host wildlife are higher in areas where human encroachment leaves forest fragments that provide habitat for reservoir species [Rulli 2017: 5].
Pressure on land and its products is increasingly pushing people into forested areas. Given the danger of zoonotic disease outbreak, any evaluation of the costs, risk, and benefits of forest loss and fragmentation should include global health considerations.
Habitat fragmentation, biodiversity loss and the risk of novel infectious disease emergence, Wilkinson 2018
Habitat loss reduces biodiversity, which leads to infectious disease emergence. The way a habitat is fragmented (how many patches it is divided into, how those patches are shaped, and what the distance is between them) further affects the extent of disease emergence. Both the number of divisions of habitat into smaller patches and the irregularity of patch shapes tend to increase habitat perimeter, which in turn increases contact between disease agents and humans.
The hazard is greatest in places with greater pre-existing biodiversity, where there is a greater diversity of microbial pathogens and associated hosts. There is a double risk of developing wilderness areas in these places because there are more pathogens to begin with, and the resulting biodiversity loss tends to amplify disease transmission.
Human encroachment into species-rich habitats may simultaneously decrease biodiversity and increase exposure of people to novel microbes [Wilkinson 2018: 1].
Integration of wildlife and environmental health into a One Health approach, Sleeman et al. 2019
This article introduces the concept of One Health, a public health framework adopted by the Centers for Disease Control in 2009, which recognizes the interdependence of humans, animals and our shared environment. The concept has gained traction as a way to address health problems arising from global environmental change.
Climate change, loss of biodiversity, habitat fragmentation and pollution, and subsequent degradation of natural environments threaten the range of ecosystem services that support all life on this planet [Sleeman 2019: 91].
…
It was the challenge of responding to these complex [environmental] problems that led to the emergence of the concept of One Health, which is defined by the Centers for Disease Control and Prevention (CDC) as the collaborative effort of multiple disciplines and sectors – working locally, nationally, regionally and globally – with the goal of achieving optimal health outcomes, recognizing the interconnection among people, animals, plants and our shared environment. This definition acknowledges that human, domestic animal and wildlife health are interconnected within the context of ecosystem/environmental health and provides a useful conceptual framework for the development of solutions to global health and environmental challenges. Given this interconnection, it follows that actions aimed primarily at improving the health of one part of the human-animal-environmental triad may have unanticipated consequences for the system as a whole if the harms they may cause to the other components are not considered. However, previous authors have noted that, despite the acknowledged interdependencies, few public or livestock health interventions include a consideration of biodiversity conservation or ecosystem/environmental health. Instead, health-promoting interventions focus largely on single-sector outcomes and, thus, may miss the opportunity to concurrently optimize outcomes in the other two sectors [Sleeman 2019: 92].
The authors suggest that despite its potential, the One Health approach does not as yet fully integrate wildlife and environmental health, instead favoring human health. Yet failure to optimize the health of all three realms can lead to unexpected and outcomes, ironically increasing risk to humans in some cases. Therefore, the authors propose the clarification of One Health values and goals, and integration of a systems approach and a harm reduction perspective into the One Health framework.
Systems biology provides methods to understand how interactions among [interrelated and interdependent] parts [livestock, humans and wildlife, for example] give rise to the function and behavior of that system [Sleeman 2019: 96].
A harm reduction perspective recognizes that solutions to complex problems require a broad societal response and that elimination of risk is not feasible for most issues. Consequently, this perspective promotes collaborative, multisectoral approaches whereby reducing harm, despite uncertainty regarding the outcome, is valued over inaction spurred by a desire for a perfect solution [Sleeman 2019: 94].
Emerging human infectious diseases and the links to global food production, Rohr et al. 2019
Increasing agricultural production to feed >11 billion people by 2100 raises several challenges for effectively managing infectious disease. Of many factors examined in this article linking agricultural expansion to infectious disease, one is conversion of natural habitat to cropland or rangeland. Land conversion increases contact between wild animals, livestock and humans.
As natural ecosystems are converted to crop land or range land, interactions among humans, and domesticated and wild animals, could increase. … These interactions are crucial because 77% of livestock pathogens are capable of infecting multiple host species, including wildlife and humans, and based on published estimates from the 2000s, over half of all recognized human pathogens are currently or originally zoonotic, as are 60–76% of recent emerging infectious disease events [Rohr 2019: 451].
“As natural ecosystems are converted to crop land or range land, interactions among humans, and domesticated and wild animals, could increase” [Rohr 2019: 451].
Land conversion pushes humans and livestock up against wilderness areas, increasing contact between species with previously little to no contact. The jumping of a pathogen to a new host species is called “spillover.”
Spillover appears to be a function of the frequency, duration and intimacy of interactions between a reservoir and novel host population. For example, influenza is believed to have jumped from horses to humans soon after domesticating horses and then made additional jumps to humans from other domesticated animals, such as poultry and swine [Rohr 2019: 451].
Furthermore, agricultural intensification tends to involve greater concentrations of a single variety of a single species, increasing the risk that any new disease will spread quickly in the population.
A central tenet of epidemiology is that the incidence of many infectious diseases should increase proportionally with host density because of increased contact rates and thus transmission among hosts. Hence, increasing human and livestock densities could cause increases in infectious diseases unless investments in disease prevention are sufficient to prevent these increases [Rohr 2019: 451].
Industrial-scale confined livestock production is
vulnerable to devastating losses of animals to disease. For instance, in just the last 25 years, an influenza A virus (H5N1) and a foot-and-mouth outbreak led to the destruction of more than 1.2 million chickens and 6 million livestock in China and Great Britain, respectively, and a ‘mad cow disease’ epizootic led to the slaughter of 11 million cattle worldwide [Rohr 2019: 449].
Increased agricultural production tends to be accompanied by new irrigation infrastructure and increased pesticide, fertilizer and antibiotic use, all of which increase infectious disease risk. Dams (often created for irrigation schemes) increase risk of mosquito-borne disease. Antibiotic overuse for livestock fosters resistance among pathogens that can also infect humans. Greater pesticide use leads to resistance among disease vectors such as mosquitoes to insecticides, while also weakening immune systems among exposed humans and wildlife hosts, increasing infection rates/severity. Nutrient enrichment caused by fertilizer can also contribute to the spread of infectious disease, for example, through mosquitos or snail vectors.
Finally, the urbanization and globalization associated with agricultural intensification/expansion elongates food supply chains, which increases movement of people and goods over borders, spreading food-born illness, flu and other infections.
In short,
These analyses revealed that agricultural drivers were associated with 25% of all diseases and nearly 50% of zoonotic diseases that emerged in humans since 1940. These values are even higher if we include the use of antimicrobial agents as an agricultural driver of human disease emergence, given that agricultural uses of antibiotics outpace medical uses in the developed world nearly nine to one [Rohr 2019: 451].
The authors recommend numerous measures for improving agricultural production while limiting infectious disease, including reducing antibiotic use for livestock, conserving biodiversity, improving and diversifying livestock and crop genetic material, investing in urban agriculture, social investments, and inter-disciplinary research and collaboration.
The subject of infectious disease became both fascinating and uncomfortably relevant with the global breakout of Covid-19 in early 2020. Are bats to blame, hunting and selling of wild game or seafood markets? It turns out that the destruction of nature is the root problem, according to the UN environment chief and lead scientists for the Intergovernmental Panel on Biodiversity and Ecosystem Services (IPBES). “Covid-19 is nature sending us a message,” writes the UN environment chief and colleague [Dasgupta & Anderson 2020]. They continue:
In fact, it reads like an SOS signal for the human enterprise, bringing into sharp focus the need to live within the planet’s means. The environmental, health and economic consequences of failing to do so are disastrous [Dasgupta & Anderson 2020].
“Covid-19 is nature sending us a message. In fact, it reads like an SOS signal for the human enterprise, bringing into sharp focus the need to live within the planet’s means. The environmental, health and economic consequences of failing to do so are disastrous.” – Sir Partha Dasgupta & Ingar Anderson
An April 2020 IPBES article [Settele 2020] echoes this alert:
As with the climate and biodiversity crises, recent pandemics are a direct consequence of human activity – particularly our global financial and economic systems, based on a limited paradigm that prizes economic growth at any cost.
… Our actions have significantly impacted more than three quarters of the Earth’s land surface, destroyed more than 85% of wetlands and dedicated more than a third of all land and almost 75% of available freshwater to crops and livestock production [Settele 2020].
“With more than 70% of all emerging diseases affecting people having originated in wildlife and domesticated animals,” the IPBES team explains, “activities that bring increasing numbers of people into direct contact and often conflict with the animals that carry these pathogens” lead to pandemics [Settele 2020].
A study [Rulli 2017] linking forest fragmentation with outbreaks of Ebola virus disease in West and Central Africa bears out this assessment. The authors stress that infectious disease emergence is among the many dangers stemming from ecosystem destruction.
The impact of forest loss on ecosystems and the services they provide is often evaluated in terms of habitat destruction, losses of biodiversity, carbon stock and emissions, land degradation, or altered climate and hydrologic conditions. This study, however, highlights that deforestation and forest fragmentation potentially have another important class of externalities associated with global health and zoonotic[3] disease outbreaks [Rulli 2017: 5].
Using satellite data on forest cover change from 2000 to 2014, these researchers found that all 11 sites of the first human Ebola infections since 2004 occurred in close proximity (within 25 km) to areas with higher rates of forest fragmentation than the regional average. Similarly, the closer one approached to the centers of first infection, the greater the fragmentation.
What is forest fragmentation? It is the cutting of forest into isolated patches and irregular shapes, resulting in greater lengths of edge between forested and non-forested areas. The increased length of forest perimeter in turn increases contact between humans and the wildlife that are potential disease agents, which would not otherwise be crossing our paths.
In addition to increasing human-wildlife contact, forest fragmentation favors some species while harming or obliterating others, throwing the whole system out of balance. Various studies have shown that the animals that thrive in degraded ecosystems are the same ones that constitute reservoirs of diseases communicable to humans. For example, in the case of West and Central Africa, the species suspected in the emergence of Ebola in humans include gorillas, chimpanzees, duikers (similar to an antelope) and a handful of bat species, all of which have been observed to increase in density following forest disturbance.
In addition to increasing human-wildlife contact, forest fragmentation favors some species while harming or obliterating others, throwing the whole system out of balance.
Thus, it could be argued that while disturbance by deforestation destroys the habitat of specialist[4] species, generalists – possibly including reservoirs of some zoonotic pathogens – thrive, thereby further enhancing the risk of infection in human populations close to the forest margins [Rulli 2017: 5].
A similar observation was made in North America. The white-footed mouse, an extremely “competent” host for Lyme Disease – meaning highly capable of harboring and transmitting the Lyme bacteria – does well in degraded forests, while many less competent hosts require more diverse, intact forests to thrive. Opossums, for instance, do poorly in impoverished ecosystems and also do not transmit the Lyme bacterium as readily as mice do. Furthermore, opossums kill ticks attempting to feed on them, making them a poorer host for the tick vector[5] as well.
There may be a direct link between a species’ susceptibility to habitat degradation and its quality as a disease host[6]. Among vertebrates,
resilience in the face of disturbances that cause biodiversity loss, such as habitat destruction and fragmentation, is facilitated by life history features such as high reproductive output and intrinsic rates of increase. Vertebrates with these features tend to invest minimally in some aspects of adaptive immunity; we hypothesize that this may make them more competent hosts for pathogens and vectors [Keesing 2010: 650].
The loss of less-competent disease host species, or of predators, for example, that would otherwise control a competent host population, thus allowing the latter to flourish, can create an “amplification effect.” The resulting higher concentration of competent hosts increases the likelihood of vector contact with infected hosts, thus increases disease transmission.
The amplification effect was observed also in the case of West Nile Virus (WNV) in North America in 2003-2004 [Ostfeld 2009]. Counties with higher passerine (perching) bird diversity were found to have lower human incidence of WNV disease, presumably due to lower concentrations of the bird species that were the primary disease reservoirs. With higher bird diversity, mosquito vectors were less likely to get infected due to a greater prevalence of uninfected birds upon which to feed.
With higher bird diversity, mosquito vectors were less likely to get infected due to a greater prevalence of uninfected birds upon which to feed.
Given multiple studies of particular disease systems like the ones described above indicating that biodiversity inhibits the spread of disease, another group of scientists [Civitello 2015] wanted to know how broadly and generally the amplification effect applies. Were these disease systems special cases, or do they suggest an inherent relationship between biodiversity and disease? Inverse of the amplification effect, “the dilution effect hypothesis suggests that diverse ecological communities limit disease spread via several mechanisms. Therefore, biodiversity losses could worsen epidemics that harm humans and wildlife” [Civitello 2015: 8667], the authors state to contextualize their research. They analyzed 202 studies of biodiversity and parasite abundance, and found “overwhelming evidence of dilution, which is independent of host density, study design, and type and specialization of parasites” [Civitello 2015: 8667]. From these results, it can be inferred that biodiversity generally limits infectious disease.
This message has not gone unheard by some in the public health field. An article [Granter 2016] published in the American Society of Clinical Pathology argues for health practitioners to become aware of the human health implications of environmental destruction, stating that:
Knowledge and prowess with infectious diseases for diagnosticians must be incorporated back into training with a reimagined lens crafted from the information we have gained by studying our environment, its destruction, and the ultimate resulting human infections [Granter 2016: 645].
A research team that included ecologists and a Center for Disease Control staff member [Kilpatrick 2017] analyzed the inclusion of biodiversity conservation among key public health tools. Without further research, the authors remain hesitant to recommend conservation generally as a public health tool (except insofar as exposure to nature boosts human wellbeing). However, they suggest that targeted interventions, such as reintroducing top predators to control host populations, installing bat or owl boxes to increase predation of mosquitos (vectors) or rodents (hosts) could be feasible public health interventions against infectious disease.
In their IPBES letter, Settele and colleagues [2020] recommend the adoption of a “One Health” approach to public health. This concept recognizes the fundamental interdependence of humans, animals, plants and our shared environment, and stresses the importance to human health of the overall health of nature. Some assessments paint that interdependence in even starker terms: “Zoonoses [human diseases of animal origin] reveal that environmental stewardship is not simply related to public health; in many cases, they are the same,” writes science journalist Ferris Jabr [2020].
In their IPBES letter, Settele and colleagues [2020] recommend the adoption of a “One Health” approach to public health. This concept recognizes the fundamental interdependence of humans, animals, plants and our shared environment, and stresses the importance to human health of the overall health of nature.
Indeed, humans are not the only species to experience disease epidemics. Bovine tuberculosis, honeybee varroasis, rabbit hemorrhagic disease virus, herpes virus in pilchard fish, West Nile virus in birds, and amphibians worldwide ravaged by a fungal disease are a few examples of how other species suffer. The underlying factors causing livestock, wildlife and human epidemics are anthropogenic environmental change driven by globalization of agriculture, commerce and human travel, all of which spread disease. Habitat destruction and toxic pollution are additional factors [Daszek 2001].
Compendium readers will be aware that biodiverse, intact ecosystems provide multiple vital functions. Healthy ecosystems absorb stormwater, forestall drought, generate rain, cool the air and land, purify air and groundwater, and pollinate crops, for example, not to mention contributing to human psychological wellbeing by providing recreational opportunities and beauty. What pandemics like Covid19 have now made us aware of is that biodiversity also plays a key role in limiting the emergence and spread of infectious disease.
Biodiversity loss and pandemics article summaries
Anthropogenic environmental change and the emergence of infectious diseases in wildlife, Daszak, Cunningham & Hyatt 2001
Humans are not the only species to suffer global pandemics. Planetwide, fungal disease ravages amphibians, just as honeybees are ravaged by varroasis. A herpes virus caused mass mortality of pilchard fish off the coast of Australia and New Zealand in 1995, and seals from Antarctica to the Caspian Sea have contracted canine distemper viruses, for which domestic dogs are also hosts.
The authors point to multiple anthropogenic environmental changes as the underlying causes of disease emergence among wildlife, livestock and humans.
Emerging infectious diseases (EIDs) are defined as diseases that have recently increased in incidence or geographic range, recently moved into new host populations, recently been discovered or are caused by newly-evolved pathogens [Daszak 2001: 103].
Two major known causes of disease emergence in wildlife are (1) “spillover” of livestock disease into wild populations; and (2) pathogen pollution, which stems from the global transport of domestic and wild animals, and contaminated products and materials. In addition, habitat destruction and fragmentation, and toxic pollution, are likely to contribute to disease emergence, although these factors hadn’t been as well studied (at least at the time of the writing in 2001).
The authors conclude with the following observation:
We have described a group of wildlife diseases that can be classified as emerging in the same way as human EIDs. These represent a link in the chain of emergence of human and domestic animal diseases, with pathogens, habitats and environmental changes shared between these populations. Parallels between causes of emergence across these groups of diseases demonstrates an important concept: that human environmental change may be the most significant driver of wildlife, domestic animal and human EIDs [Daszak 2001: 112].
Effects of species diversity on disease risk, Keesing, Holt & Ostfeld 2006
This review article describes the potential mechanisms by which biodiversity affects disease risk. The authors explore the mechanisms at play in simple systems with only host and pathogen, as well as in more complex systems that include a vector species and/or multiple hosts. The reduction of disease risk by increased diversity is called the “dilution effect.” The opposite, termed the “amplification effect,” is when disease risk increases. “Both models and literature reviews suggest that high host diversity is more likely to decrease than increase disease risk” [Keesing 2006: 485].
The mechanisms by which diversity affects disease risk are as follows:
Encounter reduction: An additional species (such as a predator) suppresses the movement of host species or vector species, thereby reducing contact between susceptible hosts and infected hosts or vectors. (Alternatively, if the presence of a different species causes host species to clump together more among their own kind, then transmission could increase in an encounter augmentation.)
Transmission reduction: An additional species in a system (such as a prey) reduces host stress, boosting immune system response and lowering pathogen load. An added species could also modify host behavior in a way that reduces the duration of their encounters and thus limits transmission.
Vector or susceptible host [population] regulation: The addition of any species that reduces birth rates or increases death rates, limiting overall population, among hosts susceptible to the pathogen or among pathogen vectors. Transmission rates may be reduced, for example, with the addition of host species predators or with the addition of species that attract vectors (ticks, for instance), but then groom themselves in a way that kills many vector individuals.
Infected host mortality: An added species outcompetes infected hosts for resources or targets infected hosts for predation.
Recovery augmentation: The addition of a prey species as an added resource for host species could, for example, increase full recovery rates of host species, creating a dilution effect, or, by contrast, increase the longevity of sick hosts in an amplification effect.
When there are many hosts for a particular pathogen, some species transmit the disease more readily than others. Often, the species that most effectively spread the disease (the most competent reservoirs) are present in species-poor, degraded ecosystems, meaning that any additional host species is likely to dilute the presences of the more contagious species.
One key question in multi-host disease systems is whether the most competent reservoir is present in species-poor communities. If so, species added to these communities have, by definition, lower (if any) reservoir competence and thus have the potential to decrease disease risk. If the most competent reservoir is not present in species-poor communities, by contrast, then an increase in diversity could include the addition of the most competent reservoir itself, which is likely to result in an amplification of disease risk. Ostfeld & Keesing (2000b) considered evidence that the most competent reservoir for a variety of vector-borne zoonoses was typically present in species-poor communities [Keesing 2006: 495].
Biodiversity loss and the rise of zoonotic pathogens, Ostfeld 2009
West Nile Virus is an infectious disease that arrived in New York City in 1999, and subsequently spread across the country to the west coast. It is transmitted to humans from passerine (perching) birds via mosquito vectors. This study tested the dilution effect hypothesis, which posits that greater diversity (of birds in this case) reduces the concentration of species that are the primary disease reservoirs (American robin, American crow, blue jay, western scrub jay, common grackle, house finch, and house sparrow), thus reducing vector contact with infected individuals, and ultimately transmission to humans. The study analyzed the incidence of human infection during 2003-2004, and found that biodiversity was indeed associated with reduced WNV infection rates among humans.
For all 3 years, the county-level human incidence of WNV disease was strongly, and significantly, negatively correlated with bird diversity within that county [Ostfeld 2009: 41].
Similar results are reported for studies of the dilution effect of biodiversity on Lyme disease risk. Furthermore, having collected data on the competence of various mammalian hosts to infect ticks with Lyme disease, as well as each host species’ average tick burden, the authors state that “we can project the number of ticks that will feed on them and the proportion of those ticks that will become infected” [Ostfeld 2009: 42].
We conclude from these studies that high vertebrate diversity is negatively correlated with human risk of exposure to Lyme disease. Furthermore, knowledge of the species composition of these communities, beyond simple measures of species richness or evenness, strongly enhances our ability to predict risk [Ostfeld 2009: 42].
In summary,
Evidence for a protective dilution effect of high diversity has been obtained for numerous infectious diseases of humans, wildlife, and plants. The weight of evidence suggests that protection against exposure to infectious diseases should be added to the list of utilitarian functions of biodiversity [Ostfeld 2009: 42].
“Evidence for a protective dilution effect of high diversity has been obtained for numerous infectious diseases of humans, wildlife, and plants. The weight of evidence suggests that protection against exposure to infectious diseases should be added to the list of utilitarian functions of biodiversity [Ostfeld 2009: 42].
Impacts of biodiversity on the emergence and transmission of infectious diseases, Keesing et al. 2010
This paper contextualizes reduced transmission of infectious disease as one of the many ecosystem services provided by biodiversity. Changes in biodiversity affect infectious disease transmission by changing the abundance of the host and/or vector; the loss of non-host species may increase the density of host species, increasing the encounter rates between pathogen and host.
Often, the species that remain when biodiversity is lost are those which are better pathogen hosts, while the lost species tend to be more resistant to infectious disease.
In several case studies, the species most likely to be lost from ecological communities as diversity declines are those most likely to reduce pathogen transmission [Keesing 2010: 648].
For example, the white-footed mouse, which are high-quality hosts both for the bacteria causing Lyme Disease and for the tick vectors, are abundant in both biodiverse systems and impoverished systems, while opossums, a poorer host for the Lyme bacterium that also kill/eat most ticks attempting to feed on them, do poorly in lower-biodiversity conditions.
Therefore, as biodiversity is lost, the host with a strong buffering effect – the opossum – disappears, while the host with a strong amplifying effect – the mouse – remains [Keesing 2010: 650].
There may be a causal link between a species’ susceptibility to biodiversity loss and its quality as a disease host. Among vertebrates,
resilience in the face of disturbances that cause biodiversity loss, such as habitat destruction and fragmentation, is facilitated by life history features such as high reproductive output and intrinsic rates of increase. Vertebrates with these features tend to invest minimally in some aspects of adaptive immunity; we hypothesize that this may make them more competent hosts for pathogens and vectors [Keesing 2010: 650].
Biodiversity also affects the emergence of infectious disease, such as the evolution of a new strain of pathogen in the same host (due to antibiotic resistance, for example), and the spillover to a new host species. Pathogen establishment in humans from other animal hosts is related to mammal species richness (a larger source pool), and land-use change (such as deforestation), which increases contact between humans and pathogen hosts. The pathogen then becomes an epidemic due to the new host species’ density (domesticated animals and humans).
The authors recommend preserving biodiversity by protecting natural habitat, while also preserving microbial diversity within organisms by limiting the use of antimicrobial agents. A diverse microbiome within an organism serves as a buffer against pathogens.
Biodiversity inhibits parasites: Broad evidence for the dilution effect, Civitello et al. 2015
Human activities are dramatically reducing biodiversity, and the frequency and severity of infectious disease outbreaks in human, wildlife, and domesticated species are increasing. These concurrent patterns have prompted suggestions that biodiversity and the spread of diseases may be causally linked. For example, the dilution effect hypothesis proposes that diverse host communities inhibit the abundance of parasites through several mechanisms, such as regulating populations of susceptible hosts or interfering with the transmission process. Thus, diverse communities may inhibit the proliferation of parasites, thereby promoting the stability of ecological communities and ecosystem services (e.g., nutrient cycling, carbon sequestration, and natural product production) [Civitello 2015: 8667].
This meta-analysis concludes that as a general rule across ecosystems, biodiversity inhibits parasitism. Previous studies had focused on particular host-parasite systems, and found that greater host diversity dilutes, or limits, the spread of disease. “Consequently, anthropogenic declines in biodiversity could increase human and wildlife diseases and decrease crop and forest production” [Civitello 2015: 8667].
Where the Wild Things Aren’t: Loss of Biodiversity, Emerging Infectious Diseases, and Implications for Diagnosticians, Granter 2016
This status-quo-challenging editorial is written for the American Society of Clinical Pathology, a group seemingly unrelated to the Bio4Climate community. The authors suggest that medical training in pathology over-emphasizes oncology at the expense of an adequate coverage of infectious disease, even though “between 1940 and 2004, a total of 335 human infectious diseases ‘emerged,’ and 60% of these were zoonotic” [Granter 2016: 645]. Having explained biodiversity loss as a factor driving disease rates, the authors make a plea for diagnosticians to become aware of the human health implications of environmental destruction.
Knowledge and prowess with infectious diseases for diagnosticians must be incorporated back into training with a reimagined lens crafted from the information we have gained by studying our environment, its destruction, and the ultimate resulting human infections. As loss of habitat, habitat fragmentation, and consequent biodiversity loss continue unabated, tools and skills will need to be in the hands of all diagnosticians if we hope to minimize the effect of these infections as they continually emerge [Granter 2016: 645].
This paper provides a particularly clear explanation of how biodiversity loss increases human infection risk.
The relationship between loss of biodiversity and human disease was first illustrated by Lyme disease. Its cause, the Borrelia burgdorferi bacterium, has the opportunity to encounter numerous vertebrate hosts – in one study estimated to be at least 125 species – in a diverse and healthy ecosystem. The potential hosts vary tremendously in their ability to harbor and transmit the bacteria, that is, their “reservoir competence.” Studies estimate the white-footed mouse infects more than 90% of ticks that complete their blood meal. While a few other hosts, such as eastern chipmunks and short-tailed shrews, are moderately competent, most tick hosts are marginally competent or dead-end hosts that are highly unlikely to transmit the infection. Since the white-footed mouse tends to thrive in impoverished ecosystems lacking biodiversity, infected ticks and, consequently, risk of human infection show a strong negative relationship with biodiversity. Because a diverse ecosystem with a range of vertebrate hosts – including many incompetent and dead-end hosts – “dilutes” the representation of the white-footed mouse and reduces human infection risk, this phenomenon has been termed the dilution effect [Granter 2016: 644].
“Because a diverse ecosystem with a range of vertebrate hosts – including many incompetent and dead-end hosts – “dilutes” the representation of the white-footed mouse and reduces human infection risk, this phenomenon has been termed the dilution effect” [Granter 2016: 644].
Conservation of biodiversity as a strategy for improving human health and wellbeing, Kilpatrick et al. 2017
This article very pragmatically addresses the question of whether biodiversity conservation could be an effective public health tool against infectious disease emergence and transmission.
Determining whether biodiversity conservation is an effective public health strategy requires answering four questions: (1) Is there a general, causal relationship between host biodiversity and disease risk? (2) If the link is causal and negative for most pathogens, does the increased diversity of pathogens with more diverse host communities result in net total increase or decrease in infectious disease burden? (3) Is the net benefit of biodiversity conservation greater than the net benefit of diversity-degrading processes (agricultural land-use change and wild animal harvesting)? (4) Are conservation interventions feasible and cost-effective compared to standard public health approaches (vaccines and treatments)?
Regarding the first question, experimental and observational research shows that increased biodiversity is associated with reduced disease burden.
Overall, the available data suggests that there is some correlational support in many zoonotic systems for a dilution effect, and that some species or species groups are more important than others in transmission [Kilpatrick 2017: 4].
The dilution effect hypothesis originated to explain the Lyme disease system. Greater numbers of hosts that are less “competent” (at spreading Lyme disease) – opossums, birds, raccoons and skunks – dilutes the transmission of Lyme bacteria to larval ticks by more competent hosts – white-footed mice, eastern chipmunks and shrews. Changes in community diversity affect, for example, host-vector encounter rates and host and vector abundances.
However,
much more research is needed to show that observed correlations are causal and to identify the mechanisms by which diversity is influencing disease risk [Kilpatrick 2017: 4].
The possibility of confounding factors in observational field studies is high because the same disturbances that change host diversity alters other aspects of transmission as well. For example, an ecosystem disturbance may, in addition to decreasing host diversity, also increase vector abundance, making it difficult to discern the proximate cause of increased disease rates. The authors note that the dilution effect may well cause decreased disease rates – more research is needed to determine this. But they caution that if the dilution effect turns out not to be the direct cause of decreased disease rates in any given pathogen system, then interventions to increase host diversity could be in vain with respect to that desired outcome.
Examples of potential conservation interventions to improve public health include preserving or restoring forest land, reintroducing top predators to control host populations, installing bat or owl boxes to increase predation of mosquitos (vectors) or rodents (hosts). Our still limited understanding of the mechanisms driving disease incidence patterns, however, make it difficult to predict outcomes for broad-scale land-use interventions, according to the authors. They argue instead that more targeted interventions aiming to reduce populations of key hosts in transmission may be more feasible public health tools than general land preservation. Even this, however, requires “deep understanding of both disease and population ecology.”
Further research to address this knowledge gap may be worth the investment, both for human wellbeing and for the planet. Exposure to nature has been shown to improve human mental and physical health and wellbeing, the authors note, regardless of biodiversity’s potential to reduce infectious disease. Furthermore,
If diverse communities can be shown to provide net benefits to human wellbeing, this could provide a powerful motivation for preserving Earth’s remaining biodiversity [Kilpatrick 2017: 7].
The nexus between forest fragmentation in Africa and Ebola virus disease outbreaks, Rulli et al. 2017
Ebola virus disease outbreaks in West and Central Africa have been linked to spillover from potential disease reservoirs such as bats, apes, and duikers (an antelope-like animal). Spillover has been thought to be related to population density, vegetation cover, and human activities such as hunting, poaching, and bushmeat consumption. In this study, forest data from satellites coupled with disease outbreak records identify a nexus between forest fragmentation and Ebola.
The researchers identified 11 sites of the first human infection of Ebola from a wild species having occurred since 2004. Changes in forest cover between the year 2000 (baseline year) and the years of first infection for each of these outbreaks were determined using high-resolution satellite data on tree cover. All 11 centers of infection were found to be located in forested areas where the rate of forest fragmentation was greater than the regional average. Similarly, forest fragmentation decreased with increasing distance from the centers of infection.
All 11 centers of infection were found to be located in forested areas where the rate of forest fragmentation was greater than the regional average.
The centers of first infection … tend to occur in areas where on the outbreak year the average degree of forest fragmentation (e.g., within a 25 km, 50 km or 100 km distance from the infection center) was significantly higher than in the rest of the region [Rulli 2017: 2].
Furthermore, eight of the 11 centers of infection were located in fragmentation “hotspots,” meaning within a cluster of highly fragmented forest areas.
Bats are the commonly accepted host to filoviruses such as Ebola and tend to increase in population in fragmented habitats. The geographic distribution of potential bat hosts was consistent with the distribution of the zoonotic niche of Ebola. A decline in the population of insectivorous bats and an increase in the frugivorous (fruit-eating) bat species as a result of forest fragmentation was observed. Reshaping forest boundaries, habitat disruption, and biodiversity loss may enhance the likelihood of zoonotic infection by increasing the abundance of a particular species and thus the prevalence of that species’ pathogens.
The fact that spillover tends to occur in hotspots of forest fragmentation rather than in clearcut areas suggests that chances of human interactions with host wildlife are higher in areas where human encroachment leaves forest fragments that provide habitat for reservoir species [Rulli 2017: 5].
Pressure on land and its products is increasingly pushing people into forested areas. Given the danger of zoonotic disease outbreak, any evaluation of the costs, risk, and benefits of forest loss and fragmentation should include global health considerations.
Habitat fragmentation, biodiversity loss and the risk of novel infectious disease emergence, Wilkinson 2018
Habitat loss reduces biodiversity, which leads to infectious disease emergence. The way a habitat is fragmented (how many patches it is divided into, how those patches are shaped, and what the distance is between them) further affects the extent of disease emergence. Both the number of divisions of habitat into smaller patches and the irregularity of patch shapes tend to increase habitat perimeter, which in turn increases contact between disease agents and humans.
The hazard is greatest in places with greater pre-existing biodiversity, where there is a greater diversity of microbial pathogens and associated hosts. There is a double risk of developing wilderness areas in these places because there are more pathogens to begin with, and the resulting biodiversity loss tends to amplify disease transmission.
Human encroachment into species-rich habitats may simultaneously decrease biodiversity and increase exposure of people to novel microbes [Wilkinson 2018: 1].
Integration of wildlife and environmental health into a One Health approach, Sleeman et al. 2019
This article introduces the concept of One Health, a public health framework adopted by the Centers for Disease Control in 2009, which recognizes the interdependence of humans, animals and our shared environment. The concept has gained traction as a way to address health problems arising from global environmental change.
Climate change, loss of biodiversity, habitat fragmentation and pollution, and subsequent degradation of natural environments threaten the range of ecosystem services that support all life on this planet [Sleeman 2019: 91].
…
It was the challenge of responding to these complex [environmental] problems that led to the emergence of the concept of One Health, which is defined by the Centers for Disease Control and Prevention (CDC) as the collaborative effort of multiple disciplines and sectors – working locally, nationally, regionally and globally – with the goal of achieving optimal health outcomes, recognizing the interconnection among people, animals, plants and our shared environment. This definition acknowledges that human, domestic animal and wildlife health are interconnected within the context of ecosystem/environmental health and provides a useful conceptual framework for the development of solutions to global health and environmental challenges. Given this interconnection, it follows that actions aimed primarily at improving the health of one part of the human-animal-environmental triad may have unanticipated consequences for the system as a whole if the harms they may cause to the other components are not considered. However, previous authors have noted that, despite the acknowledged interdependencies, few public or livestock health interventions include a consideration of biodiversity conservation or ecosystem/environmental health. Instead, health-promoting interventions focus largely on single-sector outcomes and, thus, may miss the opportunity to concurrently optimize outcomes in the other two sectors [Sleeman 2019: 92].
The authors suggest that despite its potential, the One Health approach does not as yet fully integrate wildlife and environmental health, instead favoring human health. Yet failure to optimize the health of all three realms can lead to unexpected and outcomes, ironically increasing risk to humans in some cases. Therefore, the authors propose the clarification of One Health values and goals, and integration of a systems approach and a harm reduction perspective into the One Health framework.
Systems biology provides methods to understand how interactions among [interrelated and interdependent] parts [livestock, humans and wildlife, for example] give rise to the function and behavior of that system [Sleeman 2019: 96].
A harm reduction perspective recognizes that solutions to complex problems require a broad societal response and that elimination of risk is not feasible for most issues. Consequently, this perspective promotes collaborative, multisectoral approaches whereby reducing harm, despite uncertainty regarding the outcome, is valued over inaction spurred by a desire for a perfect solution [Sleeman 2019: 94].
Emerging human infectious diseases and the links to global food production, Rohr et al. 2019
Increasing agricultural production to feed >11 billion people by 2100 raises several challenges for effectively managing infectious disease. Of many factors examined in this article linking agricultural expansion to infectious disease, one is conversion of natural habitat to cropland or rangeland. Land conversion increases contact between wild animals, livestock and humans.
As natural ecosystems are converted to crop land or range land, interactions among humans, and domesticated and wild animals, could increase. … These interactions are crucial because 77% of livestock pathogens are capable of infecting multiple host species, including wildlife and humans, and based on published estimates from the 2000s, over half of all recognized human pathogens are currently or originally zoonotic, as are 60–76% of recent emerging infectious disease events [Rohr 2019: 451].
“As natural ecosystems are converted to crop land or range land, interactions among humans, and domesticated and wild animals, could increase” [Rohr 2019: 451].
Land conversion pushes humans and livestock up against wilderness areas, increasing contact between species with previously little to no contact. The jumping of a pathogen to a new host species is called “spillover.”
Spillover appears to be a function of the frequency, duration and intimacy of interactions between a reservoir and novel host population. For example, influenza is believed to have jumped from horses to humans soon after domesticating horses and then made additional jumps to humans from other domesticated animals, such as poultry and swine [Rohr 2019: 451].
Furthermore, agricultural intensification tends to involve greater concentrations of a single variety of a single species, increasing the risk that any new disease will spread quickly in the population.
A central tenet of epidemiology is that the incidence of many infectious diseases should increase proportionally with host density because of increased contact rates and thus transmission among hosts. Hence, increasing human and livestock densities could cause increases in infectious diseases unless investments in disease prevention are sufficient to prevent these increases [Rohr 2019: 451].
Industrial-scale confined livestock production is
vulnerable to devastating losses of animals to disease. For instance, in just the last 25 years, an influenza A virus (H5N1) and a foot-and-mouth outbreak led to the destruction of more than 1.2 million chickens and 6 million livestock in China and Great Britain, respectively, and a ‘mad cow disease’ epizootic led to the slaughter of 11 million cattle worldwide [Rohr 2019: 449].
Increased agricultural production tends to be accompanied by new irrigation infrastructure and increased pesticide, fertilizer and antibiotic use, all of which increase infectious disease risk. Dams (often created for irrigation schemes) increase risk of mosquito-borne disease. Antibiotic overuse for livestock fosters resistance among pathogens that can also infect humans. Greater pesticide use leads to resistance among disease vectors such as mosquitoes to insecticides, while also weakening immune systems among exposed humans and wildlife hosts, increasing infection rates/severity. Nutrient enrichment caused by fertilizer can also contribute to the spread of infectious disease, for example, through mosquitos or snail vectors.
Finally, the urbanization and globalization associated with agricultural intensification/expansion elongates food supply chains, which increases movement of people and goods over borders, spreading food-born illness, flu and other infections.
In short,
These analyses revealed that agricultural drivers were associated with 25% of all diseases and nearly 50% of zoonotic diseases that emerged in humans since 1940. These values are even higher if we include the use of antimicrobial agents as an agricultural driver of human disease emergence, given that agricultural uses of antibiotics outpace medical uses in the developed world nearly nine to one [Rohr 2019: 451].
The authors recommend numerous measures for improving agricultural production while limiting infectious disease, including reducing antibiotic use for livestock, conserving biodiversity, improving and diversifying livestock and crop genetic material, investing in urban agriculture, social investments, and inter-disciplinary research and collaboration.
In a fitting juxtaposition, 2020 has brought us both the Covid-19 pandemic and the eve of the United Nations (UN) Decade of Ecosystem Restoration (2021-2030). As we have learned from infectious disease research, ecosystem degradation drives the emergence of novel human diseases that become pandemic. In this issue of the compendium we delve into research examining the connection between biodiversity loss and infectious disease, along with another set of articles on how best to restore the increasingly dangerous degraded lands that surround us. Perhaps a decade of restoring ecosystems as if our lives depend on it will deliver us to a gentler world come 2030.
Sacred forests/groves are not uncommon in India, especially in the biodiverse Western Ghats mountain range. These groves are community-protected patches of forest ranging in size from less than a hectare to several hundred hectares, and they are often believed to house gods [Ormsby & Bhagwat 2010]. A particular temple in the Western Ghats just outside the small town of Meenangadi in the state of Kerala owns 30 acres of forested hillside regarded as sacred.
In a clearing at the bottom of these hills, the people of the temple erected a shrine over a small stream flowing from their forest. Eventually, they built an elegant structure around the shrine with an opening in the roof to allow rain to wash onto the shrine. Praying involves touching the sacred stream water.
A few decades ago, however, in need of revenue, the temple cut a few acres at the uphill edge of their 30-acre plot and sold the wood. Following the loss of this portion of forest came a decline in the flow of the stream connecting the remaining sacred forest with the shrine. During the seasonally dry months of March and April, the stream had begun to run dry.
Concerned, temple officials went to the mayor in search of a solution to the drying of their stream. Town officials connected the temple with Kerala’s forestry department, whose mission includes increasing “the inflow of water to the reservoirs by improving the tree cover / forest cover over catchment areas,” “managing forests in such a way that it protects and enriches the social and cultural values of the state,” “protection and expansion of mangroves, sacred groves and other highly sensitive ecosystem,” and taking “Kerala to greater heights in the matter of biodiversity conservation,” among other goals [Kerala Forests and Wildlife Department website].
Given a mission such as this, the forest department knew what to do. They agreed to help by reforesting the clear-cut uphill edge of the sacred forest. Using only indigenous species, they planted some 89 different varieties of trees and shrubs, including medicinal varieties, on 1.6 hectares. This project was completed about five years ago.
On an afternoon in late January this year, as a Paradise Flycatcher darted in and out from the edge of the sacred grove to drink and bathe a few meters upstream of the shrine, a trained ear could hear another couple of dozen different bird species calling from the surrounding forest. Temple staff members said they have indeed seen more birds now that the cleared forest has been replanted. And the stream no longer runs dry, not even in March and April.
“It was a stink hole,” says Al Rizzo, the refuge manager of Prime Hook National Wildlife Refuge in Delaware Bay. Humans had messed with hydrology in an ill-conceived project aimed to convert salt marsh into a large open freshwater impoundment system to attract migrating waterfowl among others. Lines of dunes and tidal gates were constructed to barricade the inflow of salt water. However, severe storms, including Hurricane Sandy, tore open gaps in dune lines, inundating the re-engineered system with salt water, killing fresh-water marsh grass, and turning a healthy riparian forest into a wasteland of dead trees.
In order to reverse the damage of this unnatural disaster, government agencies and conservation groups used the Hurricane Sandy Disaster Relief Fund to embark on a $38 million attempt to restore 4000 acres of damaged wetland. Engineering with nature is how Rizzo and Bart Wilson, restoration project manager, describe their approach, in which they are taking cues from nature to create a more resilient ecosystem. The objective is to allow the system to adjust itself and to work based on normal coastal dynamics.
Relying on existing data, extensive hydrodynamic modeling was applied to find out what actually works. The refuge was found to have no elevation problem but rather a plumbing challenge. Work started with closing up the breaches by reconstructing the beach and dunes. The restored dunes are now 10ft high, allowing for overwash to dissipate storm and wave power. Sediments produced were cast onto the banks creating sand flats that are being colonized naturally by native grass. A neural network of channels was opened on historic waterways to let the tide flow back in and out. The result is a healthy tidal marsh with meandering channels, lush salt-tolerant grasses and mudflats that attract rich diversity of fish and birdlife. The Prime Hook is becoming a model for wetland restoration globally.
Demand for firewood in recent years led to the depletion of the mangrove forest in the Indonesia village of Paremas. For years the people’s occupations were agriculture and fishing. Depleted fish stock, poor irrigation and challenges associated with land ownership drove most of the men to work overseas in order to raise money to care for their loved ones, while some women went abroad to work as domestic servants. The women who stayed home have depended on their husbands’ remittances in addition to collecting fish and other sea life in pools.
However, about 10 years ago, the local government and environmental NGOs emphasized the significance of restoring the mangrove. With the help of the locals, everyone got to work replanting Paremas mangrove forest, which in turn now cushions the effect of tidal waves, limits coastal flooding, saves arable land from coastal erosion, reduces plastic and garbage deposit on the beach, and increases biodiversity. With the availability of crabs, vegetables and fruits, the women started making crab crackers, as well as cakes made from the flour of mangrove fruits, creating a source of income for the women to support their families. “There are many benefits now,” says Hanieti, a resident mother, “even the mosquitoes are gone.”
Sri Lanka is home to 82 lagoons and estuaries and is among the top five countries that will be impacted by climate risk. Thilakaratne De Silva, a 63-year old local fisherman, saw the Tsunami of December 2004 sweep off half his home village. He was among the first to join hands with other community members on a coastal natural buffer initiative of replanting the mangroves. In the aftermath of the tsunami, it became apparent that those who lived behind a thick buffer of mangrove forest were better shielded from the destructive waves.
The government and NGOs initiated the mangrove replanting, as well as implementing regulations aimed at curbing mangrove clearing, coral reef destruction and sand excavation. But after the Government and NGOs moved on, it’s been the local community’s engagement and consistency that is ensuring the success of a green belt along the south coast.
The critical goal now is to sustain the positive conservation effort already in place. Sustainable use enables gathering of edible mangrove varieties and collecting twigs and branches rather than cutting down trees and shrubs, according to Sarathchandra de Silva, an international agency worker involved in the replanting program.
Sugunawathi, a 51-year-old villager says that they are mindful of not cutting mangrove for fuelwood, though the burning efficiency of mangrove is preferred to forest wood, gas or kerosene. The frequency and extent to which communities access mangrove forest has much to do with poverty and lack of livelihood. However, growing tourism employs more rural men, creating livelihoods and boosting mangrove appreciation by the local community. A local NGO has sought to incentivize women because of their influence on children and the community on the need for the sustainable use of mangroves. The Galle success is now a model for replication, which inspires authorities to want to boost coastal buffer owning to the resilient nature of mangrove forest in climate risk, with an addition of 10,000 hectares to the existing 15,670 hectares already in place.
In continuation of the “blessed unrest” section of Compendium V3N1, the following sketches illustrate how people throughout the world are coming to recognise the enormous value of intact ecosystems, and are doing their part to protect and restore. Adopting Paul Hawken’s terminology and characterization of “blessed unrest” as a spontaneous, decentralized global social movement, we here present a diverse series of vignettes of everyday heroes. May such stories light the fire for new heroes to perpetually emerge in defense of all life on Earth.
Sri Lanka wields mangroves, its tsunami shield, against climate change, summarized from Mongabay News, September 2019
Sri Lanka is home to 82 lagoons and estuaries and is among the top five countries that will be impacted by climate risk. Thilakaratne De Silva, a 63-year old local fisherman, saw the Tsunami of December 2004 sweep off half his home village. He was among the first to join hands with other community members on a coastal natural buffer initiative of replanting the mangroves. In the aftermath of the tsunami, it became apparent that those who lived behind a thick buffer of mangrove forest were better shielded from the destructive waves.
The government and NGOs initiated the mangrove replanting, as well as implementing regulations aimed at curbing mangrove clearing, coral reef destruction and sand excavation. But after the Government and NGOs moved on, it’s been the local community’s engagement and consistency that is ensuring the success of a green belt along the south coast.
The critical goal now is to sustain the positive conservation effort already in place. Sustainable use enables gathering of edible mangrove varieties and collecting twigs and branches rather than cutting down trees and shrubs, according to Sarathchandra de Silva, an international agency worker involved in the replanting program.
Sugunawathi, a 51-year-old villager says that they are mindful of not cutting mangrove for fuelwood, though the burning efficiency of mangrove is preferred to forest wood, gas or kerosene. The frequency and extent to which communities access mangrove forest has much to do with poverty and lack of livelihood. However, growing tourism employs more rural men, creating livelihoods and boosting mangrove appreciation by the local community. A local NGO has sought to incentivize women because of their influence on children and the community on the need for the sustainable use of mangroves. The Galle success is now a model for replication, which inspires authorities to want to boost coastal buffer owning to the resilient nature of mangrove forest in climate risk, with an addition of 10,000 hectares to the existing 15,670 hectares already in place.
For one Indonesian village, mangrove restoration has been all upside, summarized from Mongabay News, September 2019
Demand for firewood in recent years led to the depletion of the mangrove forest in the Indonesia village of Paremas. For years the people’s occupations were agriculture and fishing. Depleted fish stock, poor irrigation and challenges associated with land ownership drove most of the men to work overseas in order to raise money to care for their loved ones, while some women went abroad to work as domestic servants. The women who stayed home have depended on their husbands’ remittances in addition to collecting fish and other sea life in pools.
However, about 10 years ago, the local government and environmental NGOs emphasized the significance of restoring the mangrove. With the help of the locals, everyone got to work replanting Paremas mangrove forest, which in turn now cushions the effect of tidal waves, limits coastal flooding, saves arable land from coastal erosion, reduces plastic and garbage deposit on the beach, and increases biodiversity. With the availability of crabs, vegetables and fruits, the women started making crab crackers, as well as cakes made from the flour of mangrove fruits, creating a source of income for the women to support their families. “There are many benefits now,” says Hanieti, a resident mother, “even the mosquitoes are gone.”
Coastal recovery: bringing a damaged wetland back to life, summarized from Yale Environment 360, May 2019
“It was a stink hole,” says Al Rizzo, the refuge manager of Prime Hook National Wildlife Refuge in Delaware Bay. Humans had messed with hydrology in an ill-conceived project aimed to convert salt marsh into a large open freshwater impoundment system to attract migrating waterfowl among others. Lines of dunes and tidal gates were constructed to barricade the inflow of salt water. However, severe storms, including Hurricane Sandy, tore open gaps in dune lines, inundating the re-engineered system with salt water, killing fresh-water marsh grass, and turning a healthy riparian forest into a wasteland of dead trees.
In order to reverse the damage of this unnatural disaster, government agencies and conservation groups used the Hurricane Sandy Disaster Relief Fund to embark on a $38 million attempt to restore 4000 acres of damaged wetland. Engineering with nature is how Rizzo and Bart Wilson, restoration project manager, describe their approach, in which they are taking cues from nature to create a more resilient ecosystem. The objective is to allow the system to adjust itself and to work based on normal coastal dynamics.
Relying on existing data, extensive hydrodynamic modeling was applied to find out what actually works. The refuge was found to have no elevation problem but rather a plumbing challenge. Work started with closing up the breaches by reconstructing the beach and dunes. The restored dunes are now 10ft high, allowing for overwash to dissipate storm and wave power. Sediments produced were cast onto the banks creating sand flats that are being colonized naturally by native grass. A neural network of channels was opened on historic waterways to let the tide flow back in and out. The result is a healthy tidal marsh with meandering channels, lush salt-tolerant grasses and mudflats that attract rich diversity of fish and birdlife. The Prime Hook is becoming a model for wetland restoration globally.
Indian temple restores sacred forest stream flow
Sacred forests/groves are not uncommon in India, especially in the biodiverse Western Ghats mountain range. These groves are community-protected patches of forest ranging in size from less than a hectare to several hundred hectares, and they are often believed to house gods [Ormsby & Bhagwat 2010]. A particular temple in the Western Ghats just outside the small town of Meenangadi in the state of Kerala owns 30 acres of forested hillside regarded as sacred.
In a clearing at the bottom of these hills, the people of the temple erected a shrine over a small stream flowing from their forest. Eventually, they built an elegant structure around the shrine with an opening in the roof to allow rain to wash onto the shrine. Praying involves touching the sacred stream water.
A few decades ago, however, in need of revenue, the temple cut a few acres at the uphill edge of their 30-acre plot and sold the wood. Following the loss of this portion of forest came a decline in the flow of the stream connecting the remaining sacred forest with the shrine. During the seasonally dry months of March and April, the stream had begun to run dry.
Concerned, temple officials went to the mayor in search of a solution to the drying of their stream. Town officials connected the temple with Kerala’s forestry department, whose mission includes increasing “the inflow of water to the reservoirs by improving the tree cover / forest cover over catchment areas,” “managing forests in such a way that it protects and enriches the social and cultural values of the state,” “protection and expansion of mangroves, sacred groves and other highly sensitive ecosystem,” and taking “Kerala to greater heights in the matter of biodiversity conservation,” among other goals [Kerala Forests and Wildlife Department website].
Given a mission such as this, the forest department knew what to do. They agreed to help by reforesting the clear-cut uphill edge of the sacred forest. Using only indigenous species, they planted some 89 different varieties of trees and shrubs, including medicinal varieties, on 1.6 hectares. This project was completed about five years ago.
On an afternoon in late January this year, as a Paradise Flycatcher darted in and out from the edge of the sacred grove to drink and bathe a few meters upstream of the shrine, a trained ear could hear another couple of dozen different bird species calling from the surrounding forest. Temple staff members said they have indeed seen more birds now that the cleared forest has been replanted. And the stream no longer runs dry, not even in March and April.
In climate policy and financing, the goals of adaptation (helping communities and ecosystems adapt to the effects of climate change) and mitigation (reducing carbon emissions and increasing carbon sinks) are often separate. This is because “adaptation and mitigation are driven by different interests and political economies, with distinct international donors and national institutions. These differences are reflected in the guidelines and requirements that climate change project developers have to follow” [Kongsager 2019: 279].
This present study, however, found that many climate change projects do or could address adaptation and mitigation jointly.
PDDs [Project Design Documents] integrating adaptation and mitigation shared several common features. They recognized ecosystems as providers of multiple services for both mitigation (carbon) and adaptation (watershed protection, forest products for livelihood diversification or safety nets, mangrove protection against storms and waves, microclimate regulation in agricultural fields) [Kongsager 2019: 278].
Ninety percent of mitigation projects mentioned adaptation goals while 30% of adaptation projects had mitigation goals. The authors speculate on the reason for this discrepancy:
There appears to be an intrinsic value in integrating adaptation into mitigation projects even without incentives provided by funders, because of reduced climatic risks and increased sustainability, as mentioned by a few project developers … By contrast, there is no clear rationale for a project developer to integrate mitigation into adaptation projects, beyond the perspective of receiving additional support from mitigation funding by selling carbon credits [Kongsager 2019: 279].
The authors recommend adjusting the institutional framework to encourage deeper, better coordinated integration of mitigation and adaptation goals into climate-related projects.
With a synergetic approach, AFOLU [agriculture, forests, and other land use] projects would be designed to combine adaptation and mitigation in a way that project components interact with each other to generate additional climate benefits compared to projects in which adaptation and mitigation are separated. Mainstreaming climate compatible development (i.e., adaptation, mitigation, and development) may avoid that projects respond to adaptation and mitigation urgencies separately. Scarce resources could be more efficiently spent, for instance, by not establishing separate institutions and processes to support adaptation and mitigation, and by avoiding conflicting policies, because a current major challenge in integrating adaptation and mitigation is the institutional complexity [Kongsager 2019: 279].
Forests currently cover a quarter of Earth’s terrestrial surface, although at least 82% of that remaining forest is degraded by human activity. While a handful of international accords rightly encourage forest conservation and reforestation to limit global warming, these agreements fail to prioritize protection specifically of intact forests, or forests that are free from human activities “known to cause physical changes in a forest that lead to declines in ecological function” [Watson 2018: 1].
The authors argue for the protection of all forests, including degraded ones, as well as reforestation, to stem global ecological collapse, while here they particularly emphasize the exceptional value of intact forests. They present these key arguments:
Carbon sequestration and storage. “Intact forests store more carbon than logged, degraded or planted forests in ecologically comparable locations. Industrial logging and conversion of forest to cropland causes heavy erosion and contributes to the loss of belowground carbon” [Watson 2018: 3].
Local climate. “Intact tropical forests are critical for rain generation because air that passes over these forests produces at least twice as much rain as air that passes over degraded or non-forest areas” [Watson 2018: 4]. By contrast, deforestation and forest degradation can increase the frequency of hot, dry days, leading to drought.
Biodiversity. Forested ecosystems support the majority of global terrestrial biodiversity, and “beyond outright forest clearance, forest degradation from logging is the most pervasive threat facing species inhabiting intact forests” [Watson 2018: 4]. “For example, a recent global analysis of nearly 20,000 vertebrate species showed that even minimal initial deforestation within an intact landscape had severe consequences for vertebrate biodiversity in a given region, emphasizing the special value of intact forests in minimizing extinction risk” [Watson 2018: 5].
Indigenous peoples. “Industrial-scale degradation of intact forest erodes the material basis for the livelihoods of indigenous forest peoples, depleting wildlife and other resources. It also renders traditional resource management strategies ineffective, and undermines the value of traditional knowledge and authority” [Watson 2018: 5], ultimately driving indigenous peoples off their land.
Human health. “Forested ecosystems are major sources of many medicinal compounds that supply millions of people with medicines worldwide” [Watson 2018: 6]. Forest degradation results in the decline or loss of medically relevant species, while also directly harming human health through increased wildfire severity and spread of disease.
Forest resilience. Forest degradation reduces the resilience of forests to climate change, leading to even greater ultimate loss of ecosystem function.
The authors warn that intact forests could soon disappear unless action is taken to protect them, which must necessarily start with greater recognition of their value compared to degraded forests.
There are still significant tracts of forest that are free from the damaging impacts of large-scale human activities. These intact forests typically provide more environmental and social values than forests that have been degraded by human activities. Despite these values, it is possible to envisage, within the current century, a world with few or no significant remaining intact forests. Humanity may be left with only degraded, damaged forests, in need of costly and sometimes unfeasible restoration, open to a cascade of further threats and lacking the resilience needed to weather the stresses of climate change. The practical tools required to address this challenge are generally well understood and include well-located and managed protected areas, indigenous territories that exemplify sound stewardship, regulatory controls and responsible behavior by logging, mining, and agricultural companies and consumers, and targeted restoration. Currently these tools are insufficiently applied, and inadequately supported by governance, policy and financial arrangements designed to incentivize conservation. Losing the remaining intact forests would exacerbate climate change effects through huge carbon emissions and the decline of a crucial, under-appreciated carbon sink. It would also result in the extinction of many species, harm communities worldwide by disrupting regional weather and hydrology, and devastate the cultures of many indigenous communities. Increased awareness of the scale and urgency of this problem is a necessary precondition for more effective conservation efforts across a wide range of spatial scales [Watson 2018: 8].
Despite their total area having increased by 10% since 1750, European forests have failed to achieve a net removal of CO2 from the atmosphere because of how they’ve been managed over that time. Eighty-five percent of Europe’s once largely unmanaged forest has been subjected to tree species conversion, wood extraction via thinning and harvesting, and litter raking, resulting in a large overall loss of carbon from biomass and soil.
Putting 417,000 km2 of previously unmanaged forest into production is estimated to have released 3.5 Pg of carbon to the atmosphere, because the carbon stock in living biomass, coarse woody debris, litter, and soil was simulated to be, respectively, 24, 43, 8, and 6% lower in managed forests compared with unmanaged forests [Naudts 2016: 598].
The sweeping change in Europe’s forest species from broad leaf trees to conifers represents a shift to a production-oriented approach to forestry undertaken to satisfy demands of a population that quadrupled between 1750 and 2010.
Whereas deforestation between 1750 and 1850 mainly replaced broadleaved forests with agricultural land, afforestation from 1850 onward was often with coniferous species. Broadleaved forests were also directly converted to coniferous forests, resulting in a total increase of 633,000 km2 in conifers at the expense of broadleaved forests (decreasing by 436,000 km2). For centuries, foresters have favored a handful of commercially successful tree species (Scots pine, Norway spruce, and beech) and, in doing so, are largely responsible for the current distribution of conifers and broadleaved species in Europe [Naudts 2016: 598].
The authors note that many other parts of the world have followed a similar path,
Wood extraction occurs in 64 to 72% of the 26.5 to 29.4 million km2 of global forest area, and substantial species changes have occurred in China (216,000 km2), Brazil (71,000 km2), Chile (24,000 km2), New Zealand (18,000 km2), and South Africa (17,000 km2) [Naudts 2016: 599].
This global trend has resulted in limiting the ability of many managed forests to sequester carbon, compared to their wild or better-managed counterparts.
Hence, any climate framework that includes land management as a pathway for climate mitigation should not only account for land-cover changes but also should equally address changes in forest management, because not all forest management contributes to climate change mitigation [Naudts 2016: 599].
The growth rate of trees – and thus their accumulation of carbon – increases continuously with tree size. Even though the leaves of smaller, younger trees are more efficient (more productive per unit area of leaf surface), larger trees have more total leaf surface area and thereby grow at a faster rate than their smaller counterparts. “For example, in our western USA old-growth forest plots, trees > 100cm in diameter comprised 6% of trees, yet contributed 33% of the annual forest mass growth” [Stephenson 2014: 92].
Since trees use atmospheric carbon to grow, the more they grow, the more carbon they sequester. “Thus, large, old trees do not act simply as senescent carbon reservoirs but actively fix large amounts of carbon compared to smaller trees” [Stephenson 2014: 90].
Phylogenetic[10] measures of diversity contain information on evolutionary divergences amongst species, thus representing the diversity of phylogenetically conserved traits related to resource use, acquisition and storage. Thereby, distantly related species are expected to respond differently to changing environmental conditions. These functional traits can be general traits related to the fast–slow growth rate spectrum, such as specific leaf area and wood density, and also physiological traits triggering plant responses to climatic fluctuations, such as flowering and leafing phenologies[11]. [Mazzochini 2019: 1431].
This study shows that (phylogenetic) plant diversity helps to stabilize ecosystem productivity – even at the landscape scale. Previous experiments have shown that plant diversity increases the stability of ecosystem productivity in small patches of vegetation where plant interaction boosts overall productivity, as does difference in response to environmental fluctuations among different species. The present study increases the scale of observation to the landscape level, and finds that biodiversity increases ecosystem stability due to varied or “asynchronous responses of distantly related species during environmental fluctuations” [Mazzochini 2019: 1431].
Our results expand by several orders of magnitude the spatial scale of evidence that high biodiversity strengthens the resistance of key ecosystem features to climatic fluctuations. Specifically, we show that the positive relationship between phylogenetic diversity and stability reported in local experiments can also be observed at larger spatial extents and grain sizes using available biodiversity databases and modelling techniques. As we expected, in the analyses at the landscape resolution, phylogenetic diversity correlates with vegetation productivity stability mainly due to a reduction in productivity variability across the years, and not by increasing average productivity, which was mostly driven by climatic variables [Mazzochini 2019: 1435].
This long-term experiment measured the difference in colonization and extinction rates of connected habitat fragments versus isolated fragments. The connected fragments were linked by a narrow (150m by 25m) strip of habitat. These habitat corridors increased the biodiversity of connected fragments by 14% after 18 years compared to their isolated counterparts.
In a large and well-replicated habitat fragmentation experiment, we find that annual colonization rates for 239 plant species in connected fragments are 5% higher and annual extinction rates 2% lower than in unconnected fragments. This has resulted in a steady, non-asymptotic increase in diversity, with nearly 14% more species in connected fragments after almost two decades. Our results show that the full biodiversity value of connectivity is much greater than previously estimated, cannot be effectively evaluated at short time scales, and can be maximized by connecting habitat sooner rather than later [Damschen 2019: 1479].
The authors note that 70% of the world’s forest area is within 1 km of an edge – meaning Earth’s forests are very fragmented. They stress that connecting habitat fragments is critical to the success of habitat and biodiversity conservation.
Conservation plans that ignore connectivity, such as plans that focus solely on habitat area, will leave unrealized the substantial, complementary, and persistent gains in biodiversity attributable specifically to landscape connectivity [Damschen 2019: 1480].
We found that wilderness areas act as a buffer against extinction risk. The global probability of species extinction in non-wilderness communities is over twice as high as that of species in wilderness communities. The buffering effect that wilderness has on extinction risk was found in every biogeographical realm, but was higher for realms with larger remaining extents of wilderness such as the Palaearctic[9] [Di Marco 2019: 26].
The remaining intact ecosystems of Earth—which are increasingly seen as essential for providing ecosystem services on which humanity relies and maintaining the bio-cultural connections of indigenous communities—have been neglected in efforts to conserve biodiversity. This is largely due to a belief that these areas are less vulnerable to threatening processes and less rich in threatened biodiversity, thereby having lower conservation value [Di Marco 2019: 585].
However,
These areas are important because they host highly unique biological communities and/or represent the majority of remaining natural habitats for biological communities that have suffered high levels of habitat loss elsewhere [Di Marco 2019: 585].
Indigenous people make up less than 5% of the global population, but their lands encompass 37% of the planet’s remaining natural lands and (partially overlapping with natural lands) 40% of Earth’s protected area, much of this in sparsely inhabited places. Like everyone, indigenous people have multiple interests (economic, political, cultural), which don’t necessarily always support conservation interests. However, “Indigenous Peoples often express deep spiritual and cultural ties to their land and contend that local ecosystems reflect millennia of their stewardship” [Garnett 2018: 369]. Indeed, “Countless Indigenous management institutions have already proven to be remarkably persistent and resilient, suggesting that such governance forms can shape sustainable human landscape relationships in many places” [Garnett 2018: 370].
Thus, the authors argue for indigenous voices to be prominent in land-use decision-making processes at global and local levels. “There is already good evidence that recognition of the practices, institutions and rights of Indigenous Peoples in global environmental governance is essential if we are to develop and achieve the next generation of global biodiversity targets” [Garnett 2018: 372].
In total, Indigenous Peoples influence land management across at least 28.1% of the land area.
About 7.8 million km2 (20.7%) of Indigenous Peoples’ lands are within protected areas, encompassing at least 40% of the global protected area with the proportion of Indigenous land in protected areas significantly higher than the proportion of other lands that are protected. The relationship between Indigenous Peoples and conserved areas varies in nature. While some protected areas (as defined by states and/or the International Union for Conservation of Nature (IUCN)) are under the governance of Indigenous Peoples themselves, others are governed by state authorities with varying degrees of respect for the presence of Indigenous Peoples. This respect ranges from collaborative governance where Indigenous Peoples are consulted on decisions, to de facto management and use of protected areas by Indigenous Peoples despite threats of eviction [Garnett 2018: 370].
Around half of the global terrestrial environment can be classified as human-dominated. Using this as a measure of human influence, we estimated that Indigenous Peoples’ lands account for 37% of all remaining natural lands across the Earth. A higher proportion (67%) of Indigenous Peoples’ lands was classified as natural compared with 44% of other lands. Even though no global data are available on other anthropogenic pressures such as grazing, burning, hunting or fishing, the drivers assessed by the Human Footprint (which range from roads, access, population density and different agricultural land use activity) are suitable surrogates. Consistent with this, most parts of the planet managed and/or owned by Indigenous Peoples have low intensity land uses: less than 3.8 million km2 (10.2%) of the world’s urban areas, villages and non-remote croplands are on Indigenous Peoples’ lands, whereas, in contrast, they encompass 24.9 million km2 (65.7%) of the remotest and least inhabited anthromes. Many of these remote Indigenous areas are nevertheless under pressure from intensive development [Garnett 2018: 370].
A spatial overview of the global importance of Indigenous lands for conservation, Garnett et al. 2018
Indigenous people make up less than 5% of the global population, but their lands encompass 37% of the planet’s remaining natural lands and (partially overlapping with natural lands) 40% of Earth’s protected area, much of this in sparsely inhabited places. Like everyone, indigenous people have multiple interests (economic, political, cultural), which don’t necessarily always support conservation interests. However, “Indigenous Peoples often express deep spiritual and cultural ties to their land and contend that local ecosystems reflect millennia of their stewardship” [Garnett 2018: 369]. Indeed, “Countless Indigenous management institutions have already proven to be remarkably persistent and resilient, suggesting that such governance forms can shape sustainable human landscape relationships in many places” [Garnett 2018: 370].
Thus, the authors argue for indigenous voices to be prominent in land-use decision-making processes at global and local levels. “There is already good evidence that recognition of the practices, institutions and rights of Indigenous Peoples in global environmental governance is essential if we are to develop and achieve the next generation of global biodiversity targets” [Garnett 2018: 372].
In total, Indigenous Peoples influence land management across at least 28.1% of the land area.
About 7.8 million km2 (20.7%) of Indigenous Peoples’ lands are within protected areas, encompassing at least 40% of the global protected area with the proportion of Indigenous land in protected areas significantly higher than the proportion of other lands that are protected. The relationship between Indigenous Peoples and conserved areas varies in nature. While some protected areas (as defined by states and/or the International Union for Conservation of Nature (IUCN)) are under the governance of Indigenous Peoples themselves, others are governed by state authorities with varying degrees of respect for the presence of Indigenous Peoples. This respect ranges from collaborative governance where Indigenous Peoples are consulted on decisions, to de facto management and use of protected areas by Indigenous Peoples despite threats of eviction [Garnett 2018: 370].
Around half of the global terrestrial environment can be classified as human-dominated. Using this as a measure of human influence, we estimated that Indigenous Peoples’ lands account for 37% of all remaining natural lands across the Earth. A higher proportion (67%) of Indigenous Peoples’ lands was classified as natural compared with 44% of other lands. Even though no global data are available on other anthropogenic pressures such as grazing, burning, hunting or fishing, the drivers assessed by the Human Footprint (which range from roads, access, population density and different agricultural land use activity) are suitable surrogates. Consistent with this, most parts of the planet managed and/or owned by Indigenous Peoples have low intensity land uses: less than 3.8 million km2 (10.2%) of the world’s urban areas, villages and non-remote croplands are on Indigenous Peoples’ lands, whereas, in contrast, they encompass 24.9 million km2 (65.7%) of the remotest and least inhabited anthromes. Many of these remote Indigenous areas are nevertheless under pressure from intensive development [Garnett 2018: 370].
Wilderness areas halve the extinction risk of terrestrial biodiversity, DiMarco et al. 2019
We found that wilderness areas act as a buffer against extinction risk. The global probability of species extinction in non-wilderness communities is over twice as high as that of species in wilderness communities. The buffering effect that wilderness has on extinction risk was found in every biogeographical realm, but was higher for realms with larger remaining extents of wilderness such as the Palaearctic[9] [Di Marco 2019: 26].
The remaining intact ecosystems of Earth—which are increasingly seen as essential for providing ecosystem services on which humanity relies and maintaining the bio-cultural connections of indigenous communities—have been neglected in efforts to conserve biodiversity. This is largely due to a belief that these areas are less vulnerable to threatening processes and less rich in threatened biodiversity, thereby having lower conservation value [Di Marco 2019: 585].
However,
These areas are important because they host highly unique biological communities and/or represent the majority of remaining natural habitats for biological communities that have suffered high levels of habitat loss elsewhere [Di Marco 2019: 585].
Ongoing accumulation of plant diversity through habitat connectivity in an 18-year experiment, Damschen et al. 2019
This long-term experiment measured the difference in colonization and extinction rates of connected habitat fragments versus isolated fragments. The connected fragments were linked by a narrow (150m by 25m) strip of habitat. These habitat corridors increased the biodiversity of connected fragments by 14% after 18 years compared to their isolated counterparts.
In a large and well-replicated habitat fragmentation experiment, we find that annual colonization rates for 239 plant species in connected fragments are 5% higher and annual extinction rates 2% lower than in unconnected fragments. This has resulted in a steady, non-asymptotic increase in diversity, with nearly 14% more species in connected fragments after almost two decades. Our results show that the full biodiversity value of connectivity is much greater than previously estimated, cannot be effectively evaluated at short time scales, and can be maximized by connecting habitat sooner rather than later [Damschen 2019: 1479].
The authors note that 70% of the world’s forest area is within 1 km of an edge – meaning Earth’s forests are very fragmented. They stress that connecting habitat fragments is critical to the success of habitat and biodiversity conservation.
Conservation plans that ignore connectivity, such as plans that focus solely on habitat area, will leave unrealized the substantial, complementary, and persistent gains in biodiversity attributable specifically to landscape connectivity [Damschen 2019: 1480].
Phylogenetic[10] measures of diversity contain information on evolutionary divergences amongst species, thus representing the diversity of phylogenetically conserved traits related to resource use, acquisition and storage. Thereby, distantly related species are expected to respond differently to changing environmental conditions. These functional traits can be general traits related to the fast–slow growth rate spectrum, such as specific leaf area and wood density, and also physiological traits triggering plant responses to climatic fluctuations, such as flowering and leafing phenologies[11]. [Mazzochini 2019: 1431].
This study shows that (phylogenetic) plant diversity helps to stabilize ecosystem productivity – even at the landscape scale. Previous experiments have shown that plant diversity increases the stability of ecosystem productivity in small patches of vegetation where plant interaction boosts overall productivity, as does difference in response to environmental fluctuations among different species. The present study increases the scale of observation to the landscape level, and finds that biodiversity increases ecosystem stability due to varied or “asynchronous responses of distantly related species during environmental fluctuations” [Mazzochini 2019: 1431].
Our results expand by several orders of magnitude the spatial scale of evidence that high biodiversity strengthens the resistance of key ecosystem features to climatic fluctuations. Specifically, we show that the positive relationship between phylogenetic diversity and stability reported in local experiments can also be observed at larger spatial extents and grain sizes using available biodiversity databases and modelling techniques. As we expected, in the analyses at the landscape resolution, phylogenetic diversity correlates with vegetation productivity stability mainly due to a reduction in productivity variability across the years, and not by increasing average productivity, which was mostly driven by climatic variables [Mazzochini 2019: 1435].
Rate of treecarbon accumulation increases continuously with tree size, Stephenson et al. 2014
The growth rate of trees – and thus their accumulation of carbon – increases continuously with tree size. Even though the leaves of smaller, younger trees are more efficient (more productive per unit area of leaf surface), larger trees have more total leaf surface area and thereby grow at a faster rate than their smaller counterparts. “For example, in our western USA old-growth forest plots, trees > 100cm in diameter comprised 6% of trees, yet contributed 33% of the annual forest mass growth” [Stephenson 2014: 92].
Since trees use atmospheric carbon to grow, the more they grow, the more carbon they sequester. “Thus, large, old trees do not act simply as senescent carbon reservoirs but actively fix large amounts of carbon compared to smaller trees” [Stephenson 2014: 90].
Europe’s forest management did not mitigate climate warming, Naudts et al. 2016
Despite their total area having increased by 10% since 1750, European forests have failed to achieve a net removal of CO2 from the atmosphere because of how they’ve been managed over that time. Eighty-five percent of Europe’s once largely unmanaged forest has been subjected to tree species conversion, wood extraction via thinning and harvesting, and litter raking, resulting in a large overall loss of carbon from biomass and soil.
Putting 417,000 km2 of previously unmanaged forest into production is estimated to have released 3.5 Pg of carbon to the atmosphere, because the carbon stock in living biomass, coarse woody debris, litter, and soil was simulated to be, respectively, 24, 43, 8, and 6% lower in managed forests compared with unmanaged forests [Naudts 2016: 598].
The sweeping change in Europe’s forest species from broad leaf trees to conifers represents a shift to a production-oriented approach to forestry undertaken to satisfy demands of a population that quadrupled between 1750 and 2010.
Whereas deforestation between 1750 and 1850 mainly replaced broadleaved forests with agricultural land, afforestation from 1850 onward was often with coniferous species. Broadleaved forests were also directly converted to coniferous forests, resulting in a total increase of 633,000 km2 in conifers at the expense of broadleaved forests (decreasing by 436,000 km2). For centuries, foresters have favored a handful of commercially successful tree species (Scots pine, Norway spruce, and beech) and, in doing so, are largely responsible for the current distribution of conifers and broadleaved species in Europe [Naudts 2016: 598].
The authors note that many other parts of the world have followed a similar path,
Wood extraction occurs in 64 to 72% of the 26.5 to 29.4 million km2 of global forest area, and substantial species changes have occurred in China (216,000 km2), Brazil (71,000 km2), Chile (24,000 km2), New Zealand (18,000 km2), and South Africa (17,000 km2) [Naudts 2016: 599].
This global trend has resulted in limiting the ability of many managed forests to sequester carbon, compared to their wild or better-managed counterparts.
Hence, any climate framework that includes land management as a pathway for climate mitigation should not only account for land-cover changes but also should equally address changes in forest management, because not all forest management contributes to climate change mitigation [Naudts 2016: 599].
The exceptional value of intact forest ecosystems, Watson et al. 2018
Forests currently cover a quarter of Earth’s terrestrial surface, although at least 82% of that remaining forest is degraded by human activity. While a handful of international accords rightly encourage forest conservation and reforestation to limit global warming, these agreements fail to prioritize protection specifically of intact forests, or forests that are free from human activities “known to cause physical changes in a forest that lead to declines in ecological function” [Watson 2018: 1].
The authors argue for the protection of all forests, including degraded ones, as well as reforestation, to stem global ecological collapse, while here they particularly emphasize the exceptional value of intact forests. They present these key arguments:
Carbon sequestration and storage. “Intact forests store more carbon than logged, degraded or planted forests in ecologically comparable locations. Industrial logging and conversion of forest to cropland causes heavy erosion and contributes to the loss of belowground carbon” [Watson 2018: 3].
Local climate. “Intact tropical forests are critical for rain generation because air that passes over these forests produces at least twice as much rain as air that passes over degraded or non-forest areas” [Watson 2018: 4]. By contrast, deforestation and forest degradation can increase the frequency of hot, dry days, leading to drought.
Biodiversity. Forested ecosystems support the majority of global terrestrial biodiversity, and “beyond outright forest clearance, forest degradation from logging is the most pervasive threat facing species inhabiting intact forests” [Watson 2018: 4]. “For example, a recent global analysis of nearly 20,000 vertebrate species showed that even minimal initial deforestation within an intact landscape had severe consequences for vertebrate biodiversity in a given region, emphasizing the special value of intact forests in minimizing extinction risk” [Watson 2018: 5].
Indigenous peoples. “Industrial-scale degradation of intact forest erodes the material basis for the livelihoods of indigenous forest peoples, depleting wildlife and other resources. It also renders traditional resource management strategies ineffective, and undermines the value of traditional knowledge and authority” [Watson 2018: 5], ultimately driving indigenous peoples off their land.
Human health. “Forested ecosystems are major sources of many medicinal compounds that supply millions of people with medicines worldwide” [Watson 2018: 6]. Forest degradation results in the decline or loss of medically relevant species, while also directly harming human health through increased wildfire severity and spread of disease.
Forest resilience. Forest degradation reduces the resilience of forests to climate change, leading to even greater ultimate loss of ecosystem function.
The authors warn that intact forests could soon disappear unless action is taken to protect them, which must necessarily start with greater recognition of their value compared to degraded forests.
There are still significant tracts of forest that are free from the damaging impacts of large-scale human activities. These intact forests typically provide more environmental and social values than forests that have been degraded by human activities. Despite these values, it is possible to envisage, within the current century, a world with few or no significant remaining intact forests. Humanity may be left with only degraded, damaged forests, in need of costly and sometimes unfeasible restoration, open to a cascade of further threats and lacking the resilience needed to weather the stresses of climate change. The practical tools required to address this challenge are generally well understood and include well-located and managed protected areas, indigenous territories that exemplify sound stewardship, regulatory controls and responsible behavior by logging, mining, and agricultural companies and consumers, and targeted restoration. Currently these tools are insufficiently applied, and inadequately supported by governance, policy and financial arrangements designed to incentivize conservation. Losing the remaining intact forests would exacerbate climate change effects through huge carbon emissions and the decline of a crucial, under-appreciated carbon sink. It would also result in the extinction of many species, harm communities worldwide by disrupting regional weather and hydrology, and devastate the cultures of many indigenous communities. Increased awareness of the scale and urgency of this problem is a necessary precondition for more effective conservation efforts across a wide range of spatial scales [Watson 2018: 8].
Addressing change mitigation and adaptation together: a global assessment of agriculture and forestry projects, Kongsager, Locatelli & Chazarin 2019
In climate policy and financing, the goals of adaptation (helping communities and ecosystems adapt to the effects of climate change) and mitigation (reducing carbon emissions and increasing carbon sinks) are often separate. This is because “adaptation and mitigation are driven by different interests and political economies, with distinct international donors and national institutions. These differences are reflected in the guidelines and requirements that climate change project developers have to follow” [Kongsager 2019: 279].
This present study, however, found that many climate change projects do or could address adaptation and mitigation jointly.
PDDs [Project Design Documents] integrating adaptation and mitigation shared several common features. They recognized ecosystems as providers of multiple services for both mitigation (carbon) and adaptation (watershed protection, forest products for livelihood diversification or safety nets, mangrove protection against storms and waves, microclimate regulation in agricultural fields) [Kongsager 2019: 278].
Ninety percent of mitigation projects mentioned adaptation goals while 30% of adaptation projects had mitigation goals. The authors speculate on the reason for this discrepancy:
There appears to be an intrinsic value in integrating adaptation into mitigation projects even without incentives provided by funders, because of reduced climatic risks and increased sustainability, as mentioned by a few project developers … By contrast, there is no clear rationale for a project developer to integrate mitigation into adaptation projects, beyond the perspective of receiving additional support from mitigation funding by selling carbon credits [Kongsager 2019: 279].
The authors recommend adjusting the institutional framework to encourage deeper, better coordinated integration of mitigation and adaptation goals into climate-related projects.
With a synergetic approach, AFOLU [agriculture, forests, and other land use] projects would be designed to combine adaptation and mitigation in a way that project components interact with each other to generate additional climate benefits compared to projects in which adaptation and mitigation are separated. Mainstreaming climate compatible development (i.e., adaptation, mitigation, and development) may avoid that projects respond to adaptation and mitigation urgencies separately. Scarce resources could be more efficiently spent, for instance, by not establishing separate institutions and processes to support adaptation and mitigation, and by avoiding conflicting policies, because a current major challenge in integrating adaptation and mitigation is the institutional complexity [Kongsager 2019: 279].
By Christopher A. Haines, Biodiversity for a Livable Climate
Christopher Haines is a seasoned architect licensed in both MA and NY who applies expertise in regenerative architectural design, healthy materials, preservation, renovation and specification writing to small commercial and urban projects. He has spoken for years at US and international forums as well as formally teaching sustainability and environmental management to undergraduate and graduate students. Christopher has been deeply involved in the Living Building Challenge and is certified as a Living Building Challenge consultant.
…
The year of 1979 was a critical one for the science of greenhouse gases and climate change. In April, the Jasons, a “mysterious coterie of elite scientists” [Rich 2019:15] published a report “The Long-Term Impact of Atmospheric Carbon Dioxide on Climate.” This report was subsequently reviewed by a National Academy of Science team commissioned by the Carter Administration and led by Jule Charney of Massachusetts Institute of Technology.
With the publication of the “Charney Report,” “Carbon Dioxide and Climate: A Scientific Assessment,” in July of 1979, the science of greenhouse gases as the sole cause of climate disruption was codified and canonized. Climate modeling was later made more sophisticated by the Intergovernmental Panel on Climate Change (IPCC), but the basic structure was not changed. We are thus the recipients of sophisticated modeling of a very complex system based on a single variable with secondary inputs.
We have spent the last 40 years so focused on the “pot lid” being the cause of the pot boiling over that we have failed to consider the possibility of turning down the stove. Climatologists all agree that the atmosphere is warmed by infrared rays radiated from the earth’s surface. Greenhouse gases are so named because they perform like a greenhouse, trapping that heat. But the ‘science’ seems to have overlooked where the heat comes from.
Sunshine is utilized differently depending on the interface it strikes. Living plant tissues promote photosynthesis and transpiration of water, causing cooling, while inanimate materials merely convert solar energy into heat. Everyone knows that walking barefoot on a summer day through a forest or across a meadow, desert or parking lot provide very different thermal experiences, particularly for your feet.
In walking across the desert and parking lot your feet experience sensible heat, the heat you can measure with a thermometer. In walking through the forest you experience latent heat, which is the heat required to convert water into vapor. Since considerable heat energy is spent converting liquid water into vapor (590 calories per gram), that heat does not increase the temperature. This is also the cooling mechanism of sweating. The evaporation of water increases humidity, and is thus experienced as sensible cooling. It’s worth noting that it takes only 80 calories to melt a gram of ice, which illustrates how powerful a cooling agent is the transfer of heat to water vapor (heat of vaporization). These experiences acknowledge different materials in different environments, generating very different temperatures.
Climatologists monitor reflectivity of land surfaces (“albedo”) as a guide to their thermal characteristics. Trees and forests have a reflectivity of between 5% and 20% for different species, and NASA uses an average of 14% for its Earth model. This turns out to be almost exactly the same as brick, concrete or urban areas overall. The question is what happens to the energy that is not reflected. Schneider and Sagan [2005] document energy balances for forests with 15% of energy reflected, 18% turned into heat, 1% turned into biomass and 66% used for transpiration. Yet for brick, concrete or urban areas, the roughly 85% of energy not reflected is turned wholly into heat.Thus albedo is an inaccurate description of the heat generating characteristics of a land surface.
Urban heat islands have been known since Luke Howard published his two-volume thesis on the Climate of London in 1818 and 1820. While we have focused on greenhouse gases that have risen 30.85% from 1800 to 2000, urban land area has increased 3345% and urban population has increased 3836% in the same timeframe.[8] And that is just the footprint. The solar interface – that is, the surface area of urban buildings that the sun strikes – could easily be ten or more times the footprint, even recognizing that some of the buildings’ surfaces remain in shade.
While the physics of urban heat islands were discovered in urban centers where the phenomenon is concentrated, what is critical to recognize is that the same physics apply everywhere. We define urban heat islands as urban centers that are warmer than non-urban areas some distance away. But the sun shines on non-urban areas as well, and any inanimate object – bare soil, a building, or a road – generates heat from insolation. Those non-urban areas are thus warmer than they would otherwise be, and we are underestimating the temperature rise of urban areas because we have no “zero zone” to measure against.
The destruction of biodiversity in any form that reduces leaf surface area, reduces the evapo-transpiration of water, the cooling it produces and the rain it promotes. It also reduces the biomass growth that increases sequestration. Thus biodiversity destruction directly causes a loss of cooling but is also the precursor to the generation of heat from bare ground, deforestation, aridification, open pit mining, mountaintop removal, roadways or the construction of buildings and urban centers. We must therefore recognize that the destruction of biodiversity causes double damage. Is it relevant to note that the IPCC has spent much of its correspondence in the last few decades admitting that it has underestimated the rate at which warming was occurring? Is this because it has failed to account for the rising temperature of the ‘stove’?
In other words, we have created what is effectively a global heat island, caused by solar energy generating sensible heat from the inanimate materials it strikes. Greenhouse gases exacerbate the problem by increasing the percentage of heat trapped, thus increasing the warming effect, but they do not cause the creation of sensible heat from solar energy.
The science of reducing urban heat islands has been known for decades, so we know what has to be done, although we surely need to dig deeper. With this perspective we open a world of opportunities to resolve our current predicament at the level of root causes.
Bare soil in agriculture does not produce as much heat as a roadway or a building, but there is a lot more of it. If all agriculture were converted to regenerative methods, such as practiced by Gabe Brown [2018], we might find that, globally, we have increased cooling and reduced heat generation sufficiently. Even short of a full transition to ecological/regenerative agriculture, a change in practices as simple as reducing fallow (bare) land in the Canadian prairie provinces produced significant improvements in temperature, rainfall, humidity and cooling [Vick 2016].
None of this diminishes the importance of reducing greenhouse gas emissions. However, it means that many other options to reverse climate disruption exist and several of the ones we have focused on may be less effective than others we have not yet considered, or even counterproductive. Perhaps some new balance between ecological regeneration and a reduction in greenhouse emissions will be found that would take us out of the danger zone.
The Heat Planet hypothesis provides a far more hopeful future where climate solutions are largely local and where impacts can be felt almost immediately.
This report, led by Ban Ki Moon (UN), Bill Gates (Bill & Melinda Gates Foundation) and Kristalina Georgieva (World Bank), calls on decision makers worldwide to facilitate coordinated action to help communities adapt to climate change. Importantly, the report makes the case for nature-based adaptation approaches, which inherently help mitigation efforts as well. Adaptation measures are much cheaper than recovery and rebuilding: every $1 invested in adaption yields $2-10 (or more by some estimates) in avoided losses and other economic benefits (such as improved crop yields), as well as social and environmental benefits.
Despite a clear global imperative for rapid adaptation planning and action to be taken at local, regional and national levels, action is desperately lagging. The report cites four reasons for inaction: (1) broad failure to internalize climate change risk in everyday decision making; (2) human tendency to prioritize short-term planning at the expense of long-term goals; (3) lack of cross-sector collaboration, which leads to fragmentation of responsibility; and (4) lack of power/voice among those most affected by climate change.
The report succinctly articulates the value of working with nature to adapt to climate change, while highlighting the extent to which this vital information is neglected.
We can already see the immense opportunity of using nature to increase societal resilience in landscapes ranging from uplands to the ocean. Restoring upland forests and watersheds could save water utilities in the world’s 534 largest cities an estimated $890 million each year and is critical for regulating water flows and managing the future’s more extreme floods. Meanwhile, lakes, marshes, and river floodplains both slow the release of floodwater and filter out sediment. The Netherlands has harnessed these capabilities with a Room for the River strategy that increases capacity of rivers and their floodplains to hold floodwaters, reducing damage and loss of life.
Ecosystem restoration also is a powerful tool for feeding the hungry, cooling sweltering cities, and protecting communities. One striking example is farmer-led reforestation in the Maradi and Zinder regions of Niger, which has boosted crop yields, improved soil fertility, and lifted communities out of poverty. Tree cover has soared ten-fold and the daily time spent gathering firewood—a task that mainly falls to women—has dropped from 3 hours to 30 minutes. For cities, an annual investment of $100 million in urban tree planting could create enough shade to cut average temperatures by 1°C for 77 million people around the world. Restoring the mangrove forests that offer protections from rising seas and storm surges is two to five times cheaper than building engineered structures like underwater breakwaters, while also storing carbon and improving water quality and local fisheries.
Yet despite the powerful case for working with nature to reduce climate risks, the world has barely begun to realize this potential. Few governments have adopted these approaches widely, even though many cite natural solutions in their NDCs. And only 3 percent of nearly 2,000 companies reported using natural ecosystems as part of their climate adaptation strategies. The barriers include lack of awareness of the critical role of natural assets in underpinning social and economic resilience and lack of accessible funds to invest in nature-based solutions. In addition, the piecemeal way adaptation is often planned and executed undervalues or ignores the many benefits of working with nature.
Humanity faces a stark choice: We can harness nature-based solutions to mitigate climate change and to better adapt—or we can continue with business as usual and lose the essential and myriad services nature provides [Global Commission on Climate Adaptation 2019: 31].
To encourage adoption of nature-based adaptation strategies, the report recommends three steps: (1) raise the level of understanding of the value of nature for climate adaptation; (2) embed nature-based solutions into adaptation planning and policy; and (3) increase investment into nature-based solutions. Indeed, it is precisely the aim of Biodiversity for a Livable Climate and its compendium series to elevate the level of understanding and appreciation for nature-based adaptation and mitigation solutions to the climate crisis.
Yet despite the powerful case for working with nature to reduce climate risks, the world has barely begun to realize this potential. Few governments have adopted these approaches widely, even though many cite natural solutions in their NDCs. And only 3 percent of nearly 2,000 companies reported using natural ecosystems as part of their climate adaptation strategies. The barriers include lack of awareness of the critical role of natural assets in underpinning social and economic resilience and lack of accessible funds to invest in nature-based solutions. In addition, the piecemeal way adaptation is often planned and executed undervalues or ignores the many benefits of working with nature.
Humanity faces a stark choice: We can harness nature-based solutions to mitigate climate change and to better adapt—or we can continue with business as usual and lose the essential and myriad services nature provides [Global Commission on Climate Adaptation 2019: 31].
The construction and operation of buildings and houses is a major source of pollution and ecosystem destruction around the world. In light of this, the Living Building Challenge invites people to reimagine the built environment as a source of social and ecological regeneration.
Nothing less than a sea change in building, infrastructure and community design is required. Indeed, this focus needs to be the great work of our generation. We must remake our cities, towns, neighborhoods, homes and offices, and all the spaces and infrastructure in between. This is part of the necessary process of reinventing our relationship with the natural world and each other—reestablishing ourselves as not separate from, but part of nature, “because the living environment is what really sustains us” (E.O. Wilson) [International Living Future Institute 2019: 8].
To that end the Living Building Challenge invites us to collaborate in building houses and buildings – or adapting existing ones – to have a positive, rather than simply less-negative, impact on the social and ecological systems where they are situated. The initiative runs educational and certification programs with several high standards, including, for example:
Projects must be observant of and responsive to the local ecological and social context of the sites, and onsite landscaping must seek to emulate local ecosystem function.
Access to locally grown food should be assured through onsite production and/or connection to local farms.
The site must ensure adequate habitat for local species.
Living Building Challenge designers must find ways to encourage pedestrian, bike and public transport options, while discouraging individual car travel.
Water should be harvested and wastewater treated onsite using living or natural/non-chemical systems.
Buildings/houses should supply their own energy on site (not through combustion), monitor their energy use, and minimize use through conservation.
Construction materials should be salvaged or sustainably and transparently sourced, and non-toxic.
In the interest of human wellbeing and social equity designs should allow natural light, beauty and comfort in the interior of the building, while the exterior must be accessible and welcoming to all members of the public, regardless of socioeconomic status.
Noting that 40% of carbon emissions in the US come from the construction (including sourcing of materials) and operation (heating, cooling, lighting) of buildings and houses, the AIA pledges to achieve zero-carbon construction and operation of all new buildings, and retrofitting of existing buildings to reduce their energy use and increase their resilience to severe weather. They will achieve these goals through education, policy advocacy, calling for zero-carbon building codes, and advocating for the reuse of historic buildings rather than new construction.
Because of the multiple threats and uncertainties of a changing climate, protecting coastal areas simply by building new seawalls (or some other such inflexible, single-tactic approach) is unlikely to be the most effective option. Instead, combined coastal adaptation strategies to allow for a dynamic response to multiple stressors are increasingly preferred. Climate scientists and coastal managers are mainstreaming inclusion of climate change into an Integrated Coastal Zone Management framework, aimed at promoting the activities of the different coastal sectors by coordinating government agencies and private participation.
Contrary to a “regret-risking option,” a no- or low-regret option is adopted to generate a net social benefit irrespective of the future outcome of climate change. Revamping early warning systems, preventing land reclamation, improving housing and transportation, capacity development in education, poverty reduction, and efforts to build resilient ecosystems are examples of a low- or no-regret options.
Traditional engineering, while sometimes protective of coastal communities, has undesired effects, such as eroding non-target, neighboring coastline and destroying adjacent ecosystems. By contrast, eco-engineering tools emphasize positive interactions among species that boost ecosystem productivity and stability, and therefore the strength of the ecosystem to withstand and buffer heavy storms, thus protecting coastal communities.
For example, sea-grasses planted with clams at their roots grow faster and in turn increase total fixed carbon. Oyster reefs attenuate up to 95% of wave height, control turbidity by removing algae, bacteria, and suspended organic matter, improve water quality through their filtration capacity, and enable seafood supply and thus job creation and recreation. Oyster reefs also support breeding ground for economically valued species, such as blue crab, red drums, flounder and spotted sea trout.
In mangroves, transplants planted in close proximity rather than the traditional spread pattern allows for a shared benefit of positive interaction that enhances plant growth and biodiversity. Restored mangrove ecosystems alleviate the impact of moderate tsunami waves, while the roots trap sediment and elevate the land surface, allowing for adaptation to sea-level rise. Intact mangrove also provides local employment as well as breeding grounds for fish.
Marshes dampen wave actions and reduce shoreline erosion, increase fish production, and are compatible with levee designs on the marshes’ landward edges that are nature-friendly. In the Netherlands, for instance, levees built to prevent flooding during storms were covered with thick grass to maintain their integrity, while the seaward marshes reduce the levees’ exposure to wave action; grasses were then grazed by sheep to provide milk and meat for consumption.
The synergy of ecology and engineering is key to addressing uncertainties related to climate-induced stressors. The combination of traditional and eco-engineering approaches coupled with the evaluation to measure the effectiveness of eco-engineered structures facilitate better decision making and prioritization of options.
While the majority of people on Earth live in cities, the majority (60%) of the world’s largest cities are located within 100 kilometers of a coast. The pollution and urban infrastructure (such as marinas, sea walls, or oil/gas platforms) emanating from cities greatly stresses coastal marine habitats. Coastal infrastructure tends to be vertical and smooth, offering little or nothing in the way of habitat niches or physical protection for various marine organisms. An eco-engineering approach to improve habitat quality and increase biodiversity is the addition of textural features, such as ledges, small holes, basins or crevices to the hard surfaces of urban marine infrastructure.
As predicted, overall microhabitat-enhancing interventions had a positive effect on the abundance and number of species across the studies. Nevertheless, the magnitude of their effects varied considerably, from zero to highly positive according to the type of intervention, the target taxa, and tidal elevation [Strain 2017: 434].
In the intertidal[5], interventions that provided moisture and shade had the greatest effect on the richness of sessile[6] and mobile organisms, while water-retaining features had the greatest effect on the richness of fish. In contrast, in the subtidal[7], small-scale depressions which provide refuge to new recruits from predators and other environmental stressors such as waves, had higher abundances of sessile organisms while elevated structures had higher numbers and abundances of fish. The taxa that responded most positively to eco-engineering in the intertidal were those whose body size most closely matched the dimensions of the resulting intervention [Strain 2017: 426].
What happened in New Orleans [during Hurricane Katrina], while a terrible “natural” disaster, was also the cumulative result of excessive and inappropriate management of the Mississippi River and delta, inadequate emergency preparation, a failure to act in time on plans to restore the wetlands and storm protection levees, and the expansion of the city into increasingly vulnerable areas [Costanza, Mitsch & Day 2006: 467].
Mismanagement here refers to damming, leveeing and canal dredging of the Mississippi River Delta, resulting in a significant loss of wetlands and the erosion of barrier islands over the past 100-plus years. Coastal marshes and barrier islands depend on regular inputs of sediments deposited by the river, which has been isolated from the delta plain and unable to thus nourish it. Two thirds of the river empties directly into the depths of the Gulf of Mexico, while one third empties into shallow waters, where it nourishes wetlands via the Atchafalaya, the river’s single remaining distributary (other distributaries having been closed off).
Damage from Hurricane Katrina was exacerbated by its prior loss of wetlands. Expansive coastal wetlands protect coastal communities from hurricanes by “decreasing the area of open water (fetch) for wind to form waves, increasing drag on water motion and hence the amplitude of a storm surge, reducing direct wind effect on the water surface, and directly absorbing wave energy” [Costanza, Mitsch & Day 2006: 468].
For the rebuilding of New Orleans after the hurricane, the authors recommended several core principles aimed at social and ecological resilience. Among their recommendations, they advise that areas of the city currently below sea level (by as much as 5 meters in some parts) not be rebuilt, but, rather, be restored to wetland. This would allow for temporary water storage within the city, water filtration, and biodiversity protection. They also suggest the reopening of distributaries and the controlled breaching of certain levees to allow the river to resume its ancient task of distributing sediment over a greater expanse of coastal marshes, allowing these marshes to gradually rise in step with sea level rise.
Given the dangerous, precipitous global decline in biodiversity, coupled with rapid urbanization, cities have a key role to play in protecting biodiversity. In fact, cities already do harbor a large share of biodiversity. This may be due to the fact that cities are often situated in places of large inherent biodiversity (along rivers, for example), and/or because of large numbers of introduced species and landscape heterogeneity in cities. Furthermore, surrounding agricultural areas are often simplified landscapes with limited biodiversity while many forests are degraded, and thus less biodiverse, due to timber harvest regimes, roads, etc. Thus, contrary to what might be assumed, rural areas are not necessarily more biodiverse than cities.
The author stresses the importance of managing cities to increase biodiversity. This process should begin with a city-wide tree inventory to identify tree species, locations and health. Management should focus on increasing biodiversity among street trees, and in parks, woodlots, abandoned lots, and back/front yards, while also fostering public awareness and appreciation for ecological principles. Planting efforts should prioritize native species, which are better adapted to local conditions, are non-invasive, and whose protection contributes to global biodiversity conservation. (While great numbers of introduced species may increase local biodiversity, it has a homogenizing effect on global biodiversity.) Furthermore, natural regeneration of parks and woodlots should be encouraged through less intensive management, whereby seeds of native (or at least non-invasive) species are allowed to germinate and establish where they fall, instead of being fastidiously mowed or weeded.
With 66% of the world’s population predicted to live in cities by 2050, the challenge of reconciling urban growth with biodiversity conservation demands attention.
Although the environment is altered by urbanization, there is potential for cities to support a great deal of biodiversity [Collas 2017: 1866].
This study shows that urban growth and biodiversity enhancement are compatible by increasing housing density (in order to reduce total surface area of development) while restoring ecosystems on remaining green space through woodlot plantings. The study was conducted in Cambridge, England, whose population is expected to grow by 22% between 2011 and 2031, and where “current green space supports relatively few trees” [Collas 2017: 54]. Green space could be maximized and restored to woodlots while additional high-density housing could accommodate the expected population growth.
Furthermore, only 2% of green space (i.e. ≥30 ha) is needed for conversion to woodlot to increase the native tree population size in Cambridge while also increasing high-density housing. This is compared to an alternative growth scenario, where new development is low-density and inhabitants are expected to plant trees in their relatively large yards, while no city-led green space ecosystem restoration occurs.
In conclusion, the authors offer this:
For other cities in the UK and across Europe, which have generally long been cleared of natural habitat, restoration in parallel with the expansion of higher density housing would appear to offer greatest scope for accommodating population growth at least cost to nature. This would require policy and economic incentives to directly link high-intensity human land-use to large-scale restoration [Collas 2017: 1871].
Like vegetative and light or reflective surfaces, water bodies have a cooling effect on cities, reducing the Urban Heat Island effect. The average temperature at the river in this study was 1C less than at a reference point elsewhere in the city. Furthermore, the form of the landscape on the banks of an urban river can either propagate (increase) or diminish the cooling effects of the river. This study found that vegetated river banks increased the cooling effect of the river by a difference of 2C compared to river banks covered in hard engineering materials (concrete/asphalt), while opening river banks (rather than enclosing them with buildings or walls) permitted significant cooling effects to be felt up to a distance of 30 meters from the river.
Overall, the results indicate that high levels of vegetation next to the river increase the cooling on the bank, that opening up the streets to the river increases the propagation of cooling and that the surface nature of the surrounding materials [e.g. vegetation versus concrete] can have a more significant effect on the air temperatures than the presence of the river [Hathaway & Sharples 2012: 20].
Overall, the results indicate that high levels of vegetation next to the river increase the cooling on the bank, that opening up the streets to the river increases the propagation of cooling and that the surface nature of the surrounding materials [e.g. vegetation versus concrete] can have a more significant effect on the air temperatures than the presence of the river [Hathaway & Sharples 2012: 20].
Urban heat islands are created when solar energy is absorbed by non-reflective, impervious, and often rather dark surfaces, such as asphalt, stone, metal, and concrete, which are ubiquitous in cities. Exacerbating this solar energy absorption effect are abundant amounts of heat released from vehicles, factories and air conditioners, for example, as well as pollutants trapped in the lower troposphere that slow down the cooling of rising air.
In New York City, where this study was conducted, the “summertime nocturnal heat island averages 7.2ºF (4ºC). This means that during the summer months the daily minimum temperature in the city is on average 7.2ºF (4ºC) warmer than surrounding suburban and rural areas” [Rosenzwieg 2006: 1]. The authors tested the cooling effects of tree plantings, living rooftops and high albedo (light colored) surfaces, and found that curbside tree plantings were the most effective form of cooling per unit area, followed by living rooftops. High albedo (light/white) surfaces were the least effective at cooling per unit area, but were the most effective overall “because 64% of New York City’s surface area could be redeveloped from dark, impervious surfaces to lighter high-albedo surfaces” [Rosenzwieg 2006: 3], whereas only 17% of the city’s surface could be planted with new street trees.
“The geometry, spacing and orientation of buildings and outdoor spaces” [Kleerekoper 2012: 30], as well as the prevalence of hard surfaces and reduced amount of vegetation, strongly modify the micro-climate of urban areas compared to rural surroundings. Characterized by an increase in temperature, a phenomenon referred to as urban heat island [UHI] effect has multiple causes. This includes, for example:
Absorption of short-wave radiation from the sun in low albedo (low-reflection/high-absorption) materials
Absorption and re-emission of longwave radiation by pollution
Heat released through combustion from traffic, heating and industries
Reduction of wind speed and obstruction of sky view by buildings, resulting in lowered heat loss from street “canyons”
Decreased evaporation due to a surfeit of impermeable surfaces
An increase in global temperature combined with UHI may have serious health implications including death. The heatwave of 2006 resulted in about a thousand heat-related deaths in the Netherlands and was rated fifth-worst natural disaster of that year. Cities can reduce the UHI effect with adaptive measures that combine vegetation, water, built form and material.
“Vegetation cools the environment actively by evapotranspiration and passively by shading surrounding surfaces that otherwise would have absorbed short-wave radiation” [Kleerekoper 2102: ]. Such methods include expanding urban forests/parks, street trees, private gardens, and green walls or roofs. “Vegetation has an average cooling effect of 1 – 4.7C that spreads 100 – 1,000m into an urban area, but is highly dependent on the amount of water the plant or tree has available” [Kleerekoper 2012: ]
Water cools by evaporation, or by transporting heat out of the city as does a river or stream. The cooling effect of water ranges from 1 – 30C to a distance of 30 – 35m, with stagnant water cooling the least and flowing and dispersed water (like a fountain) cooling the most. Water also cools through permeable pavement and water storage infrastructure that makes it available to trees for transpiration.
The built form of cities increases the UHI effect by reducing heat loss when tall buildings block the release of long-wave radiation back up toward the sky, while also blocking wind ventilation. While city form is hard to change, any new development can opt to reduce the height to width ratio of streets to allow better ventilation and heat loss. Slanted roofs also increase ventilation.
Lastly, the choice of building materials affects the UHI effect. Permeable materials facilitate evaporation and light/white (high albedo) materials reflect short-wave solar radiation, thus cooling the city. By contrast, “waterproof” and dark materials reduce evaporation and absorb short-wave radiation, thus contributing to the UHI effect. A simulation model for Sacramento, CA, showed a 1 – 4C drop in temperature from a city-wide increase in albedo (such as through white rooftops) from 25 to 40%.
Despite the existence of a substantial body of knowledge on the causes of and solutions to the UHI effect, the transfer of this knowledge to city planners is often lacking. Due to differences in aim, focus, and expression among the various actors in the city planning process, as well as the theoretical (rather than practical) nature of scientific studies discussing the UHI effect, communication about UHI-reduction design solutions can be a challenge. Furthermore, quantification is often lacking in terms of heat accumulation for a given area, maximum acceptable levels of heat, and the quantity of needed measures to reduce UHI (number of trees or square meters of green space, for example) .
However, certain cities like Stuttgart have developed spatial parameters in urban planning guidelines with respect to climate change. In California a cool-roof material with low thermal admittance has been introduced in the Building Energy Efficiency Standard regulation of the state. The city of Portland is creating a reference guide of pavement options for low-use traffic zones. The greening policy in Chicago and Edinburgh involves increasing the number of street trees, as well as species heterogeneity to ensure resistance to vegetal disease (given that species diversity limits pest infestation).
The study of urban ecology has grown rapidly over the past couple of decades as the planet becomes increasingly more urbanized. The field started as the study of ecology within the green spaces of cities, and has since evolved into a multidisciplinary approach to understanding the city itself as an ecosystem with interacting social, ecological and technical components.
A variety of social processes contribute to vulnerability to heat, including variation in social capital and legacies of disinvestment, which can affect vulnerability to heat waves. Furthermore, differences in intra-urban surface temperature can be as large or larger than urban-rural temperature differences, and a number of social-ecological-technical infrastructure interactions have been found to determine climate outcomes in cities. For instance, the dense distribution of tall buildings influences the spatial pattern of solar radiation intensity and duration and so influences air temperatures.[3] The highly heterogeneous distribution of vegetation in cities is a primary determinant of heat exposure, which is often greater for poor, elderly, and minority segments of the population, who are often less able to cope [McPhearson 2016: 9].
With an ultimate aim of fostering resilience among the world’s ever-growing cities, urban ecologists envision a transdisciplinary, participatory “ecology for cities” approach that integrates research and practice. Such collaboration could result in the beneficial integration of gray and green urban infrastructure.
Traditional risk-avoiding engineering designs for infrastructure design focus on hard, resistant elements such as increased-diameter sewage pipes for stormwater management or tanks to store sewage. In contrast, more flexible, diverse, and ecologically based elements include green infrastructure such as parks, permeable pavement, swales or retention basins, or agricultural and vacant land sites in urban areas. Urban infrastructure therefore mediates the relationships between human activities and ecosystem processes and may exacerbate or mitigate human impact depending on how it is developed [McPhearson 2016: 11].
Whereas just 10 percent of people lived in cities in 1900, now more than half the global population is urban and that proportion continues to grow. Cities occupy less than 3% of the Earth’s land surface, but generate 78% of global CO2 emissions and consume 76% of wood used for industrial purposes.
Urban dwellers depend on the productive and assimilative capacities of ecosystems well beyond their city boundaries — “ecological footprints” tens to hundreds of times the area occupied by a city — to produce the flows of energy, material goods, and nonmaterial services (including waste absorption) that sustain human well-being and quality of life [Grimm 2008: 756].
The social and environmental costs of building and servicing the world’s wealthiest cities since the colonial period to the present has been enormous:
Although exacerbated by recent globalization trends, centuries ago the demands of European consumers led to deforestation of colonial lands and, more recently, demand for beef from countries of the Western Hemisphere has transformed New World tropical rainforests into grazing land [Grimm 2008: 756].
Because cities so radically transform landscapes, creating new and less functional ecosystems in the process, they were “shunned” by ecologists during the 20th Century, “with the result that ecological knowledge contributed little to solving urban environmental problems” [Grimm 2008: 756]. However, even though cities contribute disproportionately to the current ecological crisis, they are by the same token increasingly seen as a necessary part of the solution, both in terms of mitigating their effects, and withstanding and adapting to severe weather.
The field of “urban ecology” seeks to better understand the processes and patterns of urban ecosystems, with an eye toward boosting the ecosystem services within and around cities upon which urban dwellers depend. An observable pattern is the cycling of resources through a city. “The concept of urban metabolism analogizes a city to an organism that takes in food and other required resources and releases wastes to the environment” [Grimm 2008: 757]. Unlike natural ecosystems, though, which constantly recycle resources, urban ecosystems notoriously recycle little to nothing, and are therefore reliant on fresh extraction for the provision of new resources, while letting waste products accumulate as pollution.
Through an urban ecology lens, this discrepancy between natural and urban resource metabolism is duly noted and practical solutions proposed:
Cities are hot spots of accumulation of N [nitrogen], P [phosphorus], and metals and, consequently, harbor a pool of material resources. Could high-nutrient, treated wastewater substitute for commercial N fertilizers to supply crops and lawns with nitrogen, for example? [Grimm 2008: 757]
Similarly,
A small (but growing) proportion of the copper extracted globally is recycled, yet increasing the reuse and recycling of copper and other metals would do much to stem the rapid rise in demand from sources increasingly difficult to extract. Such reuse also would alleviate problems of metal accumulation in soils [Grimm 2008: 757].
Another tendency of urban ecosystemsis to generate surplus heat, creating an urban heat island (UHI), due to reduced vegetation cover (thus, reduced cooling effects of evapotranspiration) and increased surface area absorbing solar energy (buildings, roads, etc.). This UHI effect in turn increases the use of air conditioning by 3-8% in the US, the additional energy use for which represents a positive feedback, which increases global warming. By contrast, increasing vegetation cover in cities reduces the UHI effect, while also removing greenhouse gases from the atmosphere, thus representing a negative feedback, which reduces global warming.
Another area of analysis in urban ecology involves water management (including channelization of streams and sewers, for example). The design of urban water systems is typically devoid of the ecosystem service provided by the waterways that urban systems replace, making cities vulnerable to flooding, drought and excessive pollution.
Among the most important modifications that affect streams in urban areas is increased impervious cover, which changes hydrology and funnels accumulated pollutants from buildings, roadways, and parking lots into streams [Grimm 2008: 759].
Yet,
Successful, ecologically based designs of novel urban aquatic ecosystems are becoming more common and exemplify stream-floodplain protection, retrofitting of neighborhood stormwater flow paths, and use of low-impact stormwater/water capture systems as creative solutions to urban stormwater management [Grimm 2008: 759].
Global change and the ecology of cities, Grimm et al. 2008
Whereas just 10 percent of people lived in cities in 1900, now more than half the global population is urban and that proportion continues to grow. Cities occupy less than 3% of the Earth’s land surface, but generate 78% of global CO2 emissions and consume 76% of wood used for industrial purposes.
Urban dwellers depend on the productive and assimilative capacities of ecosystems well beyond their city boundaries — “ecological footprints” tens to hundreds of times the area occupied by a city — to produce the flows of energy, material goods, and nonmaterial services (including waste absorption) that sustain human well-being and quality of life [Grimm 2008: 756].
The social and environmental costs of building and servicing the world’s wealthiest cities since the colonial period to the present has been enormous:
Although exacerbated by recent globalization trends, centuries ago the demands of European consumers led to deforestation of colonial lands and, more recently, demand for beef from countries of the Western Hemisphere has transformed New World tropical rainforests into grazing land [Grimm 2008: 756].
Because cities so radically transform landscapes, creating new and less functional ecosystems in the process, they were “shunned” by ecologists during the 20th Century, “with the result that ecological knowledge contributed little to solving urban environmental problems” [Grimm 2008: 756]. However, even though cities contribute disproportionately to the current ecological crisis, they are by the same token increasingly seen as a necessary part of the solution, both in terms of mitigating their effects, and withstanding and adapting to severe weather.
The field of “urban ecology” seeks to better understand the processes and patterns of urban ecosystems, with an eye toward boosting the ecosystem services within and around cities upon which urban dwellers depend. An observable pattern is the cycling of resources through a city. “The concept of urban metabolism analogizes a city to an organism that takes in food and other required resources and releases wastes to the environment” [Grimm 2008: 757]. Unlike natural ecosystems, though, which constantly recycle resources, urban ecosystems notoriously recycle little to nothing, and are therefore reliant on fresh extraction for the provision of new resources, while letting waste products accumulate as pollution.
Through an urban ecology lens, this discrepancy between natural and urban resource metabolism is duly noted and practical solutions proposed:
Cities are hot spots of accumulation of N [nitrogen], P [phosphorus], and metals and, consequently, harbor a pool of material resources. Could high-nutrient, treated wastewater substitute for commercial N fertilizers to supply crops and lawns with nitrogen, for example? [Grimm 2008: 757]
Similarly,
A small (but growing) proportion of the copper extracted globally is recycled, yet increasing the reuse and recycling of copper and other metals would do much to stem the rapid rise in demand from sources increasingly difficult to extract. Such reuse also would alleviate problems of metal accumulation in soils [Grimm 2008: 757].
Another tendency of urban ecosystemsis to generate surplus heat, creating an urban heat island (UHI), due to reduced vegetation cover (thus, reduced cooling effects of evapotranspiration) and increased surface area absorbing solar energy (buildings, roads, etc.). This UHI effect in turn increases the use of air conditioning by 3-8% in the US, the additional energy use for which represents a positive feedback, which increases global warming. By contrast, increasing vegetation cover in cities reduces the UHI effect, while also removing greenhouse gases from the atmosphere, thus representing a negative feedback, which reduces global warming.
Another area of analysis in urban ecology involves water management (including channelization of streams and sewers, for example). The design of urban water systems is typically devoid of the ecosystem service provided by the waterways that urban systems replace, making cities vulnerable to flooding, drought and excessive pollution.
Among the most important modifications that affect streams in urban areas is increased impervious cover, which changes hydrology and funnels accumulated pollutants from buildings, roadways, and parking lots into streams [Grimm 2008: 759].
Yet,
Successful, ecologically based designs of novel urban aquatic ecosystems are becoming more common and exemplify stream-floodplain protection, retrofitting of neighborhood stormwater flow paths, and use of low-impact stormwater/water capture systems as creative solutions to urban stormwater management [Grimm 2008: 759].
Advancing urban ecology toward a science of cities, McPhearson et al. 2016
The study of urban ecology has grown rapidly over the past couple of decades as the planet becomes increasingly more urbanized. The field started as the study of ecology within the green spaces of cities, and has since evolved into a multidisciplinary approach to understanding the city itself as an ecosystem with interacting social, ecological and technical components.
A variety of social processes contribute to vulnerability to heat, including variation in social capital and legacies of disinvestment, which can affect vulnerability to heat waves. Furthermore, differences in intra-urban surface temperature can be as large or larger than urban-rural temperature differences, and a number of social-ecological-technical infrastructure interactions have been found to determine climate outcomes in cities. For instance, the dense distribution of tall buildings influences the spatial pattern of solar radiation intensity and duration and so influences air temperatures.[3] The highly heterogeneous distribution of vegetation in cities is a primary determinant of heat exposure, which is often greater for poor, elderly, and minority segments of the population, who are often less able to cope [McPhearson 2016: 9].
With an ultimate aim of fostering resilience among the world’s ever-growing cities, urban ecologists envision a transdisciplinary, participatory “ecology for cities” approach that integrates research and practice. Such collaboration could result in the beneficial integration of gray and green urban infrastructure.
Traditional risk-avoiding engineering designs for infrastructure design focus on hard, resistant elements such as increased-diameter sewage pipes for stormwater management or tanks to store sewage. In contrast, more flexible, diverse, and ecologically based elements include green infrastructure such as parks, permeable pavement, swales or retention basins, or agricultural and vacant land sites in urban areas. Urban infrastructure therefore mediates the relationships between human activities and ecosystem processes and may exacerbate or mitigate human impact depending on how it is developed [McPhearson 2016: 11].
How to make a city climate-proof, Kleerekoper, van Esch & Salcedo 2012
“The geometry, spacing and orientation of buildings and outdoor spaces” [Kleerekoper 2012: 30], as well as the prevalence of hard surfaces and reduced amount of vegetation, strongly modify the micro-climate of urban areas compared to rural surroundings. Characterized by an increase in temperature, a phenomenon referred to as urban heat island [UHI] effect has multiple causes. This includes, for example:
Absorption of short-wave radiation from the sun in low albedo (low-reflection/high-absorption) materials
Absorption and re-emission of longwave radiation by pollution
Heat released through combustion from traffic, heating and industries
Reduction of wind speed and obstruction of sky view by buildings, resulting in lowered heat loss from street “canyons”
Decreased evaporation due to a surfeit of impermeable surfaces
An increase in global temperature combined with UHI may have serious health implications including death. The heatwave of 2006 resulted in about a thousand heat-related deaths in the Netherlands and was rated fifth-worst natural disaster of that year. Cities can reduce the UHI effect with adaptive measures that combine vegetation, water, built form and material.
“Vegetation cools the environment actively by evapotranspiration and passively by shading surrounding surfaces that otherwise would have absorbed short-wave radiation” [Kleerekoper 2102: ]. Such methods include expanding urban forests/parks, street trees, private gardens, and green walls or roofs. “Vegetation has an average cooling effect of 1 – 4.7C that spreads 100 – 1,000m into an urban area, but is highly dependent on the amount of water the plant or tree has available” [Kleerekoper 2012: ]
Water cools by evaporation, or by transporting heat out of the city as does a river or stream. The cooling effect of water ranges from 1 – 30C to a distance of 30 – 35m, with stagnant water cooling the least and flowing and dispersed water (like a fountain) cooling the most. Water also cools through permeable pavement and water storage infrastructure that makes it available to trees for transpiration.
The built form of cities increases the UHI effect by reducing heat loss when tall buildings block the release of long-wave radiation back up toward the sky, while also blocking wind ventilation. While city form is hard to change, any new development can opt to reduce the height to width ratio of streets to allow better ventilation and heat loss. Slanted roofs also increase ventilation.
Lastly, the choice of building materials affects the UHI effect. Permeable materials facilitate evaporation and light/white (high albedo) materials reflect short-wave solar radiation, thus cooling the city. By contrast, “waterproof” and dark materials reduce evaporation and absorb short-wave radiation, thus contributing to the UHI effect. A simulation model for Sacramento, CA, showed a 1 – 4C drop in temperature from a city-wide increase in albedo (such as through white rooftops) from 25 to 40%.
Despite the existence of a substantial body of knowledge on the causes of and solutions to the UHI effect, the transfer of this knowledge to city planners is often lacking. Due to differences in aim, focus, and expression among the various actors in the city planning process, as well as the theoretical (rather than practical) nature of scientific studies discussing the UHI effect, communication about UHI-reduction design solutions can be a challenge. Furthermore, quantification is often lacking in terms of heat accumulation for a given area, maximum acceptable levels of heat, and the quantity of needed measures to reduce UHI (number of trees or square meters of green space, for example) .
However, certain cities like Stuttgart have developed spatial parameters in urban planning guidelines with respect to climate change. In California a cool-roof material with low thermal admittance has been introduced in the Building Energy Efficiency Standard regulation of the state. The city of Portland is creating a reference guide of pavement options for low-use traffic zones. The greening policy in Chicago and Edinburgh involves increasing the number of street trees, as well as species heterogeneity to ensure resistance to vegetal disease (given that species diversity limits pest infestation).
Mitigating New York City’s heat island with urban forestry, living roofs and light surfaces, Rosenzwieg et al. 2006
Urban heat islands are created when solar energy is absorbed by non-reflective, impervious, and often rather dark surfaces, such as asphalt, stone, metal, and concrete, which are ubiquitous in cities. Exacerbating this solar energy absorption effect are abundant amounts of heat released from vehicles, factories and air conditioners, for example, as well as pollutants trapped in the lower troposphere that slow down the cooling of rising air.
In New York City, where this study was conducted, the “summertime nocturnal heat island averages 7.2ºF (4ºC). This means that during the summer months the daily minimum temperature in the city is on average 7.2ºF (4ºC) warmer than surrounding suburban and rural areas” [Rosenzwieg 2006: 1]. The authors tested the cooling effects of tree plantings, living rooftops and high albedo (light colored) surfaces, and found that curbside tree plantings were the most effective form of cooling per unit area, followed by living rooftops. High albedo (light/white) surfaces were the least effective at cooling per unit area, but were the most effective overall “because 64% of New York City’s surface area could be redeveloped from dark, impervious surfaces to lighter high-albedo surfaces” [Rosenzwieg 2006: 3], whereas only 17% of the city’s surface could be planted with new street trees.
The interaction of rivers and urban form in mitigating the Urban Heat Island effect: a UK case study, Hathaway & Sharples 2012
Like vegetative and light or reflective surfaces, water bodies have a cooling effect on cities, reducing the Urban Heat Island effect. The average temperature at the river in this study was 1C less than at a reference point elsewhere in the city. Furthermore, the form of the landscape on the banks of an urban river can either propagate (increase) or diminish the cooling effects of the river. This study found that vegetated river banks increased the cooling effect of the river by a difference of 2C compared to river banks covered in hard engineering materials (concrete/asphalt), while opening river banks (rather than enclosing them with buildings or walls) permitted significant cooling effects to be felt up to a distance of 30 meters from the river.
Overall, the results indicate that high levels of vegetation next to the river increase the cooling on the bank, that opening up the streets to the river increases the propagation of cooling and that the surface nature of the surrounding materials [e.g. vegetation versus concrete] can have a more significant effect on the air temperatures than the presence of the river [Hathaway & Sharples 2012: 20].
Overall, the results indicate that high levels of vegetation next to the river increase the cooling on the bank, that opening up the streets to the river increases the propagation of cooling and that the surface nature of the surrounding materials [e.g. vegetation versus concrete] can have a more significant effect on the air temperatures than the presence of the river [Hathaway & Sharples 2012: 20].
Urban development, land sharing and land sparing: the importance of considering restoration, Collas et al. 2017
With 66% of the world’s population predicted to live in cities by 2050, the challenge of reconciling urban growth with biodiversity conservation demands attention.
Although the environment is altered by urbanization, there is potential for cities to support a great deal of biodiversity [Collas 2017: 1866].
This study shows that urban growth and biodiversity enhancement are compatible by increasing housing density (in order to reduce total surface area of development) while restoring ecosystems on remaining green space through woodlot plantings. The study was conducted in Cambridge, England, whose population is expected to grow by 22% between 2011 and 2031, and where “current green space supports relatively few trees” [Collas 2017: 54]. Green space could be maximized and restored to woodlots while additional high-density housing could accommodate the expected population growth.
Furthermore, only 2% of green space (i.e. ≥30 ha) is needed for conversion to woodlot to increase the native tree population size in Cambridge while also increasing high-density housing. This is compared to an alternative growth scenario, where new development is low-density and inhabitants are expected to plant trees in their relatively large yards, while no city-led green space ecosystem restoration occurs.
In conclusion, the authors offer this:
For other cities in the UK and across Europe, which have generally long been cleared of natural habitat, restoration in parallel with the expansion of higher density housing would appear to offer greatest scope for accommodating population growth at least cost to nature. This would require policy and economic incentives to directly link high-intensity human land-use to large-scale restoration [Collas 2017: 1871].
Promoting and preserving biodiversity in the urban forest, Alvey 2006
Given the dangerous, precipitous global decline in biodiversity, coupled with rapid urbanization, cities have a key role to play in protecting biodiversity. In fact, cities already do harbor a large share of biodiversity. This may be due to the fact that cities are often situated in places of large inherent biodiversity (along rivers, for example), and/or because of large numbers of introduced species and landscape heterogeneity in cities. Furthermore, surrounding agricultural areas are often simplified landscapes with limited biodiversity while many forests are degraded, and thus less biodiverse, due to timber harvest regimes, roads, etc. Thus, contrary to what might be assumed, rural areas are not necessarily more biodiverse than cities.
The author stresses the importance of managing cities to increase biodiversity. This process should begin with a city-wide tree inventory to identify tree species, locations and health. Management should focus on increasing biodiversity among street trees, and in parks, woodlots, abandoned lots, and back/front yards, while also fostering public awareness and appreciation for ecological principles. Planting efforts should prioritize native species, which are better adapted to local conditions, are non-invasive, and whose protection contributes to global biodiversity conservation. (While great numbers of introduced species may increase local biodiversity, it has a homogenizing effect on global biodiversity.) Furthermore, natural regeneration of parks and woodlots should be encouraged through less intensive management, whereby seeds of native (or at least non-invasive) species are allowed to germinate and establish where they fall, instead of being fastidiously mowed or weeded.
A new vision for New Orleans and the Mississippi delta: applying ecological economics and ecological engineering, Costanza, Mitsch & Day 2006
What happened in New Orleans [during Hurricane Katrina], while a terrible “natural” disaster, was also the cumulative result of excessive and inappropriate management of the Mississippi River and delta, inadequate emergency preparation, a failure to act in time on plans to restore the wetlands and storm protection levees, and the expansion of the city into increasingly vulnerable areas [Costanza, Mitsch & Day 2006: 467].
Mismanagement here refers to damming, leveeing and canal dredging of the Mississippi River Delta, resulting in a significant loss of wetlands and the erosion of barrier islands over the past 100-plus years. Coastal marshes and barrier islands depend on regular inputs of sediments deposited by the river, which has been isolated from the delta plain and unable to thus nourish it. Two thirds of the river empties directly into the depths of the Gulf of Mexico, while one third empties into shallow waters, where it nourishes wetlands via the Atchafalaya, the river’s single remaining distributary (other distributaries having been closed off).
Damage from Hurricane Katrina was exacerbated by its prior loss of wetlands. Expansive coastal wetlands protect coastal communities from hurricanes by “decreasing the area of open water (fetch) for wind to form waves, increasing drag on water motion and hence the amplitude of a storm surge, reducing direct wind effect on the water surface, and directly absorbing wave energy” [Costanza, Mitsch & Day 2006: 468].
For the rebuilding of New Orleans after the hurricane, the authors recommended several core principles aimed at social and ecological resilience. Among their recommendations, they advise that areas of the city currently below sea level (by as much as 5 meters in some parts) not be rebuilt, but, rather, be restored to wetland. This would allow for temporary water storage within the city, water filtration, and biodiversity protection. They also suggest the reopening of distributaries and the controlled breaching of certain levees to allow the river to resume its ancient task of distributing sediment over a greater expanse of coastal marshes, allowing these marshes to gradually rise in step with sea level rise.
Eco-engineering urban infrastructure for marine and coastal biodiversity: which interventions have the greatest ecological benefit? Strain et al. 2017
While the majority of people on Earth live in cities, the majority (60%) of the world’s largest cities are located within 100 kilometers of a coast. The pollution and urban infrastructure (such as marinas, sea walls, or oil/gas platforms) emanating from cities greatly stresses coastal marine habitats. Coastal infrastructure tends to be vertical and smooth, offering little or nothing in the way of habitat niches or physical protection for various marine organisms. An eco-engineering approach to improve habitat quality and increase biodiversity is the addition of textural features, such as ledges, small holes, basins or crevices to the hard surfaces of urban marine infrastructure.
As predicted, overall microhabitat-enhancing interventions had a positive effect on the abundance and number of species across the studies. Nevertheless, the magnitude of their effects varied considerably, from zero to highly positive according to the type of intervention, the target taxa, and tidal elevation [Strain 2017: 434].
In the intertidal[5], interventions that provided moisture and shade had the greatest effect on the richness of sessile[6] and mobile organisms, while water-retaining features had the greatest effect on the richness of fish. In contrast, in the subtidal[7], small-scale depressions which provide refuge to new recruits from predators and other environmental stressors such as waves, had higher abundances of sessile organisms while elevated structures had higher numbers and abundances of fish. The taxa that responded most positively to eco-engineering in the intertidal were those whose body size most closely matched the dimensions of the resulting intervention [Strain 2017: 426].
Coastal adaptation with ecological engineering, Cheong et al. 2013
Because of the multiple threats and uncertainties of a changing climate, protecting coastal areas simply by building new seawalls (or some other such inflexible, single-tactic approach) is unlikely to be the most effective option. Instead, combined coastal adaptation strategies to allow for a dynamic response to multiple stressors are increasingly preferred. Climate scientists and coastal managers are mainstreaming inclusion of climate change into an Integrated Coastal Zone Management framework, aimed at promoting the activities of the different coastal sectors by coordinating government agencies and private participation.
Contrary to a “regret-risking option,” a no- or low-regret option is adopted to generate a net social benefit irrespective of the future outcome of climate change. Revamping early warning systems, preventing land reclamation, improving housing and transportation, capacity development in education, poverty reduction, and efforts to build resilient ecosystems are examples of a low- or no-regret options.
Traditional engineering, while sometimes protective of coastal communities, has undesired effects, such as eroding non-target, neighboring coastline and destroying adjacent ecosystems. By contrast, eco-engineering tools emphasize positive interactions among species that boost ecosystem productivity and stability, and therefore the strength of the ecosystem to withstand and buffer heavy storms, thus protecting coastal communities.
For example, sea-grasses planted with clams at their roots grow faster and in turn increase total fixed carbon. Oyster reefs attenuate up to 95% of wave height, control turbidity by removing algae, bacteria, and suspended organic matter, improve water quality through their filtration capacity, and enable seafood supply and thus job creation and recreation. Oyster reefs also support breeding ground for economically valued species, such as blue crab, red drums, flounder and spotted sea trout.
In mangroves, transplants planted in close proximity rather than the traditional spread pattern allows for a shared benefit of positive interaction that enhances plant growth and biodiversity. Restored mangrove ecosystems alleviate the impact of moderate tsunami waves, while the roots trap sediment and elevate the land surface, allowing for adaptation to sea-level rise. Intact mangrove also provides local employment as well as breeding grounds for fish.
Marshes dampen wave actions and reduce shoreline erosion, increase fish production, and are compatible with levee designs on the marshes’ landward edges that are nature-friendly. In the Netherlands, for instance, levees built to prevent flooding during storms were covered with thick grass to maintain their integrity, while the seaward marshes reduce the levees’ exposure to wave action; grasses were then grazed by sheep to provide milk and meat for consumption.
The synergy of ecology and engineering is key to addressing uncertainties related to climate-induced stressors. The combination of traditional and eco-engineering approaches coupled with the evaluation to measure the effectiveness of eco-engineered structures facilitate better decision making and prioritization of options.
Where we stand: climate action, The American Institute of Architects (AIA) 2019b
Noting that 40% of carbon emissions in the US come from the construction (including sourcing of materials) and operation (heating, cooling, lighting) of buildings and houses, the AIA pledges to achieve zero-carbon construction and operation of all new buildings, and retrofitting of existing buildings to reduce their energy use and increase their resilience to severe weather. They will achieve these goals through education, policy advocacy, calling for zero-carbon building codes, and advocating for the reuse of historic buildings rather than new construction.
Living Building Challenge Standard, June 2019
The construction and operation of buildings and houses is a major source of pollution and ecosystem destruction around the world. In light of this, the Living Building Challenge invites people to reimagine the built environment as a source of social and ecological regeneration.
Nothing less than a sea change in building, infrastructure and community design is required. Indeed, this focus needs to be the great work of our generation. We must remake our cities, towns, neighborhoods, homes and offices, and all the spaces and infrastructure in between. This is part of the necessary process of reinventing our relationship with the natural world and each other—reestablishing ourselves as not separate from, but part of nature, “because the living environment is what really sustains us” (E.O. Wilson) [International Living Future Institute 2019: 8].
To that end the Living Building Challenge invites us to collaborate in building houses and buildings – or adapting existing ones – to have a positive, rather than simply less-negative, impact on the social and ecological systems where they are situated. The initiative runs educational and certification programs with several high standards, including, for example:
Projects must be observant of and responsive to the local ecological and social context of the sites, and onsite landscaping must seek to emulate local ecosystem function.
Access to locally grown food should be assured through onsite production and/or connection to local farms.
The site must ensure adequate habitat for local species.
Living Building Challenge designers must find ways to encourage pedestrian, bike and public transport options, while discouraging individual car travel.
Water should be harvested and wastewater treated onsite using living or natural/non-chemical systems.
Buildings/houses should supply their own energy on site (not through combustion), monitor their energy use, and minimize use through conservation.
Construction materials should be salvaged or sustainably and transparently sourced, and non-toxic.
In the interest of human wellbeing and social equity designs should allow natural light, beauty and comfort in the interior of the building, while the exterior must be accessible and welcoming to all members of the public, regardless of socioeconomic status.
Adapt now: a global call for leadership on climate resilience, Global Commission on Adaptation, September 2019
This report, led by Ban Ki Moon (UN), Bill Gates (Bill & Melinda Gates Foundation) and Kristalina Georgieva (World Bank), calls on decision makers worldwide to facilitate coordinated action to help communities adapt to climate change. Importantly, the report makes the case for nature-based adaptation approaches, which inherently help mitigation efforts as well. Adaptation measures are much cheaper than recovery and rebuilding: every $1 invested in adaption yields $2-10 (or more by some estimates) in avoided losses and other economic benefits (such as improved crop yields), as well as social and environmental benefits.
Despite a clear global imperative for rapid adaptation planning and action to be taken at local, regional and national levels, action is desperately lagging. The report cites four reasons for inaction: (1) broad failure to internalize climate change risk in everyday decision making; (2) human tendency to prioritize short-term planning at the expense of long-term goals; (3) lack of cross-sector collaboration, which leads to fragmentation of responsibility; and (4) lack of power/voice among those most affected by climate change.
The report succinctly articulates the value of working with nature to adapt to climate change, while highlighting the extent to which this vital information is neglected.
We can already see the immense opportunity of using nature to increase societal resilience in landscapes ranging from uplands to the ocean. Restoring upland forests and watersheds could save water utilities in the world’s 534 largest cities an estimated $890 million each year and is critical for regulating water flows and managing the future’s more extreme floods. Meanwhile, lakes, marshes, and river floodplains both slow the release of floodwater and filter out sediment. The Netherlands has harnessed these capabilities with a Room for the River strategy that increases capacity of rivers and their floodplains to hold floodwaters, reducing damage and loss of life.
Ecosystem restoration also is a powerful tool for feeding the hungry, cooling sweltering cities, and protecting communities. One striking example is farmer-led reforestation in the Maradi and Zinder regions of Niger, which has boosted crop yields, improved soil fertility, and lifted communities out of poverty. Tree cover has soared ten-fold and the daily time spent gathering firewood—a task that mainly falls to women—has dropped from 3 hours to 30 minutes. For cities, an annual investment of $100 million in urban tree planting could create enough shade to cut average temperatures by 1°C for 77 million people around the world. Restoring the mangrove forests that offer protections from rising seas and storm surges is two to five times cheaper than building engineered structures like underwater breakwaters, while also storing carbon and improving water quality and local fisheries.
Yet despite the powerful case for working with nature to reduce climate risks, the world has barely begun to realize this potential. Few governments have adopted these approaches widely, even though many cite natural solutions in their NDCs. And only 3 percent of nearly 2,000 companies reported using natural ecosystems as part of their climate adaptation strategies. The barriers include lack of awareness of the critical role of natural assets in underpinning social and economic resilience and lack of accessible funds to invest in nature-based solutions. In addition, the piecemeal way adaptation is often planned and executed undervalues or ignores the many benefits of working with nature.
Humanity faces a stark choice: We can harness nature-based solutions to mitigate climate change and to better adapt—or we can continue with business as usual and lose the essential and myriad services nature provides [Global Commission on Climate Adaptation 2019: 31].
To encourage adoption of nature-based adaptation strategies, the report recommends three steps: (1) raise the level of understanding of the value of nature for climate adaptation; (2) embed nature-based solutions into adaptation planning and policy; and (3) increase investment into nature-based solutions. Indeed, it is precisely the aim of Biodiversity for a Livable Climate and its compendium series to elevate the level of understanding and appreciation for nature-based adaptation and mitigation solutions to the climate crisis.
Yet despite the powerful case for working with nature to reduce climate risks, the world has barely begun to realize this potential. Few governments have adopted these approaches widely, even though many cite natural solutions in their NDCs. And only 3 percent of nearly 2,000 companies reported using natural ecosystems as part of their climate adaptation strategies. The barriers include lack of awareness of the critical role of natural assets in underpinning social and economic resilience and lack of accessible funds to invest in nature-based solutions. In addition, the piecemeal way adaptation is often planned and executed undervalues or ignores the many benefits of working with nature.
Humanity faces a stark choice: We can harness nature-based solutions to mitigate climate change and to better adapt—or we can continue with business as usual and lose the essential and myriad services nature provides [Global Commission on Climate Adaptation 2019: 31].
The industrialization that has built today’s splendid high-tech cities isolated us from the land and water sources of the materials fueling this progress. Our cities scarcely reveal that the oxygen we breathe, the food we eat, the purification of waters, and to some extent the bucolic weather patterns we have long relished have been gifts from the ensemble of living creatures on Earth interacting in the form of ecosystems. We urbanites have managed in this brief past couple of centuries – a small fraction of human history – to harbor an illusion of independence from nature.
With their hard, gray and angled surfaces, ravenous appetites, and continuous toxic waste streams, cities in general surrendered themselves as ecological sacrifice zones. Against a foe so powerful as the drive for industrial development, there was never much hope for nature’s cause in a city. Yet to those of us with the means, cities are treasure troves of excitement, intrigue, social interaction, pleasure, and comfort, and it has perhaps been worth the sacrifice.
Yet if the wholesale exchange of ecosystem services for techno-industrial services was largely affordable in the 20th century, the growing likelihood of suffering or dying from climate-change-induced extreme weather makes this bargain harder to swallow. We’ve reached a time of reckoning. Cities of the world are now on the front lines of increasingly severe hurricanes, floods, fires, heat waves, and sea-level rise, and are thus bearing witness to our inextricable ties to nature.
As home to more than half of humanity, though, cities are uniquely positioned to drive the transition needed to salvage civilization and life in general from real-time ecological collapse. As aggregators of consumption and waste production, cities have plenty of slack to tighten. As population centers, they have the most at stake. As weavers of diverse cultures bursting with talent and creativity, cities have the people power to discover effective solutions. And as localized political bodies, cities have the potential to be cohesive enough to make key policy changes that favor adaptation and resilience to climate change.
Many effective ways to enhance urban resilience are known. They are described in various schools of thought – ecological engineering, industrial ecology, green infrastructure, sponge cities, biophilia, reconciliation ecology, and urban ecology, for example, which all generally aim to bolster ecosystem services in and around human habitation. Methods involve restoring original ecosystems (such as wetlands, forest, prairie, floodplains, mangrove forests, river delta systems and oyster reefs) adjacent to cities, and creating ecologically functional green spaces within cities, such as vegetated roofs and walls, or wooded parks. The ecosystem services these features deliver are meant to replace or fortify industrial infrastructure serving similar purposes. Here are a few examples:
Forested watersheds purify water for downstream consumers. For example, New York City compensates upstream watershed communities for protecting riparian forests and reducing agricultural runoff. These watershed investments totalling $1.5 billion over 10 years pale in comparison to the alternative of building a $6-billion dollar filtration system requiring additional annual operating and maintenance costs [Postel & Thompson 2005]. Globally, “restoring upland forests and watersheds could save water utilities in the world’s 534 largest cities an estimated $890 million each year and is critical for regulating water flows and managing the future’s more extreme floods” [Global Commission on Adaptation 2019: 31].
Salt marshes and coastal wetlands trap sediment to allow for coastal elevation rise in the face of sea-level rise, while also lowering erosion rates, and reducing wave strength in storms. “Properties behind a marsh, on average, save 16% in flood losses every year compared to properties where marshes have been lost,” according to a study of a coastal community in New Jersey hit by Hurricane Sandy in 2012 [Narayan 2017: 9463]. Coastal wetlands can even work in concert with nature friendly, grass-covered levees further inland by protecting the levees from wave damage [Cheong 2013].
An oyster reef can reduce up to 95% of wave height; trap sediment allowing the shoreline to rise as sea level rises; protect shorelines from erosion; improve water quality through filtration; and support breeding ground for economically valued species, such as blue crab, red drums, flounder and spotted sea trout [Cheong 2013]. By contrast, sea-walls, which can also protect coastal cities from wave action, have the negative side effects of eroding neighboring shoreline and damaging surrounding ecosystems.
A single full-grown tree has the cooling power of 10 air conditioning units [Kleerekoper 2012], and a patch of woods in a city can make even a broiling summer day feel fairly pleasant. Wooded areas of Washington DC are 17F degrees cooler than areas of the city with no trees on a hot summer day [NOAA 2018]. “For cities, an annual investment of $100 million in urban tree planting could create enough shade to cut average temperatures by 1°C for 77 million people around the world” [Global Commission on Adaptation 2019: 31].
Green walls and green roofs cool buildings through shading, evapotranspiration and insulation (the soil layer of a green roof insulates the building), thus significantly reducing buildings’ energy demands. Green walls have been shown to lower adjacent air temperatures by 0.2C to 3C [Kleerekoper 2012, Cameron 2014].
These methods are immediately relevant as cities continue to grow: two thirds of humanity is projected to reside in cities by 2050 [Collas 2017]. Population growth and aging infrastructure require ongoing renovation and new construction. For example, the number of houses in the US grew by 1.2 million between 2017 and 2018 [US Census 2019]. Globally, an estimated 60% of the built environment will be new or replaced by 2050 [Ahern 2014].
Individual buildings and houses are not necessarily designed, however, with climate risk in mind. The American Institute of Architects has recently created a new standard that a building must be designed to withstand environmental conditions over its entire design life. That means that if an architect today takes on the design of a commercial building whose estimated life is 80 years, it must be designed for the expected climatic conditions of 2100. For example, if climate-projection maps show a likelihood of flooding in a particular location, then a new building site should be considered; if high winds are a projected risk then special attention should be paid to roof design. However, architects are not required to consider climate projections and in many cases rely solely on building codes, which are based on historical data sometimes decades old [The American Institute of Architects 2019].
In the rush to grow and in the absence of an ecologically informed planning and design framework, new construction often increases a city’s vulnerability to severe weather and puts people in harm’s way who might otherwise have been better protected. “In several coastal U.S. states, for example, the highest rates of home construction since 2010 have occurred in flood-prone areas” [Global Commission on Adaptation 2019: 39]. Tragically, “cities are building over floodplains, forests, and wetlands that could have absorbed stormwater or offered respite and precious water during heat waves and droughts” [Global Commission on Adaptation 2019: 39]. Globally, some cities lack even basic floodplain maps.
Tragically, “cities are building over floodplains, forests, and wetlands that could have absorbed stormwater or offered respite and precious water during heat waves and droughts” [Global Commission on Adaptation 2019: 39]. Globally, some cities lack even basic floodplain maps.
Why, when information grows daily about climate risk as well as effective methods to enhance resilience, is it too often not taken into account? The answer to this question certainly involves complex political, cultural and economic analyses. Yet more superficially, a barrier may simply be that ecology and climate are not well integrated into mainstream planning and engineering, and that various types of expertise are too often siloed, resulting in insights from urban ecology being “often overlooked in engineering, planning, and policy for any sort of urban future.” Likewise, “the technical and built aspects of cities are central to the very fabric of urban systems and, perhaps unintentionally, are often overlooked or ignored in social-ecological studies” [McPhearson 2016: 11].
Even climate-focused institutions tend to unnecessarily separate mitigation and adaptation objectives, rather than thinking holistically through how solutions could be multifunctional and mutually reinforcing. This current lack of integration results in suboptimal outcomes and scarce resources being inefficiently spent. By contrast,
With a synergetic approach, AFOLU [agriculture, forests, and other land use] projects would be designed to combine adaptation and mitigation in a way that project components interact with each other to generate additional climate benefits compared to projects in which adaptation and mitigation are separated. Mainstreaming climate compatible development (i.e., adaptation, mitigation, and development) may avoid that projects respond to adaptation and mitigation urgencies separately [Kongsager 2016: 279].
Similarly, urban ecologists argue for holistic and collaborative approaches to development in the face of climate breakdown. Multidisciplinary teams must work together
to facilitate understanding of how both green and grey infrastructure can be linked – and in the future, deeply integrated – to deal with urban challenges and meet the needs of urban residents [McPhearson 2016: 11].
In addition to greater inter-sectoral collaboration, a large body of generalized and transferable knowledge in urban ecology is still needed, given the relative newness of this field, for planners to more easily incorporate new perspectives into their work. Given the urgent need for ecologically informed development, however, Ahern et al. [2014] argue that urban planners could forgo future research results by taking an ‘adaptive design’ approach. This would involve working in multidisciplinary teams that “learn by doing,” and incorporate low-risk, experimental design features into urban development plans, monitor results, make adjustments, and share what they learn.
Professionals, planners and designers need to make decisions in response to approval and development schedules. However, the “traditional” professional timeframe mandates an“imperative to act” that relies on readily available, existing knowledge and established best practices – typically without the opportunity to conduct new research. This imperative tends to favor decisions based on existing knowledge, and to inhibit innovation. The challenge of providing ecosystem services for urban sustainability planning and design will rely on emerging urban planning and design theory and new knowledge in design and engineering. Transdisciplinarity, implying the co-production of knowledge by scientists, planning professionals and urban dwellers, is a key to realize the potential of this planning approach [Ahern 2014: 255].
Further stressing the urgency to mainstream ecological principles into development planning and engineering, Mitch and Jorgenson [2003: 372] state: “The idea of nature conservation is so important that it needs to become a goal of engineering, not just one of its possible outcomes.” These authors argue that any engineering plans that omit ecological considerations have no place in today’s ecologically devastated world.
“The idea of nature conservation is so important that it needs to become a goal of engineering, not just one of its possible outcomes” [Mitch and Jorgenson 2003: 372].
We build solid waste facilities and water pollution control systems and atmospheric emissions of the greenhouse gas methane result. We use industrial wastewater treatment methods to remove heavy metals from a factory and then must dispose of metal-rich sludge. We burn sludge and solid wastes and we create air pollution problems. We are moving materials around in a shell game—if it is not under one shell, it is under another [Mitsch & Jorgenson 2003: 374].
The Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services reiterates this observation of the shortcomings of strictly human-built infrastructure and technology to deal with grave environmental problems.
People have created substitutes for some other contributions of nature, but many of them are imperfect or financially prohibitive. For example, high-quality drinking water can be realized either through ecosystems that filter pollutants or through human-engineered water treatment facilities. Similarly, coastal flooding from storm surges can be reduced either by coastal mangroves or by dikes and seawalls. In both cases, however, built infrastructure can be extremely expensive, incur high future costs and fail to provide synergistic benefits such as nursery habitats for edible fish or recreational opportunities. More generally, human-made replacements often do not provide the full range of benefits provided by nature[Diaz 2019: 13].
A great irony, as Carl Sagan once observed, is that as deeply divided as humanity is along nationality, political and class lines, not to mention professional specialization, the earth’s climate and ecosystems are deeply intertwined. What happens in Las Vegas or any other city does not stay in that city, but ripples out over the region and the globe. Likewise, how we manage landscapes upstream of and around a city significantly affects how badly battered it will be by the next big storm passing over it.
What the articles summarized below illustrate is that working with nature to boost ecosystem services to cities is eminently practical. Not only are ecological climate-change adaptation approaches highly effective as a protective buffer against severe weather; they also deliver several co-benefits – not the least which is mitigating the city’s impact on the rest of the planet by creating desperately needed wildlife habitat (at least for insects, birds and other small animals) and sequestering greenhouse gases. Urban green space also improves human health and wellbeing by reducing air pollution and providing islands of beauty and tranquility.
What’s exciting about the urban ecology opportunity is that more than half of us are perfectly situated to help shift the tide toward more ecologically coherent cities. Through collective action in shared spaces like schoolyards, churchyards or parks, even people without land of their own can restore ecosystems. Proponents for a “citizen science” model of grassroots engagement in conservation efforts [Francis & Lorimer 2011] note that urban areas have key advantages: (1) a large population base potentially available for training, implementation, monitoring and knowledge sharing related to ecosystem restoration; (2) concentrations of university and professional scientists with expertise to offer in grassroots and collaborative projects; and (3) high visibility of ecological urban adaptation efforts to generate enthusiasm and raise public awareness.
Francis and Lorimer [2011] further observe, however, that “motivation for conservation remains a major obstacle” and that “the biodiversity that emerges [from urban ecology efforts] is likely to be in untidy, unexpected and non-traditional forms” [Francis & Lorimer 2011: 1435] and that the management of expectations is therefore important. Perhaps, though, as the stakes continue to rise and to become clearer and more alarming to more and more people, the knowledge base already established on how to work with nature to better protect cities from climate change will be just enough to kick-start a paradigm shift in urban planning and development.
More generally, human-made replacements often do not provide the full range of benefits provided by nature [Diaz 2019: 13].
Compilation of article summaries on adaptation and urban resilience
Global change and the ecology of cities, Grimm et al. 2008
Whereas just 10 percent of people lived in cities in 1900, now more than half the global population is urban and that proportion continues to grow. Cities occupy less than 3% of the Earth’s land surface, but generate 78% of global CO2 emissions and consume 76% of wood used for industrial purposes.
Urban dwellers depend on the productive and assimilative capacities of ecosystems well beyond their city boundaries — “ecological footprints” tens to hundreds of times the area occupied by a city — to produce the flows of energy, material goods, and nonmaterial services (including waste absorption) that sustain human well-being and quality of life [Grimm 2008: 756].
The social and environmental costs of building and servicing the world’s wealthiest cities since the colonial period to the present has been enormous:
Although exacerbated by recent globalization trends, centuries ago the demands of European consumers led to deforestation of colonial lands and, more recently, demand for beef from countries of the Western Hemisphere has transformed New World tropical rainforests into grazing land [Grimm 2008: 756].
Because cities so radically transform landscapes, creating new and less functional ecosystems in the process, they were “shunned” by ecologists during the 20th Century, “with the result that ecological knowledge contributed little to solving urban environmental problems” [Grimm 2008: 756]. However, even though cities contribute disproportionately to the current ecological crisis, they are by the same token increasingly seen as a necessary part of the solution, both in terms of mitigating their effects, and withstanding and adapting to severe weather.
The field of “urban ecology” seeks to better understand the processes and patterns of urban ecosystems, with an eye toward boosting the ecosystem services within and around cities upon which urban dwellers depend. An observable pattern is the cycling of resources through a city. “The concept of urban metabolism analogizes a city to an organism that takes in food and other required resources and releases wastes to the environment” [Grimm 2008: 757]. Unlike natural ecosystems, though, which constantly recycle resources, urban ecosystems notoriously recycle little to nothing, and are therefore reliant on fresh extraction for the provision of new resources, while letting waste products accumulate as pollution.
Through an urban ecology lens, this discrepancy between natural and urban resource metabolism is duly noted and practical solutions proposed:
Cities are hot spots of accumulation of N [nitrogen], P [phosphorus], and metals and, consequently, harbor a pool of material resources. Could high-nutrient, treated wastewater substitute for commercial N fertilizers to supply crops and lawns with nitrogen, for example? [Grimm 2008: 757]
Similarly,
A small (but growing) proportion of the copper extracted globally is recycled, yet increasing the reuse and recycling of copper and other metals would do much to stem the rapid rise in demand from sources increasingly difficult to extract. Such reuse also would alleviate problems of metal accumulation in soils [Grimm 2008: 757].
Another tendency of urban ecosystemsis to generate surplus heat, creating an urban heat island (UHI), due to reduced vegetation cover (thus, reduced cooling effects of evapotranspiration) and increased surface area absorbing solar energy (buildings, roads, etc.). This UHI effect in turn increases the use of air conditioning by 3-8% in the US, the additional energy use for which represents a positive feedback, which increases global warming. By contrast, increasing vegetation cover in cities reduces the UHI effect, while also removing greenhouse gases from the atmosphere, thus representing a negative feedback, which reduces global warming.
Another area of analysis in urban ecology involves water management (including channelization of streams and sewers, for example). The design of urban water systems is typically devoid of the ecosystem service provided by the waterways that urban systems replace, making cities vulnerable to flooding, drought and excessive pollution.
Among the most important modifications that affect streams in urban areas is increased impervious cover, which changes hydrology and funnels accumulated pollutants from buildings, roadways, and parking lots into streams [Grimm 2008: 759].
Yet,
Successful, ecologically based designs of novel urban aquatic ecosystems are becoming more common and exemplify stream-floodplain protection, retrofitting of neighborhood stormwater flow paths, and use of low-impact stormwater/water capture systems as creative solutions to urban stormwater management [Grimm 2008: 759].
Advancing urban ecology toward a science of cities, McPhearson et al. 2016
The study of urban ecology has grown rapidly over the past couple of decades as the planet becomes increasingly more urbanized. The field started as the study of ecology within the green spaces of cities, and has since evolved into a multidisciplinary approach to understanding the city itself as an ecosystem with interacting social, ecological and technical components.
A variety of social processes contribute to vulnerability to heat, including variation in social capital and legacies of disinvestment, which can affect vulnerability to heat waves. Furthermore, differences in intra-urban surface temperature can be as large or larger than urban-rural temperature differences, and a number of social-ecological-technical infrastructure interactions have been found to determine climate outcomes in cities. For instance, the dense distribution of tall buildings influences the spatial pattern of solar radiation intensity and duration and so influences air temperatures.[3] The highly heterogeneous distribution of vegetation in cities is a primary determinant of heat exposure, which is often greater for poor, elderly, and minority segments of the population, who are often less able to cope [McPhearson 2016: 9].
With an ultimate aim of fostering resilience among the world’s ever-growing cities, urban ecologists envision a transdisciplinary, participatory “ecology for cities” approach that integrates research and practice. Such collaboration could result in the beneficial integration of gray and green urban infrastructure.
Traditional risk-avoiding engineering designs for infrastructure design focus on hard, resistant elements such as increased-diameter sewage pipes for stormwater management or tanks to store sewage. In contrast, more flexible, diverse, and ecologically based elements include green infrastructure such as parks, permeable pavement, swales or retention basins, or agricultural and vacant land sites in urban areas. Urban infrastructure therefore mediates the relationships between human activities and ecosystem processes and may exacerbate or mitigate human impact depending on how it is developed [McPhearson 2016: 11].
How to make a city climate-proof, Kleerekoper, van Esch & Salcedo 2012
“The geometry, spacing and orientation of buildings and outdoor spaces” [Kleerekoper 2012: 30], as well as the prevalence of hard surfaces and reduced amount of vegetation, strongly modify the micro-climate of urban areas compared to rural surroundings. Characterized by an increase in temperature, a phenomenon referred to as urban heat island [UHI] effect has multiple causes. This includes, for example:
Absorption of short-wave radiation from the sun in low albedo (low-reflection/high-absorption) materials
Absorption and re-emission of longwave radiation by pollution
Heat released through combustion from traffic, heating and industries
Reduction of wind speed and obstruction of sky view by buildings, resulting in lowered heat loss from street “canyons”
Decreased evaporation due to a surfeit of impermeable surfaces
An increase in global temperature combined with UHI may have serious health implications including death. The heatwave of 2006 resulted in about a thousand heat-related deaths in the Netherlands and was rated fifth-worst natural disaster of that year. Cities can reduce the UHI effect with adaptive measures that combine vegetation, water, built form and material.
“Vegetation cools the environment actively by evapotranspiration and passively by shading surrounding surfaces that otherwise would have absorbed short-wave radiation” [Kleerekoper 2102: ]. Such methods include expanding urban forests/parks, street trees, private gardens, and green walls or roofs. “Vegetation has an average cooling effect of 1 – 4.7C that spreads 100 – 1,000m into an urban area, but is highly dependent on the amount of water the plant or tree has available” [Kleerekoper 2012: ]
Water cools by evaporation, or by transporting heat out of the city as does a river or stream. The cooling effect of water ranges from 1 – 30C to a distance of 30 – 35m, with stagnant water cooling the least and flowing and dispersed water (like a fountain) cooling the most. Water also cools through permeable pavement and water storage infrastructure that makes it available to trees for transpiration.
The built form of cities increases the UHI effect by reducing heat loss when tall buildings block the release of long-wave radiation back up toward the sky, while also blocking wind ventilation. While city form is hard to change, any new development can opt to reduce the height to width ratio of streets to allow better ventilation and heat loss. Slanted roofs also increase ventilation.
Lastly, the choice of building materials affects the UHI effect. Permeable materials facilitate evaporation and light/white (high albedo) materials reflect short-wave solar radiation, thus cooling the city. By contrast, “waterproof” and dark materials reduce evaporation and absorb short-wave radiation, thus contributing to the UHI effect. A simulation model for Sacramento, CA, showed a 1 – 4C drop in temperature from a city-wide increase in albedo (such as through white rooftops) from 25 to 40%.
Despite the existence of a substantial body of knowledge on the causes of and solutions to the UHI effect, the transfer of this knowledge to city planners is often lacking. Due to differences in aim, focus, and expression among the various actors in the city planning process, as well as the theoretical (rather than practical) nature of scientific studies discussing the UHI effect, communication about UHI-reduction design solutions can be a challenge. Furthermore, quantification is often lacking in terms of heat accumulation for a given area, maximum acceptable levels of heat, and the quantity of needed measures to reduce UHI (number of trees or square meters of green space, for example) .
However, certain cities like Stuttgart have developed spatial parameters in urban planning guidelines with respect to climate change. In California a cool-roof material with low thermal admittance has been introduced in the Building Energy Efficiency Standard regulation of the state. The city of Portland is creating a reference guide of pavement options for low-use traffic zones. The greening policy in Chicago and Edinburgh involves increasing the number of street trees, as well as species heterogeneity to ensure resistance to vegetal disease (given that species diversity limits pest infestation).
Mitigating New York City’s heat island with urban forestry, living roofs and light surfaces, Rosenzwieg et al. 2006
Urban heat islands are created when solar energy is absorbed by non-reflective, impervious, and often rather dark surfaces, such as asphalt, stone, metal, and concrete, which are ubiquitous in cities. Exacerbating this solar energy absorption effect are abundant amounts of heat released from vehicles, factories and air conditioners, for example, as well as pollutants trapped in the lower troposphere that slow down the cooling of rising air.
In New York City, where this study was conducted, the “summertime nocturnal heat island averages 7.2ºF (4ºC). This means that during the summer months the daily minimum temperature in the city is on average 7.2ºF (4ºC) warmer than surrounding suburban and rural areas” [Rosenzwieg 2006: 1]. The authors tested the cooling effects of tree plantings, living rooftops and high albedo (light colored) surfaces, and found that curbside tree plantings were the most effective form of cooling per unit area, followed by living rooftops. High albedo (light/white) surfaces were the least effective at cooling per unit area, but were the most effective overall “because 64% of New York City’s surface area could be redeveloped from dark, impervious surfaces to lighter high-albedo surfaces” [Rosenzwieg 2006: 3], whereas only 17% of the city’s surface could be planted with new street trees.
The interaction of rivers and urban form in mitigating the Urban Heat Island effect: a UK case study, Hathaway & Sharples 2012
Like vegetative and light or reflective surfaces, water bodies have a cooling effect on cities, reducing the Urban Heat Island effect. The average temperature at the river in this study was 1C less than at a reference point elsewhere in the city. Furthermore, the form of the landscape on the banks of an urban river can either propagate (increase) or diminish the cooling effects of the river. This study found that vegetated river banks increased the cooling effect of the river by a difference of 2C compared to river banks covered in hard engineering materials (concrete/asphalt), while opening river banks (rather than enclosing them with buildings or walls) permitted significant cooling effects to be felt up to a distance of 30 meters from the river.
Overall, the results indicate that high levels of vegetation next to the river increase the cooling on the bank, that opening up the streets to the river increases the propagation of cooling and that the surface nature of the surrounding materials [e.g. vegetation versus concrete] can have a more significant effect on the air temperatures than the presence of the river [Hathaway & Sharples 2012: 20].
Overall, the results indicate that high levels of vegetation next to the river increase the cooling on the bank, that opening up the streets to the river increases the propagation of cooling and that the surface nature of the surrounding materials [e.g. vegetation versus concrete] can have a more significant effect on the air temperatures than the presence of the river [Hathaway & Sharples 2012: 20].
Urban development, land sharing and land sparing: the importance of considering restoration, Collas et al. 2017
With 66% of the world’s population predicted to live in cities by 2050, the challenge of reconciling urban growth with biodiversity conservation demands attention.
Although the environment is altered by urbanization, there is potential for cities to support a great deal of biodiversity [Collas 2017: 1866].
This study shows that urban growth and biodiversity enhancement are compatible by increasing housing density (in order to reduce total surface area of development) while restoring ecosystems on remaining green space through woodlot plantings. The study was conducted in Cambridge, England, whose population is expected to grow by 22% between 2011 and 2031, and where “current green space supports relatively few trees” [Collas 2017: 54]. Green space could be maximized and restored to woodlots while additional high-density housing could accommodate the expected population growth.
Furthermore, only 2% of green space (i.e. ≥30 ha) is needed for conversion to woodlot to increase the native tree population size in Cambridge while also increasing high-density housing. This is compared to an alternative growth scenario, where new development is low-density and inhabitants are expected to plant trees in their relatively large yards, while no city-led green space ecosystem restoration occurs.
In conclusion, the authors offer this:
For other cities in the UK and across Europe, which have generally long been cleared of natural habitat, restoration in parallel with the expansion of higher density housing would appear to offer greatest scope for accommodating population growth at least cost to nature. This would require policy and economic incentives to directly link high-intensity human land-use to large-scale restoration [Collas 2017: 1871].
Promoting and preserving biodiversity in the urban forest, Alvey 2006
Given the dangerous, precipitous global decline in biodiversity, coupled with rapid urbanization, cities have a key role to play in protecting biodiversity. In fact, cities already do harbor a large share of biodiversity. This may be due to the fact that cities are often situated in places of large inherent biodiversity (along rivers, for example), and/or because of large numbers of introduced species and landscape heterogeneity in cities. Furthermore, surrounding agricultural areas are often simplified landscapes with limited biodiversity while many forests are degraded, and thus less biodiverse, due to timber harvest regimes, roads, etc. Thus, contrary to what might be assumed, rural areas are not necessarily more biodiverse than cities.
The author stresses the importance of managing cities to increase biodiversity. This process should begin with a city-wide tree inventory to identify tree species, locations and health. Management should focus on increasing biodiversity among street trees, and in parks, woodlots, abandoned lots, and back/front yards, while also fostering public awareness and appreciation for ecological principles. Planting efforts should prioritize native species, which are better adapted to local conditions, are non-invasive, and whose protection contributes to global biodiversity conservation. (While great numbers of introduced species may increase local biodiversity, it has a homogenizing effect on global biodiversity.) Furthermore, natural regeneration of parks and woodlots should be encouraged through less intensive management, whereby seeds of native (or at least non-invasive) species are allowed to germinate and establish where they fall, instead of being fastidiously mowed or weeded.
A new vision for New Orleans and the Mississippi delta: applying ecological economics and ecological engineering, Costanza, Mitsch & Day 2006
What happened in New Orleans [during Hurricane Katrina], while a terrible “natural” disaster, was also the cumulative result of excessive and inappropriate management of the Mississippi River and delta, inadequate emergency preparation, a failure to act in time on plans to restore the wetlands and storm protection levees, and the expansion of the city into increasingly vulnerable areas [Costanza, Mitsch & Day 2006: 467].
Mismanagement here refers to damming, leveeing and canal dredging of the Mississippi River Delta, resulting in a significant loss of wetlands and the erosion of barrier islands over the past 100-plus years. Coastal marshes and barrier islands depend on regular inputs of sediments deposited by the river, which has been isolated from the delta plain and unable to thus nourish it. Two thirds of the river empties directly into the depths of the Gulf of Mexico, while one third empties into shallow waters, where it nourishes wetlands via the Atchafalaya, the river’s single remaining distributary (other distributaries having been closed off).
Damage from Hurricane Katrina was exacerbated by its prior loss of wetlands. Expansive coastal wetlands protect coastal communities from hurricanes by “decreasing the area of open water (fetch) for wind to form waves, increasing drag on water motion and hence the amplitude of a storm surge, reducing direct wind effect on the water surface, and directly absorbing wave energy” [Costanza, Mitsch & Day 2006: 468].
For the rebuilding of New Orleans after the hurricane, the authors recommended several core principles aimed at social and ecological resilience. Among their recommendations, they advise that areas of the city currently below sea level (by as much as 5 meters in some parts) not be rebuilt, but, rather, be restored to wetland. This would allow for temporary water storage within the city, water filtration, and biodiversity protection. They also suggest the reopening of distributaries and the controlled breaching of certain levees to allow the river to resume its ancient task of distributing sediment over a greater expanse of coastal marshes, allowing these marshes to gradually rise in step with sea level rise.
Eco-engineering urban infrastructure for marine and coastal biodiversity: which interventions have the greatest ecological benefit? Strain et al. 2017
While the majority of people on Earth live in cities, the majority (60%) of the world’s largest cities are located within 100 kilometers of a coast. The pollution and urban infrastructure (such as marinas, sea walls, or oil/gas platforms) emanating from cities greatly stresses coastal marine habitats. Coastal infrastructure tends to be vertical and smooth, offering little or nothing in the way of habitat niches or physical protection for various marine organisms. An eco-engineering approach to improve habitat quality and increase biodiversity is the addition of textural features, such as ledges, small holes, basins or crevices to the hard surfaces of urban marine infrastructure.
As predicted, overall microhabitat-enhancing interventions had a positive effect on the abundance and number of species across the studies. Nevertheless, the magnitude of their effects varied considerably, from zero to highly positive according to the type of intervention, the target taxa, and tidal elevation [Strain 2017: 434].
In the intertidal[5], interventions that provided moisture and shade had the greatest effect on the richness of sessile[6] and mobile organisms, while water-retaining features had the greatest effect on the richness of fish. In contrast, in the subtidal[7], small-scale depressions which provide refuge to new recruits from predators and other environmental stressors such as waves, had higher abundances of sessile organisms while elevated structures had higher numbers and abundances of fish. The taxa that responded most positively to eco-engineering in the intertidal were those whose body size most closely matched the dimensions of the resulting intervention [Strain 2017: 426].
Coastal adaptation with ecological engineering, Cheong et al. 2013
Because of the multiple threats and uncertainties of a changing climate, protecting coastal areas simply by building new seawalls (or some other such inflexible, single-tactic approach) is unlikely to be the most effective option. Instead, combined coastal adaptation strategies to allow for a dynamic response to multiple stressors are increasingly preferred. Climate scientists and coastal managers are mainstreaming inclusion of climate change into an Integrated Coastal Zone Management framework, aimed at promoting the activities of the different coastal sectors by coordinating government agencies and private participation.
Contrary to a “regret-risking option,” a no- or low-regret option is adopted to generate a net social benefit irrespective of the future outcome of climate change. Revamping early warning systems, preventing land reclamation, improving housing and transportation, capacity development in education, poverty reduction, and efforts to build resilient ecosystems are examples of a low- or no-regret options.
Traditional engineering, while sometimes protective of coastal communities, has undesired effects, such as eroding non-target, neighboring coastline and destroying adjacent ecosystems. By contrast, eco-engineering tools emphasize positive interactions among species that boost ecosystem productivity and stability, and therefore the strength of the ecosystem to withstand and buffer heavy storms, thus protecting coastal communities.
For example, sea-grasses planted with clams at their roots grow faster and in turn increase total fixed carbon. Oyster reefs attenuate up to 95% of wave height, control turbidity by removing algae, bacteria, and suspended organic matter, improve water quality through their filtration capacity, and enable seafood supply and thus job creation and recreation. Oyster reefs also support breeding ground for economically valued species, such as blue crab, red drums, flounder and spotted sea trout.
In mangroves, transplants planted in close proximity rather than the traditional spread pattern allows for a shared benefit of positive interaction that enhances plant growth and biodiversity. Restored mangrove ecosystems alleviate the impact of moderate tsunami waves, while the roots trap sediment and elevate the land surface, allowing for adaptation to sea-level rise. Intact mangrove also provides local employment as well as breeding grounds for fish.
Marshes dampen wave actions and reduce shoreline erosion, increase fish production, and are compatible with levee designs on the marshes’ landward edges that are nature-friendly. In the Netherlands, for instance, levees built to prevent flooding during storms were covered with thick grass to maintain their integrity, while the seaward marshes reduce the levees’ exposure to wave action; grasses were then grazed by sheep to provide milk and meat for consumption.
The synergy of ecology and engineering is key to addressing uncertainties related to climate-induced stressors. The combination of traditional and eco-engineering approaches coupled with the evaluation to measure the effectiveness of eco-engineered structures facilitate better decision making and prioritization of options.
Where we stand: climate action, The American Institute of Architects (AIA) 2019b
Noting that 40% of carbon emissions in the US come from the construction (including sourcing of materials) and operation (heating, cooling, lighting) of buildings and houses, the AIA pledges to achieve zero-carbon construction and operation of all new buildings, and retrofitting of existing buildings to reduce their energy use and increase their resilience to severe weather. They will achieve these goals through education, policy advocacy, calling for zero-carbon building codes, and advocating for the reuse of historic buildings rather than new construction.
Living Building Challenge Standard, June 2019
The construction and operation of buildings and houses is a major source of pollution and ecosystem destruction around the world. In light of this, the Living Building Challenge invites people to reimagine the built environment as a source of social and ecological regeneration.
Nothing less than a sea change in building, infrastructure and community design is required. Indeed, this focus needs to be the great work of our generation. We must remake our cities, towns, neighborhoods, homes and offices, and all the spaces and infrastructure in between. This is part of the necessary process of reinventing our relationship with the natural world and each other—reestablishing ourselves as not separate from, but part of nature, “because the living environment is what really sustains us” (E.O. Wilson) [International Living Future Institute 2019: 8].
To that end the Living Building Challenge invites us to collaborate in building houses and buildings – or adapting existing ones – to have a positive, rather than simply less-negative, impact on the social and ecological systems where they are situated. The initiative runs educational and certification programs with several high standards, including, for example:
Projects must be observant of and responsive to the local ecological and social context of the sites, and onsite landscaping must seek to emulate local ecosystem function.
Access to locally grown food should be assured through onsite production and/or connection to local farms.
The site must ensure adequate habitat for local species.
Living Building Challenge designers must find ways to encourage pedestrian, bike and public transport options, while discouraging individual car travel.
Water should be harvested and wastewater treated onsite using living or natural/non-chemical systems.
Buildings/houses should supply their own energy on site (not through combustion), monitor their energy use, and minimize use through conservation.
Construction materials should be salvaged or sustainably and transparently sourced, and non-toxic.
In the interest of human wellbeing and social equity designs should allow natural light, beauty and comfort in the interior of the building, while the exterior must be accessible and welcoming to all members of the public, regardless of socioeconomic status.
Adapt now: a global call for leadership on climate resilience, Global Commission on Adaptation, September 2019
This report, led by Ban Ki Moon (UN), Bill Gates (Bill & Melinda Gates Foundation) and Kristalina Georgieva (World Bank), calls on decision makers worldwide to facilitate coordinated action to help communities adapt to climate change. Importantly, the report makes the case for nature-based adaptation approaches, which inherently help mitigation efforts as well. Adaptation measures are much cheaper than recovery and rebuilding: every $1 invested in adaption yields $2-10 (or more by some estimates) in avoided losses and other economic benefits (such as improved crop yields), as well as social and environmental benefits.
Despite a clear global imperative for rapid adaptation planning and action to be taken at local, regional and national levels, action is desperately lagging. The report cites four reasons for inaction: (1) broad failure to internalize climate change risk in everyday decision making; (2) human tendency to prioritize short-term planning at the expense of long-term goals; (3) lack of cross-sector collaboration, which leads to fragmentation of responsibility; and (4) lack of power/voice among those most affected by climate change.
The report succinctly articulates the value of working with nature to adapt to climate change, while highlighting the extent to which this vital information is neglected.
We can already see the immense opportunity of using nature to increase societal resilience in landscapes ranging from uplands to the ocean. Restoring upland forests and watersheds could save water utilities in the world’s 534 largest cities an estimated $890 million each year and is critical for regulating water flows and managing the future’s more extreme floods. Meanwhile, lakes, marshes, and river floodplains both slow the release of floodwater and filter out sediment. The Netherlands has harnessed these capabilities with a Room for the River strategy that increases capacity of rivers and their floodplains to hold floodwaters, reducing damage and loss of life.
Ecosystem restoration also is a powerful tool for feeding the hungry, cooling sweltering cities, and protecting communities. One striking example is farmer-led reforestation in the Maradi and Zinder regions of Niger, which has boosted crop yields, improved soil fertility, and lifted communities out of poverty. Tree cover has soared ten-fold and the daily time spent gathering firewood—a task that mainly falls to women—has dropped from 3 hours to 30 minutes. For cities, an annual investment of $100 million in urban tree planting could create enough shade to cut average temperatures by 1°C for 77 million people around the world. Restoring the mangrove forests that offer protections from rising seas and storm surges is two to five times cheaper than building engineered structures like underwater breakwaters, while also storing carbon and improving water quality and local fisheries.
Yet despite the powerful case for working with nature to reduce climate risks, the world has barely begun to realize this potential. Few governments have adopted these approaches widely, even though many cite natural solutions in their NDCs. And only 3 percent of nearly 2,000 companies reported using natural ecosystems as part of their climate adaptation strategies. The barriers include lack of awareness of the critical role of natural assets in underpinning social and economic resilience and lack of accessible funds to invest in nature-based solutions. In addition, the piecemeal way adaptation is often planned and executed undervalues or ignores the many benefits of working with nature.
Humanity faces a stark choice: We can harness nature-based solutions to mitigate climate change and to better adapt—or we can continue with business as usual and lose the essential and myriad services nature provides [Global Commission on Climate Adaptation 2019: 31].
To encourage adoption of nature-based adaptation strategies, the report recommends three steps: (1) raise the level of understanding of the value of nature for climate adaptation; (2) embed nature-based solutions into adaptation planning and policy; and (3) increase investment into nature-based solutions. Indeed, it is precisely the aim of Biodiversity for a Livable Climate and its compendium series to elevate the level of understanding and appreciation for nature-based adaptation and mitigation solutions to the climate crisis.
Yet despite the powerful case for working with nature to reduce climate risks, the world has barely begun to realize this potential. Few governments have adopted these approaches widely, even though many cite natural solutions in their NDCs. And only 3 percent of nearly 2,000 companies reported using natural ecosystems as part of their climate adaptation strategies. The barriers include lack of awareness of the critical role of natural assets in underpinning social and economic resilience and lack of accessible funds to invest in nature-based solutions. In addition, the piecemeal way adaptation is often planned and executed undervalues or ignores the many benefits of working with nature.
Humanity faces a stark choice: We can harness nature-based solutions to mitigate climate change and to better adapt—or we can continue with business as usual and lose the essential and myriad services nature provides [Global Commission on Climate Adaptation 2019: 31].
We begin this issue of the Compendium by exploring the role of cities in the era of climate breakdown. This section features “Heat Planet,” an essay by architect Christopher Haines, member of Bio4Climate’s Leadership Team, exploring the global implications of the pervasive phenomenon of the “Urban Heat Island” and other heat-producing paved and de-vegetated surfaces around the globe. We then explore various general, non-urban land conservation priorities (intact forests, wilderness areas, large old trees, and habitat corridors), and finish with a sampling of stories of real communities restoring their local ecosystems.
The Gaia hypothesis invites us to imagine Earth as an integral living system in order to explore the mechanisms by which life helps create and maintain the conditions for life, such as an oxygenated atmosphere.
“The Gaia theory proposes that organisms contribute to self-regulating feedback mechanisms that have kept the Earth’s surface environment stable and habitable to life” [Lenton 2000: 439]. This theory was developed by James Lovelock, a chemist who observed that Earth’s atmosphere is in a constant state of disequilibrium,
in which highly reactive gases, such as methane and oxygen, exist together at levels that are different by many orders of magnitude from photochemical steady states. Large, biogenic fluxes of gases are involved in maintaining such disequilibrium. This perturbed state is remarkable in that the atmospheric composition is fairly stable over periods of time that are much longer than the residence times of the constituent gases, indicating that life may regulate the composition of the Earth’s atmosphere. This concept became the foundation of Gaia theory [Lenton 2000: 439].
The theory is based on the evolutionary biology concept of natural selection, focusing on traits that alter the environment and the resulting feedback from that environmental change on the organisms with the traits that produced it. Lenton offers a few examples to illustrate such feedbacks, starting with the “Daisyworld” model, where black pigment in daisies confers advantage in an environment with below-optimal temperatures. By absorbing heat, the black daisies grow better than their white counterparts and their population dominates. The global effect of a growing population of individually warm daisies raises the overall temperature of the world. At this point, the population of white daisies begins to rebound and the global temperatures cool again.
Gaia theory aims to be consistent with evolutionary biology and views the evolution of organisms and their material environment as so closely coupled that they form a single, indivisible, process. Organisms possess environment-altering traits because the benefit that these traits confer (to the fitness of the organisms) outweighs the cost in energy to the individual [Lenton 2000: 440].
Some activities that alter the environment are so advantageous (to the organisms carrying out the activities) that they become widespread, fundamental properties of organisms. (An example is photosynthesis, the implications of which have been studied by modelling the Archaean–Proterozoic transition.) Other activities are favorable only under particular environmental conditions and hence are subject to selection. In such cases, it is often changes in one environmental variable that determine whether a trait remains selectively favorable. If the spread of the trait alters this environmental variable, it also alters the forces of selection determining its own value [Lenton 2000: 442].
Furthermore, ecosystems-level environmental feedbacks can be understood in terms of natural selection. For example:
The trees of the Amazon rainforest, through generating a high level of water cycling, maintain the moist environmental conditions in which they can persist (a positive feedback on growth and selection). Nutrients are also effectively retained and recycled. If too much forest is removed, the water-regulation system can collapse, the topsoil is washed away and the region reverts to arid semi-desert, a change that may be difficult to reverse [Lenton 2000: 445].
Lenton explains that while there are geochemical mechanisms involved in regulating the climate, “it is clear that organisms are involved in many environmental feedbacks on Earth, and their effects need to be considered” [Lenton 2000: 441]. For example, acid rain weathers calcium-silicate rocks resulting in the formation of calcium carbonate by removing carbon dioxide from the atmosphere, thus cooling the Earth. Warmer average global temperatures would lead to more rain, thus more weathering and the cooling effects of that negative, self-correcting feedback. “However, geochemical feedbacks [such as this one] operate slowly and are not very responsive to perturbation” [Lenton 2000: 441]. Rock weathering organisms can amplify the weathering effects of the rain, hastening the negative feedback. Thus, there’s an intertwining of processes that regulate the climate.
The significance of this article is that if life has been at least partly responsible for creating and maintaining the habitability of Earth’s climate for the past 3.5 billion years, then it has a key role to play today as we grapple with how to keep global temperatures from rising above 1.5C. Ecosystems are clearly victimized by climate chaos, while also being directly damaged by avoidable human activity, such as land-clearing for development and agriculture and the ubiquitous use of chemical toxins and plastics. Yet if ecosystems are also a driver of climatic conditions, then it is critical to protect them from further harm and to nurture their growth and stability. Humans can become Gaia’s nursing team – we can improve the conditions for her recovery to the point when her own systems kick in and bring her back to health.
Geomorphology is the study of landforms and processes and how they developed. This conceptual commentary proposes that the emergence and evolution of life, especially vegetation, has played a major role in physically shaping the Earth. For example, plant roots trap and hold sediment (preventing erosion), resulting in the formation of hillsides, sand dunes, fluvial islands, river banks, floodplains, and river channels, for example. Without vegetation to hold sediment in place, it would be blown or washed away, creating different land patterns. Roots also contribute to rock weathering, resulting in soil formation and even the formation of marine black shale, while aboveground, plants create a rough surface which affects flows of matter and energy. Indirectly, in being the primary source of energy for animals and microorganisms, plants “also control geomorphic processes through their engineering activities in soils and at the surface of the Earth.”
Trees play a central geomorphological role:
In particular the development of the lignin-containing plants (shrubs and trees) in the middle Devonian (380 Ma[9]) have produced the most significant geomorphic changes. Their complex and resistant root and stem systems combined with their slower decomposition has contributed to increase global sediment stability and storage in time and in space on the Earth’s surface [Corenblit & Steiger 2009: 894].
The authors contextualize the role of life in geomorphology in terms of energy sources available to do geomorphic work. Vegetation dynamics, driven by photosynthesis, which converts solar energy into stored chemical energy, is one of four such energy sources. The other three are: gravity, solar energy, and geothermal activity. This article helps us to visualize Earth’s systems (in particular lithosphere and biosphere) as interwoven, where biology drives not only life processes, but land formation process as well.
The Great Barrier Reef (GBR) protects northeast Australia from wave exposure, while also creating habitat for a vast expanse of shallow- and deep-water seagrasses between the reef and the shoreline. Deep-water seagrasses here occupy an area roughly the size of Switzerland. While the carbon storage capacity of shallow-water seagrasses, dubbed ‘blue carbon,’ are known to be extremely high, the amounts of carbon stored in deep-water seagrasses (greater than 15 meters depth) is less well known, and expected to be lower due to these plants’ smaller stature and relative sparseness.
The authors found, however, that “deep-water seagrass contained similar levels of organic carbon (OC) to shallow-water species, despite being much sparser and smaller in stature” [York 2018: 1]. Furthermore, deep-water seagrass sediments contained about nine times more OC than surrounding bare areas.
If the OC stocks reported in this study are similar to deep-water [seagrass] meadows elsewhere within the GBR lagoon, then OC bound within this system is roughly estimated at 27.4 million tons [York 2018: 1].
When Australian authorities removed indigenous Martu people from their traditional lands in the desertic center of the continent in the mid-1900s, endemic species there declined or went extinct. Researchers observed that the Martu’s hunting regime of small burning patches of land reduced the size of wildfires while also boosting populations of native species such as dingo, monitor lizard and kangaroo. The absence of the Martu after the 1950s resulted in domination by invasive species, which killed much of the native wildlife.
Indigenous hunters have positive impacts on food webs in desert Australia, Penn State 2019
When Australian authorities removed indigenous Martu people from their traditional lands in the desertic center of the continent in the mid-1900s, endemic species there declined or went extinct. Researchers observed that the Martu’s hunting regime of small burning patches of land reduced the size of wildfires while also boosting populations of native species such as dingo, monitor lizard and kangaroo. The absence of the Martu after the 1950s resulted in domination by invasive species, which killed much of the native wildlife.
Blue carbon stocks of Great Barrier Reef deep-water seagrasses, York et al. 2018
The Great Barrier Reef (GBR) protects northeast Australia from wave exposure, while also creating habitat for a vast expanse of shallow- and deep-water seagrasses between the reef and the shoreline. Deep-water seagrasses here occupy an area roughly the size of Switzerland. While the carbon storage capacity of shallow-water seagrasses, dubbed ‘blue carbon,’ are known to be extremely high, the amounts of carbon stored in deep-water seagrasses (greater than 15 meters depth) is less well known, and expected to be lower due to these plants’ smaller stature and relative sparseness.
The authors found, however, that “deep-water seagrass contained similar levels of organic carbon (OC) to shallow-water species, despite being much sparser and smaller in stature” [York 2018: 1]. Furthermore, deep-water seagrass sediments contained about nine times more OC than surrounding bare areas.
If the OC stocks reported in this study are similar to deep-water [seagrass] meadows elsewhere within the GBR lagoon, then OC bound within this system is roughly estimated at 27.4 million tons [York 2018: 1].
Vegetation as a major conductor of geomorphic changes on the Earth surface: toward evolutionary geomorphology, Corenblit & Steiger 2009
Geomorphology is the study of landforms and processes and how they developed. This conceptual commentary proposes that the emergence and evolution of life, especially vegetation, has played a major role in physically shaping the Earth. For example, plant roots trap and hold sediment (preventing erosion), resulting in the formation of hillsides, sand dunes, fluvial islands, river banks, floodplains, and river channels, for example. Without vegetation to hold sediment in place, it would be blown or washed away, creating different land patterns. Roots also contribute to rock weathering, resulting in soil formation and even the formation of marine black shale, while aboveground, plants create a rough surface which affects flows of matter and energy. Indirectly, in being the primary source of energy for animals and microorganisms, plants “also control geomorphic processes through their engineering activities in soils and at the surface of the Earth.”
Trees play a central geomorphological role:
In particular the development of the lignin-containing plants (shrubs and trees) in the middle Devonian (380 Ma[9]) have produced the most significant geomorphic changes. Their complex and resistant root and stem systems combined with their slower decomposition has contributed to increase global sediment stability and storage in time and in space on the Earth’s surface [Corenblit & Steiger 2009: 894].
The authors contextualize the role of life in geomorphology in terms of energy sources available to do geomorphic work. Vegetation dynamics, driven by photosynthesis, which converts solar energy into stored chemical energy, is one of four such energy sources. The other three are: gravity, solar energy, and geothermal activity. This article helps us to visualize Earth’s systems (in particular lithosphere and biosphere) as interwoven, where biology drives not only life processes, but land formation process as well.
Gaia and natural selection, Lenton 1998
The Gaia hypothesis invites us to imagine Earth as an integral living system in order to explore the mechanisms by which life helps create and maintain the conditions for life, such as an oxygenated atmosphere.
“The Gaia theory proposes that organisms contribute to self-regulating feedback mechanisms that have kept the Earth’s surface environment stable and habitable to life” [Lenton 2000: 439]. This theory was developed by James Lovelock, a chemist who observed that Earth’s atmosphere is in a constant state of disequilibrium,
in which highly reactive gases, such as methane and oxygen, exist together at levels that are different by many orders of magnitude from photochemical steady states. Large, biogenic fluxes of gases are involved in maintaining such disequilibrium. This perturbed state is remarkable in that the atmospheric composition is fairly stable over periods of time that are much longer than the residence times of the constituent gases, indicating that life may regulate the composition of the Earth’s atmosphere. This concept became the foundation of Gaia theory [Lenton 2000: 439].
The theory is based on the evolutionary biology concept of natural selection, focusing on traits that alter the environment and the resulting feedback from that environmental change on the organisms with the traits that produced it. Lenton offers a few examples to illustrate such feedbacks, starting with the “Daisyworld” model, where black pigment in daisies confers advantage in an environment with below-optimal temperatures. By absorbing heat, the black daisies grow better than their white counterparts and their population dominates. The global effect of a growing population of individually warm daisies raises the overall temperature of the world. At this point, the population of white daisies begins to rebound and the global temperatures cool again.
Gaia theory aims to be consistent with evolutionary biology and views the evolution of organisms and their material environment as so closely coupled that they form a single, indivisible, process. Organisms possess environment-altering traits because the benefit that these traits confer (to the fitness of the organisms) outweighs the cost in energy to the individual [Lenton 2000: 440].
Some activities that alter the environment are so advantageous (to the organisms carrying out the activities) that they become widespread, fundamental properties of organisms. (An example is photosynthesis, the implications of which have been studied by modelling the Archaean–Proterozoic transition.) Other activities are favorable only under particular environmental conditions and hence are subject to selection. In such cases, it is often changes in one environmental variable that determine whether a trait remains selectively favorable. If the spread of the trait alters this environmental variable, it also alters the forces of selection determining its own value [Lenton 2000: 442].
Furthermore, ecosystems-level environmental feedbacks can be understood in terms of natural selection. For example:
The trees of the Amazon rainforest, through generating a high level of water cycling, maintain the moist environmental conditions in which they can persist (a positive feedback on growth and selection). Nutrients are also effectively retained and recycled. If too much forest is removed, the water-regulation system can collapse, the topsoil is washed away and the region reverts to arid semi-desert, a change that may be difficult to reverse [Lenton 2000: 445].
Lenton explains that while there are geochemical mechanisms involved in regulating the climate, “it is clear that organisms are involved in many environmental feedbacks on Earth, and their effects need to be considered” [Lenton 2000: 441]. For example, acid rain weathers calcium-silicate rocks resulting in the formation of calcium carbonate by removing carbon dioxide from the atmosphere, thus cooling the Earth. Warmer average global temperatures would lead to more rain, thus more weathering and the cooling effects of that negative, self-correcting feedback. “However, geochemical feedbacks [such as this one] operate slowly and are not very responsive to perturbation” [Lenton 2000: 441]. Rock weathering organisms can amplify the weathering effects of the rain, hastening the negative feedback. Thus, there’s an intertwining of processes that regulate the climate.
The significance of this article is that if life has been at least partly responsible for creating and maintaining the habitability of Earth’s climate for the past 3.5 billion years, then it has a key role to play today as we grapple with how to keep global temperatures from rising above 1.5C. Ecosystems are clearly victimized by climate chaos, while also being directly damaged by avoidable human activity, such as land-clearing for development and agriculture and the ubiquitous use of chemical toxins and plastics. Yet if ecosystems are also a driver of climatic conditions, then it is critical to protect them from further harm and to nurture their growth and stability. Humans can become Gaia’s nursing team – we can improve the conditions for her recovery to the point when her own systems kick in and bring her back to health.
iMatter is a tight-knit national group of passionate pre-college individuals who are making real impacts in their communities. They are showing up in city halls and state offices, demanding their elected officials at every level possible commit to bold and visionary climate action. Students from Brookline High School in Massachusetts submitted resolutions to their town legislators, saying they’re worried about their future and the future of the environment; their cities agreed and are supporting the Green New Deal. Alec Loorz started the organization Kids vs. Global Warming in California with his mom, Victoria Loorz in 2007, when he was 13 years old. The organization eventually changed its name to iMatter. Alec went on to spearhead the Our Children’s Trust lawsuit against federal and state governments of the United States to secure climate recovery plans that will restore the balance of Earth’s climate systems.
The deep winter snow falls on the mountains around the Methow Valley in the state of Washington are declining. To manage problems with drought, the Methow Beaver Project has been capturing, tagging, matching male and female beavers and releasing them in key valley areas. The project workers know beavers are master engineers that know how to preserve their homes and food supply, to the benefit of water quality and many other animals and plants in the area. Beaver reintroduction projects are also underway in Nevada, Utah and Wyoming. In 2018, Scott Helker, a Libertarian candidate running to become governor of Colorado, was asked, “Would you support asking Coloradans to raise billions of tax dollars for projects that would increase water supplies and help prevent a projected water shortage mid-century?” He answered, “No. I can create the same results without raising billions of dollars. Ask me how.” Answer: with beaver reintroduction projects.
On the other side of the Amazon and across the South Atlantic Ocean, the small South African community of Xolobeni won a similar court case. Like the Waorani, the people of Xolobeni demanded that they be consulted rather than being forced to cede their land to mining interests – in this case to an Australian titanium mining company. Also like the Waorani, they were defending not only their lives, livelihoods, their health and wellbeing, but also an ecologically rich corner of the planet. Xolobeni is in Pondoland, a dune-covered stretch of the coast that is home to endemic species and frequented offshore by whales.
The law says we have a right to be consulted, but what we say doesn’t seem to matter. We have told the company many times that we don’t want their mine. How many times do we have to say no?
– Nonhle Mbuthuma, local resident
The court agreed that local communities must give their consent before mining is allowed on their land.
When the Waorani people of the Ecuadorian Amazon heard their government was planning to sell drilling rights to their land to international oil companies, they mobilized. They mapped the land to illustrate to the Western world its otherwise unseen cultural, historical and ecological richness. These maps include “historic battle sites, ancient cave-carvings, jaguar trails, medicinal plants, animal reproductive zones, important fishing holes, creek-crossings and sacred waterfalls,” according to an online petition they launched in partnership with the NGO Amazon Frontlines. Then the Waorani sued the government for not properly consulting them when the decision was made in 2012 to dice up the rainforest into auctionable blocks of land. In April 2019, the Ecuadorian court ruled in favor of the Waorani, immediately suspending any sale of the land and setting a precedent for other communities resisting oil extraction in their lands.
The government’s interests in oil is not more valuable than our rights, our forests, our lives.
– Nemonte Nenquimo, one of the Waorani plaintiffs and representative of the Coordinating Council of the Waorani Nationality Ecuador Pastaza (CONCONAWEP).
The Waorani win follows a win against mining operations last year by the indigenous Kofan community also in the Ecuadorian Amazon.
At the age of 15, Greta Thunberg began sitting on the steps of the Swedish parliament with a handmade sign reading: “skolstrejk för klimatet” or “school strike for the climate.” The decision to act came about seven years after she first learned of climate change. The fact that adults didn’t seem bothered to do anything about the global crisis shocked her, and then sent her into a depression. Activism pulled her out of depression and thrust her onto the international stage. It didn’t take long for her solo picketing efforts to spark a global movement spanning 125 countries of more than a million kids striking from school for climate. Greta intends to continue striking outside the Swedish Parliament until it passes legislation that upholds commitments made in the Paris Climate Accord.
Excerpted from a Guardian guest editorial by climate strikers Greta Thunberg (Sweden), Anna Taylor (UK), Luisa Neubauer (Germany), Kyra Gantois, Anuna De Wever and Adélaïde Charlier (Belgium), Holly Gillibrand (Scotland), and Alexandria Villasenor (USA):
This movement had to happen, we didn’t have a choice. The vast majority of climate strikers taking action today aren’t allowed to vote. Imagine for a second what that feels like. Despite watching the climate crisis unfold, despite knowing the facts, we aren’t allowed to have a say in who makes the decisions about climate change. And then ask yourself this: wouldn’t you go on strike too, if you thought doing so could help protect your own future?
So today we walk out of school, we quit our college lessons, and we take to the streets to say enough is enough. Some adults say we shouldn’t be walking out of classes – that we should be “getting an education”. We think organising against an existential threat – and figuring out how to make our voices heard – is teaching us some important lessons.
Hearing of plans underway for a four-lane highway near their home in Nantes, France, local residents Jim and Stephanie responded by planting a small forest. The idea was not only to block out the added sound and air pollution, but also to try to compensate for the assault on the planet of any road expansion. The couple was inspired by Shubhendhu Sharma, who spoke at the 2018 Nantes festival Aux Arbes[8]. Sharma showed the audience how 300 trees of 30-some species could be planted in the space of six parking places. He described the Miyawaki Method, which mimics natural forests in terms of biodiversity and density, outperforms the growth rate of monoculture plantations tenfold, and works well in urban areas because it takes so little space. Within the following year, Jim and Stephanie, along with dozens of volunteers and school kids, planted more than 2000 trees on two sites. To encourage Miyawaki-style afforestation projects everywhere, the couple launched the initiative Minibigforest; this is only the beginning for them.
The following sketches are but a tiny sampling of the countless ways people throughout the world push back against the socio-economic and political forces of destruction both of ecosystems and of the social fabric of society. Adopting Paul Hawken’s terminology and characterization of “blessed unrest” as a spontaneous, decentralized global social movement, we here present a diverse though far from representative series of vignettes of everyday heroes. May such stories light the fire for new heroes to perpetually emerge in defense of all life on Earth.
Minibigforest in Nantes
Hearing of plans underway for a four-lane highway near their home in Nantes, France, local residents Jim and Stephanie responded by planting a small forest. The idea was not only to block out the added sound and air pollution, but also to try to compensate for the assault on the planet of any road expansion. The couple was inspired by Shubhendhu Sharma, who spoke at the 2018 Nantes festival Aux Arbes[8]. Sharma showed the audience how 300 trees of 30-some species could be planted in the space of six parking places. He described the Miyawaki Method, which mimics natural forests in terms of biodiversity and density, outperforms the growth rate of monoculture plantations tenfold, and works well in urban areas because it takes so little space. Within the following year, Jim and Stephanie, along with dozens of volunteers and school kids, planted more than 2000 trees on two sites. To encourage Miyawaki-style afforestation projects everywhere, the couple launched the initiative Minibigforest; this is only the beginning for them.
Greta Thunberg and a million international student strikers
At the age of 15, Greta Thunberg began sitting on the steps of the Swedish parliament with a handmade sign reading: “skolstrejk för klimatet” or “school strike for the climate.” The decision to act came about seven years after she first learned of climate change. The fact that adults didn’t seem bothered to do anything about the global crisis shocked her, and then sent her into a depression. Activism pulled her out of depression and thrust her onto the international stage. It didn’t take long for her solo picketing efforts to spark a global movement spanning 125 countries of more than a million kids striking from school for climate. Greta intends to continue striking outside the Swedish Parliament until it passes legislation that upholds commitments made in the Paris Climate Accord.
Excerpted from a Guardian guest editorial by climate strikers Greta Thunberg (Sweden), Anna Taylor (UK), Luisa Neubauer (Germany), Kyra Gantois, Anuna De Wever and Adélaïde Charlier (Belgium), Holly Gillibrand (Scotland), and Alexandria Villasenor (USA):
This movement had to happen, we didn’t have a choice. The vast majority of climate strikers taking action today aren’t allowed to vote. Imagine for a second what that feels like. Despite watching the climate crisis unfold, despite knowing the facts, we aren’t allowed to have a say in who makes the decisions about climate change. And then ask yourself this: wouldn’t you go on strike too, if you thought doing so could help protect your own future?
So today we walk out of school, we quit our college lessons, and we take to the streets to say enough is enough. Some adults say we shouldn’t be walking out of classes – that we should be “getting an education”. We think organising against an existential threat – and figuring out how to make our voices heard – is teaching us some important lessons.
The Waorani people stand up for their rainforest homeland
When the Waorani people of the Ecuadorian Amazon heard their government was planning to sell drilling rights to their land to international oil companies, they mobilized. They mapped the land to illustrate to the Western world its otherwise unseen cultural, historical and ecological richness. These maps include “historic battle sites, ancient cave-carvings, jaguar trails, medicinal plants, animal reproductive zones, important fishing holes, creek-crossings and sacred waterfalls,” according to an online petition they launched in partnership with the NGO Amazon Frontlines. Then the Waorani sued the government for not properly consulting them when the decision was made in 2012 to dice up the rainforest into auctionable blocks of land. In April 2019, the Ecuadorian court ruled in favor of the Waorani, immediately suspending any sale of the land and setting a precedent for other communities resisting oil extraction in their lands.
The government’s interests in oil is not more valuable than our rights, our forests, our lives.
– Nemonte Nenquimo, one of the Waorani plaintiffs and representative of the Coordinating Council of the Waorani Nationality Ecuador Pastaza (CONCONAWEP).
The Waorani win follows a win against mining operations last year by the indigenous Kofan community also in the Ecuadorian Amazon.
On the other side of the Amazon and across the South Atlantic Ocean, the small South African community of Xolobeni won a similar court case. Like the Waorani, the people of Xolobeni demanded that they be consulted rather than being forced to cede their land to mining interests – in this case to an Australian titanium mining company. Also like the Waorani, they were defending not only their lives, livelihoods, their health and wellbeing, but also an ecologically rich corner of the planet. Xolobeni is in Pondoland, a dune-covered stretch of the coast that is home to endemic species and frequented offshore by whales.
The law says we have a right to be consulted, but what we say doesn’t seem to matter. We have told the company many times that we don’t want their mine. How many times do we have to say no?
– Nonhle Mbuthuma, local resident
The court agreed that local communities must give their consent before mining is allowed on their land.
Methow Beaver Project:enlisting beavers to make wetlands in compensation for declining mountain snowpack
The deep winter snow falls on the mountains around the Methow Valley in the state of Washington are declining. To manage problems with drought, the Methow Beaver Project has been capturing, tagging, matching male and female beavers and releasing them in key valley areas. The project workers know beavers are master engineers that know how to preserve their homes and food supply, to the benefit of water quality and many other animals and plants in the area. Beaver reintroduction projects are also underway in Nevada, Utah and Wyoming. In 2018, Scott Helker, a Libertarian candidate running to become governor of Colorado, was asked, “Would you support asking Coloradans to raise billions of tax dollars for projects that would increase water supplies and help prevent a projected water shortage mid-century?” He answered, “No. I can create the same results without raising billions of dollars. Ask me how.” Answer: with beaver reintroduction projects.
iMatter is a tight-knit national group of passionate pre-college individuals who are making real impacts in their communities. They are showing up in city halls and state offices, demanding their elected officials at every level possible commit to bold and visionary climate action. Students from Brookline High School in Massachusetts submitted resolutions to their town legislators, saying they’re worried about their future and the future of the environment; their cities agreed and are supporting the Green New Deal. Alec Loorz started the organization Kids vs. Global Warming in California with his mom, Victoria Loorz in 2007, when he was 13 years old. The organization eventually changed its name to iMatter. Alec went on to spearhead the Our Children’s Trust lawsuit against federal and state governments of the United States to secure climate recovery plans that will restore the balance of Earth’s climate systems.
Taking an altogether different angle, Jason Bradford of the Post Carbon Institute assumes radical societal change is inevitable and imminent, and focuses not on how to precipitate change but instead on how to adapt to it. “The future is rural” [Bradford 2019] is essentially a primer on how to navigate the profound changes society will undergo during the 21st Century due to climate breakdown and resource scarcity. It begins with an assertion that today’s “mass urbanization has been made possible by the prodigious exploitation of fossil fuels.” In other words,
Due to the concentrated energy in oil, with its ability to power heavy equipment and transport goods over long distances, cities have been able to achieve the scale they do today by drawing support from a land base often several hundred times their own area.
Yet these resources are dwindling. Furthermore,
Not only are concentrated raw resources becoming rarer, but previous investments in infrastructure (for example, ports) are in the process of decay and facing accelerating threats from climate change and social disruptions [Bradford 2019: 1].
Thus, “contrary to the forecasts of most demographers, urbanization will reverse course as globalization unwinds during the 21st century” [Bradford 2019: 1].
The report explains that for multiple reasons, renewable energy will not seamlessly or completely be able replace fossil fuel use, in spite of a deep cultural belief in technological progress. And as cities falter and urban food shortages occur, people will be compelled to disperse into the countryside and to develop skills to ensure their food security.
Food, its scarcity, the desire and opportunity to grow it, and the need to do it in ways that are appropriate to place and circumstance, will drive demographic shifts this century. People with life experiences and training aimed at urbanism are going to need a rapid education on what it takes to live off the land, and so-called conventional farmers and ranchers will have a steep learning curve to adopt more frugal and sustainable methods [Bradford 2019: 19].
.
Having established a vision of the unfolding of the 21st Century, dubbed the “Great Simplification” and “characterized by fewer monetary transactions and an increase in subsistence and informal economies,” the author presents alternative agricultural systems, including agroecology, permaculture and holistic management, with potential to overcome the problems created by current farming systems. Included at the margins of the text is key technical information about soil composition, soil types and horizons, and livestock anatomy, as if to get laymen up to speed on agricultural basics for their future rural livelihoods.
In short, the Post Carbon Institute anticipates that resource scarcity will precipitate the collapse and subsequent reorganization of societies, along with their guiding narratives. By necessity, people will learn to consume less and better appreciate our inexorable dependence on the land. Other authors reviewed above suggest the potential to avoid ecological and social collapse by changing the cultural narratives that perpetuate overconsumption and overexploitation of people and nature.
In the U.S., some visions for food system change are anchored in the policy framework of the Ocasio-Cortez/Markley Green New Deal, itself viewed by many as a proposal for transformative change. Noting that the way we eat accounts for a quarter of greenhouse gas emissions and that “the food system is breaking the planet,” Patel and Goodman argue that the Green New Deal could redirect public funds from grain commodities, used largely for processed foods, bioenergy and meats raised in confinement, toward production of healthy foods. More evenly distributed support for greater numbers of farmers could significantly reduce rural poverty while easing pressure on and even regenerating ecosystems.
Yet farmer organizations on the right (Farm Bureau) and left (Farmers Union) have disparaged the Green New Deal, which the authors explain is because the proposal challenges a politically driven cultural view that industrial farming systems are more efficient and thus superior. Thus, transformative change, according to these authors who draw on the original New Deal for guidance, lies in building alliances among farmers, farm workers and consumers, which arecapable of confronting this culturalnarrative.
They suggest that confronting food system monopolies (especially in the meat industry) could bind farmers of all political stripes together, and that the question of food prices could bring farmers and consumers together. “For a rural Green New Deal to work in the 21st Century, everyone’s income needs to increase… Instead of driving down the costs of farming to make food cheap enough for urban workers to buy on stagnating wages, all workers must make enough to afford food that’s sustainably produced.”
In the lead up to the 2020 UN Convention on Biological Diversity, a consortium of conservation groups has also called for 30% both of oceans and 30% of land surface to be conservation protected. Specifically, The United Nations Foundation, Birdlife International, National Geographic and 10 other organizations call for a New Deal for Nature and People to prevent extinctions, reverse the decline of species populations, stabilize natural ecosystems and their services, and restore degraded lands. The call acknowledges the leadership of Indigenous Peoples, who should play a key role in the management of protected areas.
This 2-page vision statement opens with the following:
The Convention on Biological Diversity aims to ensure the conservation, sustainable use and equitable sharing of the benefits of biological diversity. Securing Earth’s biological diversity is a moral obligation. It is also critical in averting catastrophic climate change and ecosystem collapse. Achieving the aims of the Convention on Biological Diversity is integrally linked with tackling climate change and is critical for realizing the Sustainable Development Goals, as a diverse and healthy planet and is the foundation of human health, security, well-being and development.
This paper recommends protecting 30% of Earth’s surface for conservation by 2030 and 50% by 2050. It also proposes building capacity for indigenous and other local peoples to enhance ecosystem integrity and sequester carbon in non-protected lands, halting energy infrastructure projects, and reducing plastics and toxic pollution.
The authors frame a “Global Deal for Nature” (GDN) as complementary to the 2015 Paris Agreement for achieving the internationally recognized goal of limiting global warming to 1.5°C. While the Paris Agreement focuses on critical emissions reductions, the GDN expands the scope of measures needed to include ecosystem and biodiversity conservation. The authors explain that the Paris agreement serves as a strong starting point, but is only a half-deal. It will not alone save the diversity of life on Earth or conserve ecosystem services upon which humanity depends.
Since the crucial role of intact, diverse systems has also been demonstrated to be essential for carbon storage, the GDN will need to emphasize mechanisms for protecting intactness both inside and outside of protected areas … well before 2050 [Dinerstein 2019: 12].
Contextualizing development of the idea of a GDN as part of a greater reassessment of the role of nature in the midst of a planetary emergency, the authors explain that:
The concept of a GDN as a policy mechanism emerged from an earlier study restricted to protecting biodiversity in the terrestrial realm. We expand that perspective to the freshwater and marine realms while simultaneously lending support to an alternative pathway to remaining below 1.5°C that relies heavily on aggressive conservation of remaining habitats. This approach not only safeguards biodiversity but also is the cheapest and fastest alternative for addressing climate change and is not beholden to developing carbon removal technologies unlikely to be effective or to scale in the time-bound nature of the current twin crises [Dinerstein 2019: 1].
The authors recommend that a GDN prioritize: (1) protecting biodiversity, (2) mitigating climate change, and (3) reducing threats to ecosystem intactness and persistence of species. The strategy for the first priority – protecting biodiversity – is to expand the percentage of Earth’s surface that is conservation protected to 30% by 2030, aiming for 50% protection by 2050. Currently, less than 15% of land is protected, and only half of currently protected land is connected by ecological corridors to facilitate animal migrations, while only 2-4% of the world’s ocean area is protected. To avoid the risks of (a) prioritizing low-biodiversity sites at the expense of biodiversity hotspots, or (b) an uneven representation of ecoregion types achieving protected status, the authors organized conservation targets according to 846 terrestrial ecoregions.
For the second priority – mitigating climate change – the authors propose that land outside conservation protected areas be managed in a way that maintains ecosystem intactness, prevents emissions and sequesters carbon. This would include, for example, indigenous lands, where people often lack tenure rights. Ensuring secure land tenure to indigenous people would allow them to continue managing land in a way that supports the vitality of the ecosystems on which they depend.
The third priority – reducing major threats – would involve scrutinizing or halting new infrastructure projects (especially agricultural land expansion, road construction and energy development) on natural lands. In addition, it would involve reducing hunting and poaching, as well as the production and use of plastics and toxins.
This paper calls for efforts to make farmers, land managers, policy makers, and the public at large keenly aware of the link between soil carbon and its more widely appreciated social outcomes, such as agricultural productivity and food security, improved water quality, flood and drought mitigation, lower rates of migration, biodiversity preservation, and climate change mitigation.
The authors identify three priorities for global action to build soil carbon stocks: (1) build an overarching case and vision for action, led by political champions; (2) build a stronger business case and track-record of success among public and private investors; and (3) establish a more compelling value proposition for farmers and land managers.
Specifically, Vermeulen et al. propose that champions in countries already prioritizing soil carbon in national policy lead efforts to generate greater awareness, such as through
persuasive narratives and campaigns [that] might link soil health and carbon storage to broader societal outcomes with wider political traction. These include double-digit increases in yield potential, particularly on degraded lands, higher household and national food security, reduced risks from disasters, improved water quality and lower rates of displacement and migration [Vermeulen 2019: 3].
They also propose that soil carbon take center stage in discourses on sustainable agriculture, from which it has largely been absent.
To build a business case among potential investors in soil health, the authors suggest, for example, creating “small-scale funds to flow to commercial demonstrations of soil organic carbon that can then be ready for widespread proliferation” [Vermeulen 2019: 3].
The authors stress the importance of demonstrating to farmers the multi-faceted long-term value achieved by incorporating soil-building practices into day-to-day farming operations, including: (1) enhanced productivity, (2) improved risk management (for example, resilience to drought), (3) superior market access (for example, certified value chains), (4) financial returns to carbon assets, and (5) government support (for example, environmental subsidies).
Another priority is “to move beyond stand-alone protocols by building soil organic carbon into existing frameworks from which it is absent, such as UNCCD, UNFCCC, Ramsar and the Global Reporting Initiative” [Vermeulen 2019: 4]. In other words, given the foundational role of soils in ecosystem function (and thus the delivery of vital ecosystem services), improving soil health must be treated as the powerful leverage point that it is for resolving multiple overlapping crises.
This paper calls for efforts to make farmers, land managers, policy makers, and the public at large keenly aware of the link between soil carbon and its more widely appreciated social outcomes, such as agricultural productivity and food security, improved water quality, flood and drought mitigation, lower rates of migration, biodiversity preservation, and climate change mitigation.
The authors identify three priorities for global action to build soil carbon stocks: (1) build an overarching case and vision for action, led by political champions; (2) build a stronger business case and track-record of success among public and private investors; and (3) establish a more compelling value proposition for farmers and land managers.
Specifically, Vermeulen et al. propose that champions in countries already prioritizing soil carbon in national policy lead efforts to generate greater awareness, such as through
persuasive narratives and campaigns [that] might link soil health and carbon storage to broader societal outcomes with wider political traction. These include double-digit increases in yield potential, particularly on degraded lands, higher household and national food security, reduced risks from disasters, improved water quality and lower rates of displacement and migration [Vermeulen 2019: 3].
They also propose that soil carbon take center stage in discourses on sustainable agriculture, from which it has largely been absent.
To build a business case among potential investors in soil health, the authors suggest, for example, creating “small-scale funds to flow to commercial demonstrations of soil organic carbon that can then be ready for widespread proliferation” [Vermeulen 2019: 3].
The authors stress the importance of demonstrating to farmers the multi-faceted long-term value achieved by incorporating soil-building practices into day-to-day farming operations, including: (1) enhanced productivity, (2) improved risk management (for example, resilience to drought), (3) superior market access (for example, certified value chains), (4) financial returns to carbon assets, and (5) government support (for example, environmental subsidies).
Another priority is “to move beyond stand-alone protocols by building soil organic carbon into existing frameworks from which it is absent, such as UNCCD, UNFCCC, Ramsar and the Global Reporting Initiative” [Vermeulen 2019: 4]. In other words, given the foundational role of soils in ecosystem function (and thus the delivery of vital ecosystem services), improving soil health must be treated as the powerful leverage point that it is for resolving multiple overlapping crises.
A global deal for nature, Dinerstein 2019
This paper recommends protecting 30% of Earth’s surface for conservation by 2030 and 50% by 2050. It also proposes building capacity for indigenous and other local peoples to enhance ecosystem integrity and sequester carbon in non-protected lands, halting energy infrastructure projects, and reducing plastics and toxic pollution.
The authors frame a “Global Deal for Nature” (GDN) as complementary to the 2015 Paris Agreement for achieving the internationally recognized goal of limiting global warming to 1.5°C. While the Paris Agreement focuses on critical emissions reductions, the GDN expands the scope of measures needed to include ecosystem and biodiversity conservation. The authors explain that the Paris agreement serves as a strong starting point, but is only a half-deal. It will not alone save the diversity of life on Earth or conserve ecosystem services upon which humanity depends.
Since the crucial role of intact, diverse systems has also been demonstrated to be essential for carbon storage, the GDN will need to emphasize mechanisms for protecting intactness both inside and outside of protected areas … well before 2050 [Dinerstein 2019: 12].
Contextualizing development of the idea of a GDN as part of a greater reassessment of the role of nature in the midst of a planetary emergency, the authors explain that:
The concept of a GDN as a policy mechanism emerged from an earlier study restricted to protecting biodiversity in the terrestrial realm. We expand that perspective to the freshwater and marine realms while simultaneously lending support to an alternative pathway to remaining below 1.5°C that relies heavily on aggressive conservation of remaining habitats. This approach not only safeguards biodiversity but also is the cheapest and fastest alternative for addressing climate change and is not beholden to developing carbon removal technologies unlikely to be effective or to scale in the time-bound nature of the current twin crises [Dinerstein 2019: 1].
The authors recommend that a GDN prioritize: (1) protecting biodiversity, (2) mitigating climate change, and (3) reducing threats to ecosystem intactness and persistence of species. The strategy for the first priority – protecting biodiversity – is to expand the percentage of Earth’s surface that is conservation protected to 30% by 2030, aiming for 50% protection by 2050. Currently, less than 15% of land is protected, and only half of currently protected land is connected by ecological corridors to facilitate animal migrations, while only 2-4% of the world’s ocean area is protected. To avoid the risks of (a) prioritizing low-biodiversity sites at the expense of biodiversity hotspots, or (b) an uneven representation of ecoregion types achieving protected status, the authors organized conservation targets according to 846 terrestrial ecoregions.
For the second priority – mitigating climate change – the authors propose that land outside conservation protected areas be managed in a way that maintains ecosystem intactness, prevents emissions and sequesters carbon. This would include, for example, indigenous lands, where people often lack tenure rights. Ensuring secure land tenure to indigenous people would allow them to continue managing land in a way that supports the vitality of the ecosystems on which they depend.
The third priority – reducing major threats – would involve scrutinizing or halting new infrastructure projects (especially agricultural land expansion, road construction and energy development) on natural lands. In addition, it would involve reducing hunting and poaching, as well as the production and use of plastics and toxins.
Joint statement on post-2020 global biodiversity framework 2050 Convention on Biological Diversity vision: “Living in Harmony with Nature,” Birdlife International et al.
In the lead up to the 2020 UN Convention on Biological Diversity, a consortium of conservation groups has also called for 30% both of oceans and 30% of land surface to be conservation protected. Specifically, The United Nations Foundation, Birdlife International, National Geographic and 10 other organizations call for a New Deal for Nature and People to prevent extinctions, reverse the decline of species populations, stabilize natural ecosystems and their services, and restore degraded lands. The call acknowledges the leadership of Indigenous Peoples, who should play a key role in the management of protected areas.
This 2-page vision statement opens with the following:
The Convention on Biological Diversity aims to ensure the conservation, sustainable use and equitable sharing of the benefits of biological diversity. Securing Earth’s biological diversity is a moral obligation. It is also critical in averting catastrophic climate change and ecosystem collapse. Achieving the aims of the Convention on Biological Diversity is integrally linked with tackling climate change and is critical for realizing the Sustainable Development Goals, as a diverse and healthy planet and is the foundation of human health, security, well-being and development.
A Green New Deal for Agriculture, Patel & Goodman 2019
In the U.S., some visions for food system change are anchored in the policy framework of the Ocasio-Cortez/Markley Green New Deal, itself viewed by many as a proposal for transformative change. Noting that the way we eat accounts for a quarter of greenhouse gas emissions and that “the food system is breaking the planet,” Patel and Goodman argue that the Green New Deal could redirect public funds from grain commodities, used largely for processed foods, bioenergy and meats raised in confinement, toward production of healthy foods. More evenly distributed support for greater numbers of farmers could significantly reduce rural poverty while easing pressure on and even regenerating ecosystems.
Yet farmer organizations on the right (Farm Bureau) and left (Farmers Union) have disparaged the Green New Deal, which the authors explain is because the proposal challenges a politically driven cultural view that industrial farming systems are more efficient and thus superior. Thus, transformative change, according to these authors who draw on the original New Deal for guidance, lies in building alliances among farmers, farm workers and consumers, which arecapable of confronting this culturalnarrative.
They suggest that confronting food system monopolies (especially in the meat industry) could bind farmers of all political stripes together, and that the question of food prices could bring farmers and consumers together. “For a rural Green New Deal to work in the 21st Century, everyone’s income needs to increase… Instead of driving down the costs of farming to make food cheap enough for urban workers to buy on stagnating wages, all workers must make enough to afford food that’s sustainably produced.”
The future is rural, Bradford 2019
Taking an altogether different angle, Jason Bradford of the Post Carbon Institute assumes radical societal change is inevitable and imminent, and focuses not on how to precipitate change but instead on how to adapt to it. “The future is rural” [Bradford 2019] is essentially a primer on how to navigate the profound changes society will undergo during the 21st Century due to climate breakdown and resource scarcity. It begins with an assertion that today’s “mass urbanization has been made possible by the prodigious exploitation of fossil fuels.” In other words,
Due to the concentrated energy in oil, with its ability to power heavy equipment and transport goods over long distances, cities have been able to achieve the scale they do today by drawing support from a land base often several hundred times their own area.
Yet these resources are dwindling. Furthermore,
Not only are concentrated raw resources becoming rarer, but previous investments in infrastructure (for example, ports) are in the process of decay and facing accelerating threats from climate change and social disruptions [Bradford 2019: 1].
Thus, “contrary to the forecasts of most demographers, urbanization will reverse course as globalization unwinds during the 21st century” [Bradford 2019: 1].
The report explains that for multiple reasons, renewable energy will not seamlessly or completely be able replace fossil fuel use, in spite of a deep cultural belief in technological progress. And as cities falter and urban food shortages occur, people will be compelled to disperse into the countryside and to develop skills to ensure their food security.
Food, its scarcity, the desire and opportunity to grow it, and the need to do it in ways that are appropriate to place and circumstance, will drive demographic shifts this century. People with life experiences and training aimed at urbanism are going to need a rapid education on what it takes to live off the land, and so-called conventional farmers and ranchers will have a steep learning curve to adopt more frugal and sustainable methods [Bradford 2019: 19].
.
Having established a vision of the unfolding of the 21st Century, dubbed the “Great Simplification” and “characterized by fewer monetary transactions and an increase in subsistence and informal economies,” the author presents alternative agricultural systems, including agroecology, permaculture and holistic management, with potential to overcome the problems created by current farming systems. Included at the margins of the text is key technical information about soil composition, soil types and horizons, and livestock anatomy, as if to get laymen up to speed on agricultural basics for their future rural livelihoods.
In short, the Post Carbon Institute anticipates that resource scarcity will precipitate the collapse and subsequent reorganization of societies, along with their guiding narratives. By necessity, people will learn to consume less and better appreciate our inexorable dependence on the land. Other authors reviewed above suggest the potential to avoid ecological and social collapse by changing the cultural narratives that perpetuate overconsumption and overexploitation of people and nature.
One million of an estimated 8 million species on Earth are at risk of extinction in the coming decades, according to a May 2019 report from the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES). Children today will live as adults in a world without the Milky Stork, without the Caquetá Tití Monkey, and without the Thongaree’s Disc-nosed Bat, and more generally without 40% of today’s amphibian species and with a third fewer shark and reef-forming coral species.
It’s not merely a sentimental loss of charming creatures we face, but an unravelling of ecosystems, which are knit together by biodiversity, and the accompanying loss of the life-sustaining services nature provides to humans. This was the message of the report’s 400-plus authors, who emphasized that biodiversity collapse is comparable to and intertwined with climate change in scale and severity.
Nature, through its ecological and evolutionary processes, sustains the quality of the air, fresh water and soils on which humanity depends, distributes fresh water, regulates the climate, provides pollination and pest control and reduces the impact of natural hazards. For example, more than 75 percent of global food crop types, including fruits and vegetables and some of the most important cash crops such as coffee, cocoa and almonds, rely on animal pollination. Marine and terrestrial ecosystems are the sole sinks for anthropogenic carbon emissions, with a gross sequestration of 5.6 gigatons of carbon per year (the equivalent of some 60 percent of global anthropogenic emissions). Nature underpins all dimensions of human health and contributes to non-material aspects of quality of life – inspiration and learning, physical and psychological experiences, and supporting identities – that are central to quality of life and cultural integrity, even if their aggregated value is difficult to quantify [Diaz 2019: 2].
The report warns that ecosystem deterioration is global and that its direct drivers – pollution, climate change, land and sea-use change, direct exploitation, and invasive species invasions – are accelerating. “Most international societal and environmental goals, such as those embodied in the Aichi Biodiversity Targets and the 2030 Agenda for Sustainable Development, will not be achieved based on current trajectories” [Diaz 2019: 5]. Reversing course from the catastrophic outcomes realizable throughout the 21st Century is possible, according to the report, but only through “transformative change” – in other words “fundamental, system-wide reorganization across technological, economic and social factors, including paradigms, goals and values” [Diaz 2019: 21].
Acting immediately and simultaneously on multiple indirect and direct drivers has the potential to slow, halt and even reverse some aspects of biodiversity and ecosystem loss [Diaz 2019: 7].
Change is transformative when aimed at the underlying drivers of a system. As IPBES explains, indirect drivers of ecosystem deterioration are the institutions that govern social relations including, for example, systems of property rights, governance systems, treaties, and informal social norms and rules. Institutions are designed to produce certain beneficial outcomes, yet in the process also produce predictable negative outcomes.
Since current structures often inhibit sustainable development and actually represent the indirect drivers of biodiversity loss, such fundamental, structural change is called for. By its very nature, transformative change can expect opposition from those with interests vested in the status quo, but such opposition can be overcome for the broader public good. If obstacles are overcome, commitment to mutually supportive international goals and targets, supporting actions by indigenous peoples and local communities at the local level, new frameworks for private sector investment and innovation, inclusive and adaptive governance approaches and arrangements, multi-sectoral planning and strategic policy mixes can help to transform the public and private sectors to achieve sustainability at the local, national and global levels [Diaz 2019: 7].
The transformative change envisioned here is a world where local and national governments are inclusive and internationally accountable. In other words, our societies are governed by institutions disposed to an equitable sharing of the responsibilities and benefits of global citizenship, and they act in service not only to the powerful, but are also responsive and supportive toward those without money or power. Capacity building is focused on local communities, where policy changes become manifest.
How to realize such change? IPBES suggests several leverage points, where efforts made to create a sustainable society can have a disproportionate effect. Cultural or value-system leverage points include “enabling visions of a good quality of life that do not entail ever-increasing material consumption” [Diaz 2019: 8] expanding a widely held value of responsibility to include the impacts of consumption; addressing social inequalities; ensuring inclusive decision-making; and promoting education and the maintenance of diverse knowledge systems, including scientific, local and indigenous knowledge.
Specific actions recommended include: promoting sustainable/agroecological farming and ecosystem-based fishing practices, and the development of urban green infrastructure to bolster climate change mitigation and adaptation. It also recommends fostering an evolution in economic/financial systems to reduce inequality and overconsumption and to steer away from the paradigm of economic growth.
Emerging in tandem with the IPBES assessment are a diversity of complementary proposals/analyses, summarized below, which articulate pathways forward deemed transformative enough to limit the severity of and/or adapt to climate breakdown.
Overall, these papers point to the importance of changing the cultural narratives that guide human behavior. Education that draws onscientific and indigenous knowledge, local capacity building and alliance building across political divisions are offered as levers with the potential to change societies’ dominant narratives. An emergent cultural narrative might recognize biologically active, carbon-rich soil as a firmer basis than i-phones, plastic water bottles and the like for the provision of human safety, pleasure and happiness in life. And with widespread adoption of such a narrative, the public might be galvanized to win policy changes robust enough to reverse course from catastrophic outcomes.
An emergent cultural narrative might recognize biologically active, carbon-rich soil as a firmer basis than i-phones, plastic water bottles and the like for the provision of human safety, pleasure and happiness in life.
Compilation of article summaries envisioning societal change
A global agenda for soil carbon, Vermeulen 2019
This paper calls for efforts to make farmers, land managers, policy makers, and the public at large keenly aware of the link between soil carbon and its more widely appreciated social outcomes, such as agricultural productivity and food security, improved water quality, flood and drought mitigation, lower rates of migration, biodiversity preservation, and climate change mitigation.
The authors identify three priorities for global action to build soil carbon stocks: (1) build an overarching case and vision for action, led by political champions; (2) build a stronger business case and track-record of success among public and private investors; and (3) establish a more compelling value proposition for farmers and land managers.
Specifically, Vermeulen et al. propose that champions in countries already prioritizing soil carbon in national policy lead efforts to generate greater awareness, such as through
persuasive narratives and campaigns [that] might link soil health and carbon storage to broader societal outcomes with wider political traction. These include double-digit increases in yield potential, particularly on degraded lands, higher household and national food security, reduced risks from disasters, improved water quality and lower rates of displacement and migration [Vermeulen 2019: 3].
They also propose that soil carbon take center stage in discourses on sustainable agriculture, from which it has largely been absent.
To build a business case among potential investors in soil health, the authors suggest, for example, creating “small-scale funds to flow to commercial demonstrations of soil organic carbon that can then be ready for widespread proliferation” [Vermeulen 2019: 3].
The authors stress the importance of demonstrating to farmers the multi-faceted long-term value achieved by incorporating soil-building practices into day-to-day farming operations, including: (1) enhanced productivity, (2) improved risk management (for example, resilience to drought), (3) superior market access (for example, certified value chains), (4) financial returns to carbon assets, and (5) government support (for example, environmental subsidies).
Another priority is “to move beyond stand-alone protocols by building soil organic carbon into existing frameworks from which it is absent, such as UNCCD, UNFCCC, Ramsar and the Global Reporting Initiative” [Vermeulen 2019: 4]. In other words, given the foundational role of soils in ecosystem function (and thus the delivery of vital ecosystem services), improving soil health must be treated as the powerful leverage point that it is for resolving multiple overlapping crises.
A global deal for nature, Dinerstein 2019
This paper recommends protecting 30% of Earth’s surface for conservation by 2030 and 50% by 2050. It also proposes building capacity for indigenous and other local peoples to enhance ecosystem integrity and sequester carbon in non-protected lands, halting energy infrastructure projects, and reducing plastics and toxic pollution.
The authors frame a “Global Deal for Nature” (GDN) as complementary to the 2015 Paris Agreement for achieving the internationally recognized goal of limiting global warming to 1.5°C. While the Paris Agreement focuses on critical emissions reductions, the GDN expands the scope of measures needed to include ecosystem and biodiversity conservation. The authors explain that the Paris agreement serves as a strong starting point, but is only a half-deal. It will not alone save the diversity of life on Earth or conserve ecosystem services upon which humanity depends.
Since the crucial role of intact, diverse systems has also been demonstrated to be essential for carbon storage, the GDN will need to emphasize mechanisms for protecting intactness both inside and outside of protected areas … well before 2050 [Dinerstein 2019: 12].
Contextualizing development of the idea of a GDN as part of a greater reassessment of the role of nature in the midst of a planetary emergency, the authors explain that:
The concept of a GDN as a policy mechanism emerged from an earlier study restricted to protecting biodiversity in the terrestrial realm. We expand that perspective to the freshwater and marine realms while simultaneously lending support to an alternative pathway to remaining below 1.5°C that relies heavily on aggressive conservation of remaining habitats. This approach not only safeguards biodiversity but also is the cheapest and fastest alternative for addressing climate change and is not beholden to developing carbon removal technologies unlikely to be effective or to scale in the time-bound nature of the current twin crises [Dinerstein 2019: 1].
The authors recommend that a GDN prioritize: (1) protecting biodiversity, (2) mitigating climate change, and (3) reducing threats to ecosystem intactness and persistence of species. The strategy for the first priority – protecting biodiversity – is to expand the percentage of Earth’s surface that is conservation protected to 30% by 2030, aiming for 50% protection by 2050. Currently, less than 15% of land is protected, and only half of currently protected land is connected by ecological corridors to facilitate animal migrations, while only 2-4% of the world’s ocean area is protected. To avoid the risks of (a) prioritizing low-biodiversity sites at the expense of biodiversity hotspots, or (b) an uneven representation of ecoregion types achieving protected status, the authors organized conservation targets according to 846 terrestrial ecoregions.
For the second priority – mitigating climate change – the authors propose that land outside conservation protected areas be managed in a way that maintains ecosystem intactness, prevents emissions and sequesters carbon. This would include, for example, indigenous lands, where people often lack tenure rights. Ensuring secure land tenure to indigenous people would allow them to continue managing land in a way that supports the vitality of the ecosystems on which they depend.
The third priority – reducing major threats – would involve scrutinizing or halting new infrastructure projects (especially agricultural land expansion, road construction and energy development) on natural lands. In addition, it would involve reducing hunting and poaching, as well as the production and use of plastics and toxins.
Joint statement on post-2020 global biodiversity framework 2050 Convention on Biological Diversity vision: “Living in Harmony with Nature,” Birdlife International et al.
In the lead up to the 2020 UN Convention on Biological Diversity, a consortium of conservation groups has also called for 30% both of oceans and 30% of land surface to be conservation protected. Specifically, The United Nations Foundation, Birdlife International, National Geographic and 10 other organizations call for a New Deal for Nature and People to prevent extinctions, reverse the decline of species populations, stabilize natural ecosystems and their services, and restore degraded lands. The call acknowledges the leadership of Indigenous Peoples, who should play a key role in the management of protected areas.
This 2-page vision statement opens with the following:
The Convention on Biological Diversity aims to ensure the conservation, sustainable use and equitable sharing of the benefits of biological diversity. Securing Earth’s biological diversity is a moral obligation. It is also critical in averting catastrophic climate change and ecosystem collapse. Achieving the aims of the Convention on Biological Diversity is integrally linked with tackling climate change and is critical for realizing the Sustainable Development Goals, as a diverse and healthy planet and is the foundation of human health, security, well-being and development.
A Green New Deal for Agriculture, Patel & Goodman 2019
In the U.S., some visions for food system change are anchored in the policy framework of the Ocasio-Cortez/Markley Green New Deal, itself viewed by many as a proposal for transformative change. Noting that the way we eat accounts for a quarter of greenhouse gas emissions and that “the food system is breaking the planet,” Patel and Goodman argue that the Green New Deal could redirect public funds from grain commodities, used largely for processed foods, bioenergy and meats raised in confinement, toward production of healthy foods. More evenly distributed support for greater numbers of farmers could significantly reduce rural poverty while easing pressure on and even regenerating ecosystems.
Yet farmer organizations on the right (Farm Bureau) and left (Farmers Union) have disparaged the Green New Deal, which the authors explain is because the proposal challenges a politically driven cultural view that industrial farming systems are more efficient and thus superior. Thus, transformative change, according to these authors who draw on the original New Deal for guidance, lies in building alliances among farmers, farm workers and consumers, which arecapable of confronting this culturalnarrative.
They suggest that confronting food system monopolies (especially in the meat industry) could bind farmers of all political stripes together, and that the question of food prices could bring farmers and consumers together. “For a rural Green New Deal to work in the 21st Century, everyone’s income needs to increase… Instead of driving down the costs of farming to make food cheap enough for urban workers to buy on stagnating wages, all workers must make enough to afford food that’s sustainably produced.”
The future is rural, Bradford 2019
Taking an altogether different angle, Jason Bradford of the Post Carbon Institute assumes radical societal change is inevitable and imminent, and focuses not on how to precipitate change but instead on how to adapt to it. “The future is rural” [Bradford 2019] is essentially a primer on how to navigate the profound changes society will undergo during the 21st Century due to climate breakdown and resource scarcity. It begins with an assertion that today’s “mass urbanization has been made possible by the prodigious exploitation of fossil fuels.” In other words,
Due to the concentrated energy in oil, with its ability to power heavy equipment and transport goods over long distances, cities have been able to achieve the scale they do today by drawing support from a land base often several hundred times their own area.
Yet these resources are dwindling. Furthermore,
Not only are concentrated raw resources becoming rarer, but previous investments in infrastructure (for example, ports) are in the process of decay and facing accelerating threats from climate change and social disruptions [Bradford 2019: 1].
Thus, “contrary to the forecasts of most demographers, urbanization will reverse course as globalization unwinds during the 21st century” [Bradford 2019: 1].
The report explains that for multiple reasons, renewable energy will not seamlessly or completely be able replace fossil fuel use, in spite of a deep cultural belief in technological progress. And as cities falter and urban food shortages occur, people will be compelled to disperse into the countryside and to develop skills to ensure their food security.
Food, its scarcity, the desire and opportunity to grow it, and the need to do it in ways that are appropriate to place and circumstance, will drive demographic shifts this century. People with life experiences and training aimed at urbanism are going to need a rapid education on what it takes to live off the land, and so-called conventional farmers and ranchers will have a steep learning curve to adopt more frugal and sustainable methods [Bradford 2019: 19].
.
Having established a vision of the unfolding of the 21st Century, dubbed the “Great Simplification” and “characterized by fewer monetary transactions and an increase in subsistence and informal economies,” the author presents alternative agricultural systems, including agroecology, permaculture and holistic management, with potential to overcome the problems created by current farming systems. Included at the margins of the text is key technical information about soil composition, soil types and horizons, and livestock anatomy, as if to get laymen up to speed on agricultural basics for their future rural livelihoods.
In short, the Post Carbon Institute anticipates that resource scarcity will precipitate the collapse and subsequent reorganization of societies, along with their guiding narratives. By necessity, people will learn to consume less and better appreciate our inexorable dependence on the land. Other authors reviewed above suggest the potential to avoid ecological and social collapse by changing the cultural narratives that perpetuate overconsumption and overexploitation of people and nature.
Stories of blessed unrest
The following sketches are but a tiny sampling of the countless ways people throughout the world push back against the socio-economic and political forces of destruction both of ecosystems and of the social fabric of society. Adopting Paul Hawken’s terminology and characterization of “blessed unrest” as a spontaneous, decentralized global social movement, we here present a diverse though far from representative series of vignettes of everyday heroes. May such stories light the fire for new heroes to perpetually emerge in defense of all life on Earth.
Minibigforest in Nantes
Hearing of plans underway for a four-lane highway near their home in Nantes, France, local residents Jim and Stephanie responded by planting a small forest. The idea was not only to block out the added sound and air pollution, but also to try to compensate for the assault on the planet of any road expansion. The couple was inspired by Shubhendhu Sharma, who spoke at the 2018 Nantes festival Aux Arbes[8]. Sharma showed the audience how 300 trees of 30-some species could be planted in the space of six parking places. He described the Miyawaki Method, which mimics natural forests in terms of biodiversity and density, outperforms the growth rate of monoculture plantations tenfold, and works well in urban areas because it takes so little space. Within the following year, Jim and Stephanie, along with dozens of volunteers and school kids, planted more than 2000 trees on two sites. To encourage Miyawaki-style afforestation projects everywhere, the couple launched the initiative Minibigforest; this is only the beginning for them.
Greta Thunberg and a million international student strikers
At the age of 15, Greta Thunberg began sitting on the steps of the Swedish parliament with a handmade sign reading: “skolstrejk för klimatet” or “school strike for the climate.” The decision to act came about seven years after she first learned of climate change. The fact that adults didn’t seem bothered to do anything about the global crisis shocked her, and then sent her into a depression. Activism pulled her out of depression and thrust her onto the international stage. It didn’t take long for her solo picketing efforts to spark a global movement spanning 125 countries of more than a million kids striking from school for climate. Greta intends to continue striking outside the Swedish Parliament until it passes legislation that upholds commitments made in the Paris Climate Accord.
Excerpted from a Guardian guest editorial by climate strikers Greta Thunberg (Sweden), Anna Taylor (UK), Luisa Neubauer (Germany), Kyra Gantois, Anuna De Wever and Adélaïde Charlier (Belgium), Holly Gillibrand (Scotland), and Alexandria Villasenor (USA):
This movement had to happen, we didn’t have a choice. The vast majority of climate strikers taking action today aren’t allowed to vote. Imagine for a second what that feels like. Despite watching the climate crisis unfold, despite knowing the facts, we aren’t allowed to have a say in who makes the decisions about climate change. And then ask yourself this: wouldn’t you go on strike too, if you thought doing so could help protect your own future?
So today we walk out of school, we quit our college lessons, and we take to the streets to say enough is enough. Some adults say we shouldn’t be walking out of classes – that we should be “getting an education”. We think organising against an existential threat – and figuring out how to make our voices heard – is teaching us some important lessons.
The Waorani people stand up for their rainforest homeland
When the Waorani people of the Ecuadorian Amazon heard their government was planning to sell drilling rights to their land to international oil companies, they mobilized. They mapped the land to illustrate to the Western world its otherwise unseen cultural, historical and ecological richness. These maps include “historic battle sites, ancient cave-carvings, jaguar trails, medicinal plants, animal reproductive zones, important fishing holes, creek-crossings and sacred waterfalls,” according to an online petition they launched in partnership with the NGO Amazon Frontlines. Then the Waorani sued the government for not properly consulting them when the decision was made in 2012 to dice up the rainforest into auctionable blocks of land. In April 2019, the Ecuadorian court ruled in favor of the Waorani, immediately suspending any sale of the land and setting a precedent for other communities resisting oil extraction in their lands.
The government’s interests in oil is not more valuable than our rights, our forests, our lives.
– Nemonte Nenquimo, one of the Waorani plaintiffs and representative of the Coordinating Council of the Waorani Nationality Ecuador Pastaza (CONCONAWEP).
The Waorani win follows a win against mining operations last year by the indigenous Kofan community also in the Ecuadorian Amazon.
On the other side of the Amazon and across the South Atlantic Ocean, the small South African community of Xolobeni won a similar court case. Like the Waorani, the people of Xolobeni demanded that they be consulted rather than being forced to cede their land to mining interests – in this case to an Australian titanium mining company. Also like the Waorani, they were defending not only their lives, livelihoods, their health and wellbeing, but also an ecologically rich corner of the planet. Xolobeni is in Pondoland, a dune-covered stretch of the coast that is home to endemic species and frequented offshore by whales.
The law says we have a right to be consulted, but what we say doesn’t seem to matter. We have told the company many times that we don’t want their mine. How many times do we have to say no?
– Nonhle Mbuthuma, local resident
The court agreed that local communities must give their consent before mining is allowed on their land.
Methow Beaver Project:enlisting beavers to make wetlands in compensation for declining mountain snowpack
The deep winter snow falls on the mountains around the Methow Valley in the state of Washington are declining. To manage problems with drought, the Methow Beaver Project has been capturing, tagging, matching male and female beavers and releasing them in key valley areas. The project workers know beavers are master engineers that know how to preserve their homes and food supply, to the benefit of water quality and many other animals and plants in the area. Beaver reintroduction projects are also underway in Nevada, Utah and Wyoming. In 2018, Scott Helker, a Libertarian candidate running to become governor of Colorado, was asked, “Would you support asking Coloradans to raise billions of tax dollars for projects that would increase water supplies and help prevent a projected water shortage mid-century?” He answered, “No. I can create the same results without raising billions of dollars. Ask me how.” Answer: with beaver reintroduction projects.
iMatter is a tight-knit national group of passionate pre-college individuals who are making real impacts in their communities. They are showing up in city halls and state offices, demanding their elected officials at every level possible commit to bold and visionary climate action. Students from Brookline High School in Massachusetts submitted resolutions to their town legislators, saying they’re worried about their future and the future of the environment; their cities agreed and are supporting the Green New Deal. Alec Loorz started the organization Kids vs. Global Warming in California with his mom, Victoria Loorz in 2007, when he was 13 years old. The organization eventually changed its name to iMatter. Alec went on to spearhead the Our Children’s Trust lawsuit against federal and state governments of the United States to secure climate recovery plans that will restore the balance of Earth’s climate systems.
This paper analyzes 49 studies (1515 landscapes encompassing both organic and conventional agricultural production) in Europe to determine “effects of landscape composition (% habitats) and configuration (edge density) on arthropods[7] in fields and their margins, pest control, pollination and yield” [Martin 2019: 1].
Edge density is measured as the length of edge per area of land. Edges between adjacent crop fields and between crop fields and semi-natural areas such as grasslands or other land patches not used for crops allow for “exchange between landscape patches” [Martin 2019: 4] for pollinators, pest predators and other providers of ecosystem services. High edge density is associated with smaller field size, and lower edge density with larger field size.
Complex landscapes where small and/or irregularly shaped fields and habitat patches prevail have a high density of edges. Due to increased opportunities for exchange, these landscapes are likely to support spillover of dispersal-limited populations between patches [Martin 2019: 3].
Researchers found that:
In landscapes with high edge density, 70% of pollinator and 44% of natural enemy species reached highest abundances and pollination and pest control improved 1.7- and 1.4-fold, respectively. Arable-dominated landscapes with high edge densities achieved high yields. This suggests that enhancing edge density in European agroecosystems can promote functional biodiversity and yield-enhancing ecosystem services [Martin 2019: 1].
Just as high edge density is shown here to maintain yield, low edge density, especially when combined with a lower amount of surrounding semi-natural habitat, can reduce yield.
Reduced pollination and pest control at low edge density may have been compensated by external inputs in productive landscapes. … Intermediate to low yields in landscapes with high % arable, low % semi-natural habitat and low edge density may underpin the risks of ongoing conventional intensification resulting in yield stagnation or reduction despite high agricultural inputs [Martin 2019: 9].
This article illustrates the important role of ecosystem services in maintaining crop yield, as well as the relatively simple management decisions farmers can make to enhance the habitat of arthropods providing those services.
Citing the problems posed globally by pesticide use and farmland expansion, this study looks at the potential of organic farming, seen as a popular prototype of ecological intensification, to limit pest infestations. Ecological intensification “is based on optimizing the ecological functions that support ecosystem services to increase the productivity of agro-ecosystems” [Muneret 2018: 361], and thus serves as a framework for evaluating farming system changes that could handle both ecological stress/collapse and human population growth. “Organic farming is a certified production system based on the principle of using farming practices that are expected to enhance ecological processes while prohibiting the use of external synthetic inputs” [Muneret 2018: 361].
Our findings in particular show that organic farming practices are able to match or outperform conventional pest control practices against pathogens and animal pests [such as insects] whereas weeds are much more abundant in organic than in conventional systems. Thus, ecological intensification based on organic farming can contribute to the control of animal pests and pathogens by enhancing biological control services and limiting their infestation levels [Muneret 2018: 365].
Whereas conventional pest control emphasizes top-down control with pesticide, ecological pest control is achieved through multiple processes:
Once established, pest populations within agro-ecosystems are affected, to varying degrees, by three ecological processes: bottom-up effects mediated by soil or plant communities involving, for instance, plant quality or habitat structure, horizontal processes within a given trophic level such as competition for resources between individuals or populations, and top-down control by natural antagonists such as predation or parasitism [Muneret 2018: 363].
Given the benefits of biodiversity for enhancing these three ecological effects, the authors explain that weeds, which are not as well suppressed in organic systems, may actually be beneficial in terms of limiting infestation by animals/insects and pathogens.
Our analysis shows that organic farming results in much higher weed infestation. This result is supported by previous studies that have shown higher abundance and diversity of plant communities within organic arable fields. We assume that this higher weed infestation, in turn, most likely influences animal pest and pathogen populations. These bottom-up effects of plant communities on higher trophic levels have been demonstrated and more abundant or diverse plant communities have been found to limit insect and disease infestation through direct and indirect mechanisms because of higher structural complexity or lower habitat quality under increased plant diversity. Although this needs further investigation, the observed performance of organic farming on animal pest and pathogen infestation may result from bottom-up effects generated by the higher weed infestation levels in organic cropping systems [Muneret 2018: 364].
Although the authors didn’t examine the effects of pest infestations on yield, they note that previous studies have suggested that weeds do not necessarily result in crop yield reductions in organic systems.
Although this needs further investigation, the observed performance of organic farming on animal pest and pathogen infestation may result from bottom-up effects generated by the higher weed infestation levels in organic cropping systems [Muneret 2018: 364].
This review examines the concept of ecological intensification as a way to increase global food production by enhancing the ecological functionality of farmland.
We present ecological intensification as an alternative approach for mainstream agriculture to meet [future climatic, economic and social] challenges. Ecological intensification aims to match or augment yield levels while minimizing negative impacts on the environment and ensuing negative feedbacks on agricultural productivity, by integrating the management of ecosystem services delivered by biodiversity into crop production systems [Bommarco 2013: 230].
The idea of ecological intensification stems from the concept of ecosystem services, which refers to the benefits humans derive from ecosystems. These services are grouped into four types: supporting (such as soil formation by microorganisms), regulating (such as pest control, crop pollination, climate regulation and water purification), provisioning (such as food, fiber, fuel and water) and cultural (such as education, recreation and aesthetic).
Ecological intensification is based on managing service-providing organisms that make a quantifiable direct or indirect contribution to agricultural production [Bommarco 2013: 230].
The authors specify that: “crop yield has been defined as a provisioning ecosystem service, but the yield that is harvested in a given location depends largely on several supporting and regulating services” [Bommarco 2013: 231], such as soil production and pollination. And they note that these supporting and regulating ecosystem services underpin all agricultural production, including high-input industrial systems. For example, no matter how healthy and productive a crop is, yield will suffer if it’s not well pollinated, an observation consistent with Liebig’s Law of the Minimum. “One or several of these services can limit production and, even if all other services are optimized, no or little additional output will be attained until this ecosystem service shortfall is addressed” [Bommarco 2013: 231].
Beyond fulfilling a simple mechanistic role as a medium for crops to root into, soils provide multiple ecosystem services that support crop growth.
Soil services that promote plant growth include pest and disease regulation, nutrient flow, and soil formation and structure that allow for root penetration, gas exchange, water retention, and erosion control. These processes are mediated by an immense, diverse, and largely unexplored biological community of mainly bacteria and fungi, but also protozoa, nematodes, arthropods, and earthworms [Bommarco 2013: 232].
Soil services that promote plant growth include pest and disease regulation, nutrient flow, and soil formation and structure that allow for root penetration, gas exchange, water retention, and erosion control. These processes are mediated by an immense, diverse, and largely unexplored biological community of mainly bacteria and fungi, but also protozoa, nematodes, arthropods, and earthworms [Bommarco 2013: 232].
The management practices required to activate and optimize these soil services involve increasing soil organic matter (SOM) and diversifying crop rotation.
Ecosystems also provide the regulating services of biological pest control and crop pollination. Natural pest control can enhance or maintain yield even in pesticide-based production systems. However, the overuse of pesticides can severely damage ecosystem-based pest regulation, leading to pest resurgence or crop production system collapse. Strategies to enhance pest-predator populations “include landscape-level diversification by creation or conservation of natural and resource-rich habitat, combined with directed or diversified crop rotation and decreased pesticide pressure” [Bommarco 2013: 234]. Similarly, “pollinators can be promoted at the field or farm scale by enhancing floral resources and nesting sites, thereby potentially reducing the part of the yield gap caused by pollination deficits” [Bommarco 2013: 234].
In conclusion, the authors recommend that ecological intensification strategies increasingly replace conventional, industrial practices in developed countries, where the average yield potential has largely already been met, while using ecological intensification in combination with conventional strategies to close the yield gap in parts of the world where yields are low.
World agriculture cumulatively produces enough to feed the whole human population and more, yet hundreds of millions of people on the planet are hungry due to problems of access to food. Noting that agricultural productivity is unevenly distributed around the globe, this book chapter proposes food security through ecological intensification in areas with low productivity and higher rates of hunger. This strategy runs counter to a dominant narrative that agricultural productivity even in high-input, high-yielding farming systems in industrialized countries should increase to fight world hunger. Rather, these authors posit, developed countries should adopt ecological intensification to maintain existing high levels of productivity by replacing synthetic and high-tech inputs with practices enlisting ecosystem services.
In the most productive and industrialised areas of the world the concept of ‘more with less’ is certainly engaging but rather utopic, as these agricultural systems operate mostly beyond their physical and economic efficiencies already. It is hard to get ‘more’ from these systems and this should not be a priority from a global food security perspective, as such production does not contribute to alleviate hunger in the poorest regions of the world. The greatest contribution to humanity from the most productive and industrialised areas of the world would be to maintain current productivity using less inputs of non-renewable resources and reducing their huge environmental impact; in other words, producing “the same with less” [Tittonell 2016: 23].
To illustrate the point that best practices are context specific, the authors describe a variety of approaches to ecological intensification undertaken in various parts of the world. In Uruguay, ranchers help preserve ecologically important, biodiverse grasslands by changing their grazing practices to enhance pasture and livestock productivity with no external inputs. In addition to revitalizing the grasslands, ranchers increased their incomes, allowing them to stay in business and preserve the grassland rather than selling it for conversion to crop production.
In Ethiopia, wheat productivity improves when grown under the canopy of Faidherbia albida, a prevalent local tree, which provides shade at critical moments of wheat development, increases moisture availability, and decreases the incidence of disease. “These benefits were found to result in wheat producing 23% more grain and 24% more straw under the canopy of F. albida compared to sole wheat [Tittonell 2016: 12].”
The analysis of agricultural production systems that reproduce the ecological structure of the native savannah in the Ethiopian highlands showed that biodiversity should not only be seen as a ‘service’ from farming landscapes but rather as the basis for their functioning [Tittonell 2016: 22].
The authors call for the anchoring of ecological intensification of agriculture into social, cultural and policy structures. This could be done through local innovation, policy supporting such innovation, and through multi-stakeholder platforms for dialogue bringing together researchers, local, niche innovators, and actors representing the dominant food system.
Options for the ecological intensification of agriculture can be inspired by the type of interactions between structures and functions that can be observed in nature, by the practical experience of local indigenous knowledge, and by combining these with the latest scientific knowledge and technologies. Ecological intensification calls for a constant dialogue between the practical wisdom of farmers and our own scientific wisdom [Tittonell 2016: 25].
To accelerate change, grassroots movements should seek to influence policy toward acknowledging “diversity in development directions for the agricultural sector” [Tittonell 2016: 20].
Thus, as the private sector will continue to invest in patentable technologies – understandably – to reinforce their position in the current socio-technical regime, the key role of the public sector should be to reinforce the diversity of approaches, prioritizing alternative rather than mainstream technologies, creating favorable ‘openings’ in established socio-technical regimes, and embracing the complexity and the associated transaction costs of system innovation programs or what could be called ‘co-innovation systems’. In other words, investing in the creation and support of new niches rather than supporting technological ‘solutions’ that are already embedded in current regimes [Tittonell 2016: 25].
Ecological intensification: local innovation to address global challenges, Tittonell et al. 2016
World agriculture cumulatively produces enough to feed the whole human population and more, yet hundreds of millions of people on the planet are hungry due to problems of access to food. Noting that agricultural productivity is unevenly distributed around the globe, this book chapter proposes food security through ecological intensification in areas with low productivity and higher rates of hunger. This strategy runs counter to a dominant narrative that agricultural productivity even in high-input, high-yielding farming systems in industrialized countries should increase to fight world hunger. Rather, these authors posit, developed countries should adopt ecological intensification to maintain existing high levels of productivity by replacing synthetic and high-tech inputs with practices enlisting ecosystem services.
In the most productive and industrialised areas of the world the concept of ‘more with less’ is certainly engaging but rather utopic, as these agricultural systems operate mostly beyond their physical and economic efficiencies already. It is hard to get ‘more’ from these systems and this should not be a priority from a global food security perspective, as such production does not contribute to alleviate hunger in the poorest regions of the world. The greatest contribution to humanity from the most productive and industrialised areas of the world would be to maintain current productivity using less inputs of non-renewable resources and reducing their huge environmental impact; in other words, producing “the same with less” [Tittonell 2016: 23].
To illustrate the point that best practices are context specific, the authors describe a variety of approaches to ecological intensification undertaken in various parts of the world. In Uruguay, ranchers help preserve ecologically important, biodiverse grasslands by changing their grazing practices to enhance pasture and livestock productivity with no external inputs. In addition to revitalizing the grasslands, ranchers increased their incomes, allowing them to stay in business and preserve the grassland rather than selling it for conversion to crop production.
In Ethiopia, wheat productivity improves when grown under the canopy of Faidherbia albida, a prevalent local tree, which provides shade at critical moments of wheat development, increases moisture availability, and decreases the incidence of disease. “These benefits were found to result in wheat producing 23% more grain and 24% more straw under the canopy of F. albida compared to sole wheat [Tittonell 2016: 12].”
The analysis of agricultural production systems that reproduce the ecological structure of the native savannah in the Ethiopian highlands showed that biodiversity should not only be seen as a ‘service’ from farming landscapes but rather as the basis for their functioning [Tittonell 2016: 22].
The authors call for the anchoring of ecological intensification of agriculture into social, cultural and policy structures. This could be done through local innovation, policy supporting such innovation, and through multi-stakeholder platforms for dialogue bringing together researchers, local, niche innovators, and actors representing the dominant food system.
Options for the ecological intensification of agriculture can be inspired by the type of interactions between structures and functions that can be observed in nature, by the practical experience of local indigenous knowledge, and by combining these with the latest scientific knowledge and technologies. Ecological intensification calls for a constant dialogue between the practical wisdom of farmers and our own scientific wisdom [Tittonell 2016: 25].
To accelerate change, grassroots movements should seek to influence policy toward acknowledging “diversity in development directions for the agricultural sector” [Tittonell 2016: 20].
Thus, as the private sector will continue to invest in patentable technologies – understandably – to reinforce their position in the current socio-technical regime, the key role of the public sector should be to reinforce the diversity of approaches, prioritizing alternative rather than mainstream technologies, creating favorable ‘openings’ in established socio-technical regimes, and embracing the complexity and the associated transaction costs of system innovation programs or what could be called ‘co-innovation systems’. In other words, investing in the creation and support of new niches rather than supporting technological ‘solutions’ that are already embedded in current regimes [Tittonell 2016: 25].
Ecological intensification: harnessing ecosystem services for food security, Bommarco et al. 2013
This review examines the concept of ecological intensification as a way to increase global food production by enhancing the ecological functionality of farmland.
We present ecological intensification as an alternative approach for mainstream agriculture to meet [future climatic, economic and social] challenges. Ecological intensification aims to match or augment yield levels while minimizing negative impacts on the environment and ensuing negative feedbacks on agricultural productivity, by integrating the management of ecosystem services delivered by biodiversity into crop production systems [Bommarco 2013: 230].
The idea of ecological intensification stems from the concept of ecosystem services, which refers to the benefits humans derive from ecosystems. These services are grouped into four types: supporting (such as soil formation by microorganisms), regulating (such as pest control, crop pollination, climate regulation and water purification), provisioning (such as food, fiber, fuel and water) and cultural (such as education, recreation and aesthetic).
Ecological intensification is based on managing service-providing organisms that make a quantifiable direct or indirect contribution to agricultural production [Bommarco 2013: 230].
The authors specify that: “crop yield has been defined as a provisioning ecosystem service, but the yield that is harvested in a given location depends largely on several supporting and regulating services” [Bommarco 2013: 231], such as soil production and pollination. And they note that these supporting and regulating ecosystem services underpin all agricultural production, including high-input industrial systems. For example, no matter how healthy and productive a crop is, yield will suffer if it’s not well pollinated, an observation consistent with Liebig’s Law of the Minimum. “One or several of these services can limit production and, even if all other services are optimized, no or little additional output will be attained until this ecosystem service shortfall is addressed” [Bommarco 2013: 231].
Beyond fulfilling a simple mechanistic role as a medium for crops to root into, soils provide multiple ecosystem services that support crop growth.
Soil services that promote plant growth include pest and disease regulation, nutrient flow, and soil formation and structure that allow for root penetration, gas exchange, water retention, and erosion control. These processes are mediated by an immense, diverse, and largely unexplored biological community of mainly bacteria and fungi, but also protozoa, nematodes, arthropods, and earthworms [Bommarco 2013: 232].
Soil services that promote plant growth include pest and disease regulation, nutrient flow, and soil formation and structure that allow for root penetration, gas exchange, water retention, and erosion control. These processes are mediated by an immense, diverse, and largely unexplored biological community of mainly bacteria and fungi, but also protozoa, nematodes, arthropods, and earthworms [Bommarco 2013: 232].
The management practices required to activate and optimize these soil services involve increasing soil organic matter (SOM) and diversifying crop rotation.
Ecosystems also provide the regulating services of biological pest control and crop pollination. Natural pest control can enhance or maintain yield even in pesticide-based production systems. However, the overuse of pesticides can severely damage ecosystem-based pest regulation, leading to pest resurgence or crop production system collapse. Strategies to enhance pest-predator populations “include landscape-level diversification by creation or conservation of natural and resource-rich habitat, combined with directed or diversified crop rotation and decreased pesticide pressure” [Bommarco 2013: 234]. Similarly, “pollinators can be promoted at the field or farm scale by enhancing floral resources and nesting sites, thereby potentially reducing the part of the yield gap caused by pollination deficits” [Bommarco 2013: 234].
In conclusion, the authors recommend that ecological intensification strategies increasingly replace conventional, industrial practices in developed countries, where the average yield potential has largely already been met, while using ecological intensification in combination with conventional strategies to close the yield gap in parts of the world where yields are low.
Evidence that organic farming promotes pest control, Muneret et al. 2018
Citing the problems posed globally by pesticide use and farmland expansion, this study looks at the potential of organic farming, seen as a popular prototype of ecological intensification, to limit pest infestations. Ecological intensification “is based on optimizing the ecological functions that support ecosystem services to increase the productivity of agro-ecosystems” [Muneret 2018: 361], and thus serves as a framework for evaluating farming system changes that could handle both ecological stress/collapse and human population growth. “Organic farming is a certified production system based on the principle of using farming practices that are expected to enhance ecological processes while prohibiting the use of external synthetic inputs” [Muneret 2018: 361].
Our findings in particular show that organic farming practices are able to match or outperform conventional pest control practices against pathogens and animal pests [such as insects] whereas weeds are much more abundant in organic than in conventional systems. Thus, ecological intensification based on organic farming can contribute to the control of animal pests and pathogens by enhancing biological control services and limiting their infestation levels [Muneret 2018: 365].
Whereas conventional pest control emphasizes top-down control with pesticide, ecological pest control is achieved through multiple processes:
Once established, pest populations within agro-ecosystems are affected, to varying degrees, by three ecological processes: bottom-up effects mediated by soil or plant communities involving, for instance, plant quality or habitat structure, horizontal processes within a given trophic level such as competition for resources between individuals or populations, and top-down control by natural antagonists such as predation or parasitism [Muneret 2018: 363].
Given the benefits of biodiversity for enhancing these three ecological effects, the authors explain that weeds, which are not as well suppressed in organic systems, may actually be beneficial in terms of limiting infestation by animals/insects and pathogens.
Our analysis shows that organic farming results in much higher weed infestation. This result is supported by previous studies that have shown higher abundance and diversity of plant communities within organic arable fields. We assume that this higher weed infestation, in turn, most likely influences animal pest and pathogen populations. These bottom-up effects of plant communities on higher trophic levels have been demonstrated and more abundant or diverse plant communities have been found to limit insect and disease infestation through direct and indirect mechanisms because of higher structural complexity or lower habitat quality under increased plant diversity. Although this needs further investigation, the observed performance of organic farming on animal pest and pathogen infestation may result from bottom-up effects generated by the higher weed infestation levels in organic cropping systems [Muneret 2018: 364].
Although the authors didn’t examine the effects of pest infestations on yield, they note that previous studies have suggested that weeds do not necessarily result in crop yield reductions in organic systems.
Although this needs further investigation, the observed performance of organic farming on animal pest and pathogen infestation may result from bottom-up effects generated by the higher weed infestation levels in organic cropping systems [Muneret 2018: 364].
The interplay of landscape composition and configuration: new pathways to manage functional biodiversity and agroecosystem services across Europe, Martin et al. 2019
This paper analyzes 49 studies (1515 landscapes encompassing both organic and conventional agricultural production) in Europe to determine “effects of landscape composition (% habitats) and configuration (edge density) on arthropods[7] in fields and their margins, pest control, pollination and yield” [Martin 2019: 1].
Edge density is measured as the length of edge per area of land. Edges between adjacent crop fields and between crop fields and semi-natural areas such as grasslands or other land patches not used for crops allow for “exchange between landscape patches” [Martin 2019: 4] for pollinators, pest predators and other providers of ecosystem services. High edge density is associated with smaller field size, and lower edge density with larger field size.
Complex landscapes where small and/or irregularly shaped fields and habitat patches prevail have a high density of edges. Due to increased opportunities for exchange, these landscapes are likely to support spillover of dispersal-limited populations between patches [Martin 2019: 3].
Researchers found that:
In landscapes with high edge density, 70% of pollinator and 44% of natural enemy species reached highest abundances and pollination and pest control improved 1.7- and 1.4-fold, respectively. Arable-dominated landscapes with high edge densities achieved high yields. This suggests that enhancing edge density in European agroecosystems can promote functional biodiversity and yield-enhancing ecosystem services [Martin 2019: 1].
Just as high edge density is shown here to maintain yield, low edge density, especially when combined with a lower amount of surrounding semi-natural habitat, can reduce yield.
Reduced pollination and pest control at low edge density may have been compensated by external inputs in productive landscapes. … Intermediate to low yields in landscapes with high % arable, low % semi-natural habitat and low edge density may underpin the risks of ongoing conventional intensification resulting in yield stagnation or reduction despite high agricultural inputs [Martin 2019: 9].
This article illustrates the important role of ecosystem services in maintaining crop yield, as well as the relatively simple management decisions farmers can make to enhance the habitat of arthropods providing those services.
The concept of ecological intensification in agriculture offers a framework for handling the question of how to produce enough food for a growing global human population while simultaneously protecting biodiversity. It draws on the language of ecosystem services, which includes supporting services such as soil formation, regulating services (pollination and pest control), provisioning services (production of a consumable good) and cultural services (educational and recreational) tendered by nature upon which humans depend.
Despite being anthropocentric simplification the complex web of relationships that make up an ecosystem, the concept of ecosystem services is useful in drawing attention to humanity’s reliance on nature. In the context of agriculture, recognizing the processes (especially soil creation, habitat structure, nutrient mobilization through microbe-plant symbioses, and pollination) that undergird crop health, growth and yield can enable farmers to design farming systems that meet agronomic objectives while restoring ecosystem function to cropland.
Farmers can activate certain elements of ecosystem function according to the specific problems and opportunities they see on their farmland. For example, Martin et al. [2019] explain that simply maximizing the amount of edge around cropland (for example, with smaller fields) boosts pollinator and pest predator activity in crop fields, thereby increasing yield. Similarly, increasing soil organic matter reduces the need for fertilizer, thereby reducing pressure from pests such as aphids that proliferate on nitrogen fertilized crops, and thus also reducing the need for pesticides [Garratt 2018]. Furthermore, weed colonization of cropland can potentially limit insect and pathogenic infestations [Muneret 2018].
Scenarios for meeting global food demand through ecological intensification of agriculture focus on closing yield gaps through a combination of methods in countries with low rates of agricultural productivity and higher rates of food insecurity, while maintaining yields in already high-yielding countries through a transition to ecological practices. This way, more food is produced in the places where it is most needed and in a way that minimizes (and potentially even halts) biodiversity loss. In the meantime, agricultural productivity is maintained through a transition to ecological practices in already high-yielding contexts.
The greatest contribution to humanity from the most productive and industrialized areas of the world would be to maintain current productivity using less inputs of non-renewable resources and reducing their huge environmental impact; in other words, producing ‘the same with less’ [Tittonell 2016: 23].
The greatest contribution to humanity from the most productive and industrialized areas of the world would be to maintain current productivity using less inputs of non-renewable resources and reducing their huge environmental impact; in other words, producing ‘the same with less’ [Tittonell 2016: 23].
To clarify the premises of ecological intensification, one can examine a contrasting approach to increasing global crop yield while minimizing biodiversity loss (its name, “sustainable intensification,” is similar to “ecological intensification,” and may confuse matters somewhat). An underlying premise of sustainable intensification is that increasing both agricultural yield and wildlife habitat in a particular place and time are mutually exclusive objectives.
Eschewing the viability of agricultural extensification (expanding the amount of land dedicated to agriculture), given well-known environmental problems associated with conversion of wild or semi-wild land to cropland, Egli et al. [2019] argue for increasing yields on existing farmland. The authors presume an inherent conflict between biodiversity preservation and agricultural intensification, however, where the latter is deemed achievable only through the high input methods that dramatically boosted yields in the 20th Century while also eroding biodiversity.
High input agriculture negatively affects multiple taxa and multiple dimensions of biodiversity, in particular farmland species. These negative effects have mostly been attributed to habitat simplification, inputs of fertilizer, pesticides, and irrigation.
Despite such externalities, the authors remain optimistic about future yield increases through industrial practices.
Past trends and future projections suggest large production increases through intensification on existing croplands. Yield increases contributed three quarters of the agricultural production gains between 1985 and 2005, and were mainly achieved through enhanced fertilization, irrigation and pest control, shortening of crop rotations and fallow periods, mechanization, and planting of improved crop varieties [Egli 2019: 2].
The authors introduce the possibility, therefore, of global land-use optimization: countries whose biodiversity-loss potential from agricultural intensification is lower (such as in Eastern Europe, Russia and North America) should maximize agricultural production. This would increase the global food supply enough that agricultural production could be reduced in global hotspots of biodiversity, thereby protecting biodiversity where it is the richest.
There are several problems with this strategy. As the authors themselves note, it would result in reduced agricultural output in high-biodiversity countries with an already lower level of food security and higher economic dependencies on the agricultural sector. In addition, further concentration of food production would deepen reliance on a global food system subject to international market volatility and relying on emissions-heavy long-distance shipping. Moreover, this study assumes yield on existing cropland can still increase to reach 80% of its potential, despite that yield growth from industrial innovation has already stagnated and the mineral resources that drive these techniques are dwindling. Furthermore, flooding, heat waves and droughts occurring with ever greater frequency will particularly stress crops growing in conditions of monoculture and damaged soils.
Lastly, the authors neglect to account for the potential of urban agriculture and forest food farming to increase agricultural production without converting wildlands, and, more generally, the possibility that agroecological practices are capable of increasing or maintaining yield in the process of restoring ecosystem function.
When all you have is a hammer, everything looks like a nail. For more than 50 years, the Green Revolution approach to farming has been our hammer, making weeds and insects look uniformly like nails, to stretch a metaphor. We’ve been taught to treat every other living thing growing among the crops in our fields (or yards or gardens) as the enemy, while ecological intensification teaches us to understand the interdependent relationships between different species growing together. It calls on us to search out the ways our crops benefit from the presence of various species in their midst (including the microorganisms we cannot even see), and to optimize those synergies in the skilled expression of our craft as farmers (or gardeners).
Compilation of article summaries on ecological intensification
Ecological intensification: local innovation to address global challenges, Tittonell et al. 2016
World agriculture cumulatively produces enough to feed the whole human population and more, yet hundreds of millions of people on the planet are hungry due to problems of access to food. Noting that agricultural productivity is unevenly distributed around the globe, this book chapter proposes food security through ecological intensification in areas with low productivity and higher rates of hunger. This strategy runs counter to a dominant narrative that agricultural productivity even in high-input, high-yielding farming systems in industrialized countries should increase to fight world hunger. Rather, these authors posit, developed countries should adopt ecological intensification to maintain existing high levels of productivity by replacing synthetic and high-tech inputs with practices enlisting ecosystem services.
In the most productive and industrialised areas of the world the concept of ‘more with less’ is certainly engaging but rather utopic, as these agricultural systems operate mostly beyond their physical and economic efficiencies already. It is hard to get ‘more’ from these systems and this should not be a priority from a global food security perspective, as such production does not contribute to alleviate hunger in the poorest regions of the world. The greatest contribution to humanity from the most productive and industrialised areas of the world would be to maintain current productivity using less inputs of non-renewable resources and reducing their huge environmental impact; in other words, producing “the same with less” [Tittonell 2016: 23].
To illustrate the point that best practices are context specific, the authors describe a variety of approaches to ecological intensification undertaken in various parts of the world. In Uruguay, ranchers help preserve ecologically important, biodiverse grasslands by changing their grazing practices to enhance pasture and livestock productivity with no external inputs. In addition to revitalizing the grasslands, ranchers increased their incomes, allowing them to stay in business and preserve the grassland rather than selling it for conversion to crop production.
In Ethiopia, wheat productivity improves when grown under the canopy of Faidherbia albida, a prevalent local tree, which provides shade at critical moments of wheat development, increases moisture availability, and decreases the incidence of disease. “These benefits were found to result in wheat producing 23% more grain and 24% more straw under the canopy of F. albida compared to sole wheat [Tittonell 2016: 12].”
The analysis of agricultural production systems that reproduce the ecological structure of the native savannah in the Ethiopian highlands showed that biodiversity should not only be seen as a ‘service’ from farming landscapes but rather as the basis for their functioning [Tittonell 2016: 22].
The authors call for the anchoring of ecological intensification of agriculture into social, cultural and policy structures. This could be done through local innovation, policy supporting such innovation, and through multi-stakeholder platforms for dialogue bringing together researchers, local, niche innovators, and actors representing the dominant food system.
Options for the ecological intensification of agriculture can be inspired by the type of interactions between structures and functions that can be observed in nature, by the practical experience of local indigenous knowledge, and by combining these with the latest scientific knowledge and technologies. Ecological intensification calls for a constant dialogue between the practical wisdom of farmers and our own scientific wisdom [Tittonell 2016: 25].
To accelerate change, grassroots movements should seek to influence policy toward acknowledging “diversity in development directions for the agricultural sector” [Tittonell 2016: 20].
Thus, as the private sector will continue to invest in patentable technologies – understandably – to reinforce their position in the current socio-technical regime, the key role of the public sector should be to reinforce the diversity of approaches, prioritizing alternative rather than mainstream technologies, creating favorable ‘openings’ in established socio-technical regimes, and embracing the complexity and the associated transaction costs of system innovation programs or what could be called ‘co-innovation systems’. In other words, investing in the creation and support of new niches rather than supporting technological ‘solutions’ that are already embedded in current regimes [Tittonell 2016: 25].
Ecological intensification: harnessing ecosystem services for food security, Bommarco et al. 2013
This review examines the concept of ecological intensification as a way to increase global food production by enhancing the ecological functionality of farmland.
We present ecological intensification as an alternative approach for mainstream agriculture to meet [future climatic, economic and social] challenges. Ecological intensification aims to match or augment yield levels while minimizing negative impacts on the environment and ensuing negative feedbacks on agricultural productivity, by integrating the management of ecosystem services delivered by biodiversity into crop production systems [Bommarco 2013: 230].
The idea of ecological intensification stems from the concept of ecosystem services, which refers to the benefits humans derive from ecosystems. These services are grouped into four types: supporting (such as soil formation by microorganisms), regulating (such as pest control, crop pollination, climate regulation and water purification), provisioning (such as food, fiber, fuel and water) and cultural (such as education, recreation and aesthetic).
Ecological intensification is based on managing service-providing organisms that make a quantifiable direct or indirect contribution to agricultural production [Bommarco 2013: 230].
The authors specify that: “crop yield has been defined as a provisioning ecosystem service, but the yield that is harvested in a given location depends largely on several supporting and regulating services” [Bommarco 2013: 231], such as soil production and pollination. And they note that these supporting and regulating ecosystem services underpin all agricultural production, including high-input industrial systems. For example, no matter how healthy and productive a crop is, yield will suffer if it’s not well pollinated, an observation consistent with Liebig’s Law of the Minimum. “One or several of these services can limit production and, even if all other services are optimized, no or little additional output will be attained until this ecosystem service shortfall is addressed” [Bommarco 2013: 231].
Beyond fulfilling a simple mechanistic role as a medium for crops to root into, soils provide multiple ecosystem services that support crop growth.
Soil services that promote plant growth include pest and disease regulation, nutrient flow, and soil formation and structure that allow for root penetration, gas exchange, water retention, and erosion control. These processes are mediated by an immense, diverse, and largely unexplored biological community of mainly bacteria and fungi, but also protozoa, nematodes, arthropods, and earthworms [Bommarco 2013: 232].
Soil services that promote plant growth include pest and disease regulation, nutrient flow, and soil formation and structure that allow for root penetration, gas exchange, water retention, and erosion control. These processes are mediated by an immense, diverse, and largely unexplored biological community of mainly bacteria and fungi, but also protozoa, nematodes, arthropods, and earthworms [Bommarco 2013: 232].
The management practices required to activate and optimize these soil services involve increasing soil organic matter (SOM) and diversifying crop rotation.
Ecosystems also provide the regulating services of biological pest control and crop pollination. Natural pest control can enhance or maintain yield even in pesticide-based production systems. However, the overuse of pesticides can severely damage ecosystem-based pest regulation, leading to pest resurgence or crop production system collapse. Strategies to enhance pest-predator populations “include landscape-level diversification by creation or conservation of natural and resource-rich habitat, combined with directed or diversified crop rotation and decreased pesticide pressure” [Bommarco 2013: 234]. Similarly, “pollinators can be promoted at the field or farm scale by enhancing floral resources and nesting sites, thereby potentially reducing the part of the yield gap caused by pollination deficits” [Bommarco 2013: 234].
In conclusion, the authors recommend that ecological intensification strategies increasingly replace conventional, industrial practices in developed countries, where the average yield potential has largely already been met, while using ecological intensification in combination with conventional strategies to close the yield gap in parts of the world where yields are low.
Evidence that organic farming promotes pest control, Muneret et al. 2018
Citing the problems posed globally by pesticide use and farmland expansion, this study looks at the potential of organic farming, seen as a popular prototype of ecological intensification, to limit pest infestations. Ecological intensification “is based on optimizing the ecological functions that support ecosystem services to increase the productivity of agro-ecosystems” [Muneret 2018: 361], and thus serves as a framework for evaluating farming system changes that could handle both ecological stress/collapse and human population growth. “Organic farming is a certified production system based on the principle of using farming practices that are expected to enhance ecological processes while prohibiting the use of external synthetic inputs” [Muneret 2018: 361].
Our findings in particular show that organic farming practices are able to match or outperform conventional pest control practices against pathogens and animal pests [such as insects] whereas weeds are much more abundant in organic than in conventional systems. Thus, ecological intensification based on organic farming can contribute to the control of animal pests and pathogens by enhancing biological control services and limiting their infestation levels [Muneret 2018: 365].
Whereas conventional pest control emphasizes top-down control with pesticide, ecological pest control is achieved through multiple processes:
Once established, pest populations within agro-ecosystems are affected, to varying degrees, by three ecological processes: bottom-up effects mediated by soil or plant communities involving, for instance, plant quality or habitat structure, horizontal processes within a given trophic level such as competition for resources between individuals or populations, and top-down control by natural antagonists such as predation or parasitism [Muneret 2018: 363].
Given the benefits of biodiversity for enhancing these three ecological effects, the authors explain that weeds, which are not as well suppressed in organic systems, may actually be beneficial in terms of limiting infestation by animals/insects and pathogens.
Our analysis shows that organic farming results in much higher weed infestation. This result is supported by previous studies that have shown higher abundance and diversity of plant communities within organic arable fields. We assume that this higher weed infestation, in turn, most likely influences animal pest and pathogen populations. These bottom-up effects of plant communities on higher trophic levels have been demonstrated and more abundant or diverse plant communities have been found to limit insect and disease infestation through direct and indirect mechanisms because of higher structural complexity or lower habitat quality under increased plant diversity. Although this needs further investigation, the observed performance of organic farming on animal pest and pathogen infestation may result from bottom-up effects generated by the higher weed infestation levels in organic cropping systems [Muneret 2018: 364].
Although the authors didn’t examine the effects of pest infestations on yield, they note that previous studies have suggested that weeds do not necessarily result in crop yield reductions in organic systems.
Although this needs further investigation, the observed performance of organic farming on animal pest and pathogen infestation may result from bottom-up effects generated by the higher weed infestation levels in organic cropping systems [Muneret 2018: 364].
The interplay of landscape composition and configuration: new pathways to manage functional biodiversity and agroecosystem services across Europe, Martin et al. 2019
This paper analyzes 49 studies (1515 landscapes encompassing both organic and conventional agricultural production) in Europe to determine “effects of landscape composition (% habitats) and configuration (edge density) on arthropods[7] in fields and their margins, pest control, pollination and yield” [Martin 2019: 1].
Edge density is measured as the length of edge per area of land. Edges between adjacent crop fields and between crop fields and semi-natural areas such as grasslands or other land patches not used for crops allow for “exchange between landscape patches” [Martin 2019: 4] for pollinators, pest predators and other providers of ecosystem services. High edge density is associated with smaller field size, and lower edge density with larger field size.
Complex landscapes where small and/or irregularly shaped fields and habitat patches prevail have a high density of edges. Due to increased opportunities for exchange, these landscapes are likely to support spillover of dispersal-limited populations between patches [Martin 2019: 3].
Researchers found that:
In landscapes with high edge density, 70% of pollinator and 44% of natural enemy species reached highest abundances and pollination and pest control improved 1.7- and 1.4-fold, respectively. Arable-dominated landscapes with high edge densities achieved high yields. This suggests that enhancing edge density in European agroecosystems can promote functional biodiversity and yield-enhancing ecosystem services [Martin 2019: 1].
Just as high edge density is shown here to maintain yield, low edge density, especially when combined with a lower amount of surrounding semi-natural habitat, can reduce yield.
Reduced pollination and pest control at low edge density may have been compensated by external inputs in productive landscapes. … Intermediate to low yields in landscapes with high % arable, low % semi-natural habitat and low edge density may underpin the risks of ongoing conventional intensification resulting in yield stagnation or reduction despite high agricultural inputs [Martin 2019: 9].
This article illustrates the important role of ecosystem services in maintaining crop yield, as well as the relatively simple management decisions farmers can make to enhance the habitat of arthropods providing those services.
This study combines archaeology, archaeobotany, palaeoecology and palaeoclimate investigation to shed light on the legacy of pre-Columbian land management practices on today’s Amazon rainforest. Evidence points to a millennial-scale cultivation practice that at once maintained ecosystem integrity while sustaining a large and growing human civilization.
Here, we show that persistent anthropogenic landscapes for the past 4,500 years have had an enduring legacy on the hyperdominance of edible plants in modern forests in the eastern Amazon. We found an abrupt enrichment of edible plant species in fossil lake and terrestrial records associated with pre-Columbian occupation. Our results demonstrate that, through closed-canopy forest enrichment, limited clearing for crop cultivation and low-severity fire management, long-term food security was attained despite climate and social changes. Our results suggest that, in the eastern Amazon, the subsistence basis for the development of complex societies began ~4,500 years ago with the adoption of polyculture agroforestry, combining the cultivation of multiple annual crops with the progressive enrichment of edible forest species and the exploitation of aquatic resources. This subsistence strategy intensified with the later development of Amazonian dark earths, enabling the expansion of maize cultivation to the Belterra Plateau, providing a food production system that sustained growing human populations in the eastern Amazon. Furthermore, these millennial-scale polyculture agroforestry systems have an enduring legacy on the hyperdominance of edible plants in modern forests in the eastern Amazon. Together, our data provide a long-term example of past anthropogenic land use that can inform management and conservation efforts in modern Amazonian ecosystems [Maezumi 2018: 540].
This largely hidden history of the Amazon illuminates a path forward today as humanity grapples with the combined challenges of maintaining food production for a growing global population, while preserving and restoring forests and curbing biodiversity collapse. As suggested here, ecological restoration and agricultural productivity to sustain growing populations are not mutually exclusive enterprises, but in fact can be synergistic.
Natural environments have been devastated and destroyed worldwide by recent rapid development, urbanization and industrialization. It is no exaggeration to say that the basis of human life is now threatened (Miyawaki 1982a,b).
We ecologists have been giving warnings against the devastation of nature through study results, and have produced some good effects. Besides criticism, however, we should contribute to the wholesome development of human society by active concern for nature restoration and reconstruction (Miyawaki 1975, 1981) [Miyawaki 2004: 83].
As suggested in these introductory words, Akira Miyawaki is a Japanese ecologist who has dedicated decades of his life to the study and implementation of forest restoration. He emphasizes the importance of restoring barren or degraded land more quickly than the time it takes for natural forest succession to occur, which can be 150-300 years, depending on the regional climate. By contrast, the methods he recommends can yield results within 15-20 years in terms of establishing forests mature enough to protect communities against natural disasters, such as earthquakes and storms. The principles of what has become known as the “Miyawaki Method” are based on mimicking natural forest growth patterns and thus feature: high biodiversity, preference for native species, relatively high planting densities, and healthy soil.
Communities undertaking such restoration efforts must first survey the landscape to determine the “potential vegetation” for the area based on what remains of native tree communities. Next, seeds must be gathered for some 30-50 species of native trees, and then propagated in greenhouses. After a year or two, once the seedlings have strong, well-developed roots, they can be planted. Miyawaki refers to planting events as “festivals” because the community dynamic is important for increasing public understanding of the relevance of ecological restoration and igniting a collective willingness to protect the plantings well into the future.
Miyawaki concludes with these words:
These forests of complex multilayer communities have disaster-mitigation and environmental protection functions in each region. In the Great Hanshin Earthquake, which hit the Kobe district, western Japan in January 1995, there was no damage to trees in Japanese traditional temple forests, the potential natural vegetation, however, huge structures made of non-living materials collapsed, including elevated railways, highways and tall buildings (Miyawaki 1998). On a global scale, natural forests help to avoid global warming by absorbing carbon dioxide. Restoration and regeneration of ecologically diverse forests is inevitable for citizens in every region to survive in the next century, and the next millennium [Miyawaki 2004: 89].
Using data from 130,210 forest plots across the US, this study examines the effects of tree diversity on pest invasions. The authors found that tree diversity increases pest diversity by increasing the variety of host species available (i.e., facilitation), while also decreasing establishment of pests by increasing the number of non-hosts for any given pest species relative to the total number of trees (i.e., dilution). In other words:
The relative proportion of component tree species (hosts vs. nonhosts) plays a key role in determining pest invasions, as indicated by our evidence that host diversity may promote pest diversity while neighboring nonhost species could enhance the associational resistance of host species to nonnative pest invasions [Guo 2019].
More specifically, the study observed a hump-shaped relationship between tree and pest diversity.
Pest diversity increases with tree diversity at low tree diversity (because of facilitation or amplification) and is reduced at higher tree diversity (as a result of dilution). Thus, tree diversity likely regulates forest pest invasion through both facilitation and dilution that operate simultaneously, but their relative strengths vary with overall diversity [Guo 2019].
Other factors that influence pest invasions in forest ecosystems include: “climate, resource availability, spatial scale, and habitat fragmentation related to human disturbances.” Furthermore, “recent analyses indicate that pest species continue to be introduced and spread around the globe. Under climate and land use changes, many tree species could expand, contract, or undergo latitudinal/elevational shifts in their geographical ranges” [Guo 2019].
These findings underscore the importance of biodiversity in maintaining healthy and stable ecosystems, while also highlighting the complexity of ecosystems (given the non-linear relationship between tree and pest diversity) and the challenges that poses for restoration.
Higher forest biodiversity (specifically plant functional diversity related to water, or hydraulic, transport) engenders greater ecosystem resilience to drought. This is because different species respond differently to water stress – some species slow down their release of water (and heat) through transpiration sooner than others do. Plants’ response to water availability in turn affects the local climate.
Water, carbon and energy exchanges from the land surface strongly influence the atmosphere and climate; these exchanges are dominated by plants in most ecosystems. Plant physiological responses to water stress influence these fluxes, and the resulting land-surface feedback effects influence local weather as well as the regional atmospheric circulation. Furthermore, changes in vegetation physiology and cover can drive shifts in sensible and latent heat fluxes that intensify droughts [Anderegg 2018: 538].
We have documented a fundamental effect of trait variation on ecosystem stability that directly influences the atmosphere and climate system. Temperate and boreal forest ecosystems with higher hydraulic diversity are more buffered to changing drought conditions [Anderegg 2018: 540].
This article describes three major types of microbial tree symbionts, why they matter, and maps their global distribution.
Microbial symbionts strongly influence the functioning of forest ecosystems. Root-associated microorganisms exploit inorganic, organic and/or atmospheric forms of nutrients that enable plant growth, determine how trees respond to increased concentrations of CO2, regulate the respiratory activity of soil microorganisms and affect plant species diversity by altering the strength of conspecific[5] negative density dependence [Steidinger 2019: 404].
Arbuscular mycorrhizal and ectomycorrhizal fungi and nitrogen-fixing bacteria are the focus of the study.
Plants that are involved in arbuscular mycorrhizal symbiosis comprise nearly 80% of all terrestrial plant species; these plants principally rely on arbuscular mycorrhizal fungi for enhancing mineral phosphorus uptake. In contrast to arbuscular mycorrhizal fungi, ectomycorrhizal fungi evolved from multiple lineages of saprotrophic[6] ancestors and, as a result, some ectomycorrhizal fungi are capable of directly mobilizing organic sources of soil nutrients (particularly nitrogen). Associations with ectomycorrhizal fungi—but not arbuscular mycorrhizal fungi—have previously been shown to enable trees to accelerate photosynthesis in response to increased concentrations of atmospheric CO2 when soil nitrogen is limiting, and to inhibit soil respiration by decomposer microorganisms. Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change.
In contrast to mycorrhizal fungi, which extract nutrients from the soil, symbiotic N-fixers (Rhizobia and Actinobacteria) convert atmospheric N2 to plant-usable forms. Symbiotic N-fixers are responsible for a large fraction of biological soil-nitrogen inputs, which can increase nitrogen availability in forests in which N-fixers are locally abundant [Steidinger 2019: 404].
Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change [Steidinger 2019: 404].
The study finds that climatic controls on litter breakdown determine fungi type in a given region, where colder climates favor ectomycorrhizal fungi, which are more efficient at extracting nutrients from organic material, and warmer climates favor arbuscular mycorrhizal fungi, which efficiently extract phosphorus from the soil. Warmer climates also favor nitrogen-fixing bacterial symbionts. Based on symbiosis distribution vis-a-vis existing spatial climate gradients, the authors predict changes in forest symbiosis distribution as the climate changes overtime.
To illustrate the sensitivity of global patterns of tree symbiosis to climate change, we use the relationships that we observed for current climates to project potential changes in the symbiotic status of forests in the future. Relative to our global predictions that use the most-recent climate data, model predictions that use the projected climates for 2070 suggest that the abundance of ectomycorrhizal trees will decline by as much as 10%… Our models predict that the largest declines in ectomycorrhizal abundance will occur
along the boreal–temperate ecotone, where small increases in climatic decomposition coefficients cause abrupt transitions to arbuscular mycorrhizal forests [Steidinger 2019: 407].
The authors explain that existing transitions between arbuscular and ectomycorrhizal forests are abrupt due to positive feedbacks maintaining these systems. For instance, the chemical composition of the leaves of trees forming ectomycorrhizal symbioses resists decomposition, meaning that their leaf litter reinforces the presence of ectomycorrhiza in cooler regions. Once a small temperature threshold is breached, however, climate controls on decomposition speed up litter breakdown and favor arbuscular mycorrhizal fungi, along with their tree hosts.
Red alder fix atmospheric nitrogen through a symbiosis with bacteria that colonize their roots. This study showed that when more nitrogen is produced than is needed by the plant, the resulting excess of nitric acid acts to dissolve bedrock minerals in the soil, making them available to plants.
The substantial increase in mineral weathering by N-fixing [nitrogen-fixing] alder helps explain how this species takes up 65% more P [phosphorus] and 200% more Ca [calcium] than non-fixing Douglas-fir. Enhanced access to P is most likely important to N fixers, and is used to increase photosynthetic tissue mass and N-fixing nodule production to support growth. … Ecosystem supplies of both P and Ca can limit nonfixer tree growth where N is abundant, including in our forests. Alder-enhanced uptake of rock-derived Ca and its subsequent redistribution via litterfall may especially benefit bigleaf maple and western red cedar, two nonfixers with consistently high Ca demands that have limited direct access to rock-derived nutrients [Perakisa & Pett-Ridge 2019: 5012].
This study suggests possibilities for increasing forests’ capacity to absorb carbon and mitigate climate change through the ability of red alder (and potentially other nitrogen-fixing trees) to make otherwise limiting nutrients, including nitrogen, phosphorus and calcium, available within the forest ecosystem.
Our finding that an N-fixing tree species can directly access rock-derived nutrients has implications for nutrient supplies that regulate tree growth and C uptake in forests. Inputs of fixed N can increase tree growth in N-limited forests, and could be further stimulated by access to rock-derived nutrients. Where N is already abundant and other nutrients are limiting, supplies of rock-derived nutrients can be even more important to forest growth and C uptake. It is presently unknown whether high rates of N fixation by trees are geographically widespread, and whether N fixers other than red alder can similarly access rock-derived nutrients [Perakisa & Pett-Ridge 2019: 5013].
This study illustrates the important role of soil fungi in tree population dynamics of temperate forests. In general, when a particular plant species dominates an area of land, it attracts species that feed on it. In an experiment conducted in this study, the roots of surviving seedlings had 60% fewer lesions when they were planted beneath a tree species different than their own, compared to when they were planted beneath a member of their own species, “potentially because of increased root damage by antagonists” [Bennett 2017: 2].
However, seedlings inoculated by ectomycorrhizal (EM) fungi, which forms a protective sheath around its host’s roots and also efficiently transferring nitrogen to its host,
had 840% higher survival and 75% lower lesion densities than those of uninoculated seedlings when planted beneath conspecifics [members of the same species], but inoculation had no effect beneath heterospecifics [members of another species]. In contrast, AM seedlings did not benefit from pre-inoculation, nor did pre-inoculation affect lesion densities, regardless of transplant location [Bennett 2017: 2].
Mycorrhizal fungi colonize plant roots, where they transfer nutrients to hosts in exchange for sugar produced through photosynthesis. Ectomycorrhizal fungi form a sheath around tree roots, while arbuscular mycorrhizal (AM) fungi colonize tree roots by penetrating root cell walls. In this study, as noted above, trees with EM fungi symbionts appear to be better protected from pathogens than are AM trees when growing amongst others of their own species. This dynamic affects the population dynamics of the forest by facilitating larger stands of EM trees, while inhibiting the clustering of AM trees.
In summary, “These results suggest that mycorrhizal type, through effects on plant-soil feedbacks, could be an important contributor to population regulation and community structure in temperate forests” [Bennett 2017: 1].
Industrialized forestry simplifies forest structure and harms biodiversity. To mitigate this harm, retention forestry has been adopted in places such as Sweden, where this study was conducted. “Retention forestry” avoids clearcutting and instead preserves some 5-30 percent of trees to benefit populations of birds, lichens, fungi and other types of organisms.
The authors focused on the effects of retention on ectomycorrhizal (ECM) fungi (also commonly abbreviated as “EM” fungi), an ecologically important group of species.
ECM fungi represent a large part of the biodiversity in boreal forests. They depend on carbohydrates from their host trees and are vital for forest production, as uptake of nutrients and water by the trees is mediated by the soil ECM symbiosis. ECM fungal mycelium forms a basis for soil food webs. The largely cryptic life of ECM fungi has hampered understanding of their biology and their importance for ecosystem processes, impeding adaptation of forestry to sustain ECM fungal diversity [Sterkenburg 2019: 2].
Aiming to quantify the decline in ECM fungi species abundance and richness in relation to the proportion of trees logged, the authors established an experiment with two levels of trees retained (30% and 60%), which was then compared to unlogged forest (100% retained) and clear-cut forest (0% retained). They found that ECM fungal diversity and relative abundance is preserved in proportion to the amount of retained trees.
“In clear-cuts, ECM fungal relative abundance had decreased by 95%, while ECM fungal species richness had declined by 75%, compared to unlogged plots” [Sterkenburg 2019: 1]. The latter result meant that the less common species of ECM fungi were lost, while the more dominant ones survived. The authors noted that even at the Swedish Forestry Council’s sustainability threshold of 5% tree retention (i.e., 95% logged), some 75% of ECM species are lost. In other words, there’s no significant difference between clearcutting and retaining 5% of the trees in terms of the effects on the number of fungal species lost. To preserve fungal diversity, many more trees must be retained when logging.
This study illustrates the unseen damage to forest ecosystems of intensive logging, as well as the potential challenges of re-growing forests following clear-cutting, given a likely dearth of ECM symbionts to aid sapling development.
This study models the total amount of land globally that is suitable for reforestation, finding that there is sufficient space to meet the IPCC’s recommendation of reforestation on 1 billion hectares to limit global warming to 1.5C by 2050. The potential forest land identified in this study excludes urban and agricultural land; rather, it “exists in areas that were previously degraded, dominated by sparse vegetation, grasslands, and degraded bare soils” [Bastin 2019]. Yet for reforestation efforts to meet this potential, time is of the essence. By 2050, climate change will have shrunk the additional amount of land capable of supporting forest ecosystems by about a quarter.
The restoration of trees remains among the most effective strategies for climate change mitigation. We mapped the global potential tree coverage to show that 4.4 billion hectares of canopy cover could exist under the current climate. Excluding existing trees and agricultural and urban areas, we found that there is room for an extra 0.9 billion hectares of canopy cover, which could store 205 gigatonnes of carbon in areas that would naturally support woodlands and forests.This highlights global tree restoration as our most effective climate change solution to date. However, climate change will alter this potential tree coverage.We estimate that if we cannot deviate from the current trajectory, the global potential canopy cover may shrink by ~223 million hectares by 2050, with the vast majority of losses occurring in the tropics. Our results highlight the opportunity of climate change mitigation through global tree restoration but also the urgent need for action [Bastin 2019: 1].
The restoration of trees remains among the most effective strategies for climate change mitigation [Bastin 2019: 1].
More than 50% of the tree restoration potential can be found in only six countries (in million hectares: Russia,+151; United States,+103; Canada, +78.4; Australia, +58; Brazil, +49.7; and China, +40.2), stressing the important responsibility of some of the world’s leading economies.
In order to keep global warming under the 1.5C threshold, the IPCC warns that not only must we cut carbon emissions nearly in half by 2030, we must also draw massive amounts of CO2 out of the atmosphere.
The Intergovernmental Panel on Climate Change (IPCC) suggests that around 730 billion tons of CO2 (730 petagrams of CO2, or 199 petagrams of carbon, Pg C) must be taken out of the atmosphere by the end of this century. That is equivalent to all the CO2 emitted by the United States, the United Kingdom, Germany and China since the Industrial Revolution [Lewis 2019: 25].
The IPCC further advises that forests and wooded savannas could store enough carbon to get us a quarter of the way there. “In the near term, this means adding up to 24 million hectares (Mha) of forest every year from now until 2030” [Lewis 2019: 26].
Through the Bonn Challenge, 43 countries have pledged to reforest nearly 300Mha out of a goal of 350Mha by 2030. “However, plantations are the most popular restoration plan: 45% of all commitments involve planting vast monocultures of trees as profitable enterprises” [Lewis 2019: 26], which stores much less carbon than do natural forests. Agroforestry accounts for 21% of pledged land, while natural forest regeneration accounts for 34% of commitments.
While timber plantations technically fit the definition of a forest (greater than 0.5 hectares in area, trees at least five meters high and more than 10% canopy cover, according to UN FAO),
the key components of climate-change mitigation and biodiversity protection are missing. Plantations are important economically, but they should not be classified as forest restoration. That definition urgently needs an overhaul to exclude monoculture plantations [Lewis 2019: 27].
Illustrating vast differences in mitigation potential, Lewis et al. state that “if the entire 350 Mha [of the Bonn Challenge goal] is given over to natural forests, they would store an additional 42 Pg C by 2100. Giving the same area exclusively to plantations would sequester just 1 Pg C or, if used only for agroforestry, 7 Pg C” [Lewis 2019: 27].
The authors make four specific recommendations to ensure more effective climate change mitigation through conservation and restoration efforts:
(1) Countries should significantly increase the proportion of natural forest restoration in their commitments. (Natural forest restoration over an area the size of South Carolina could store 1 Pg of carbon by 2100.)
(2) Natural forest restoration should be prioritized in the tropics, where trees grow fastest and don’t risk countering the albedo effect since there’s never any reflective snow there anyway.
(3) “Target degraded forests and partly wooded areas for natural regeneration; focus plantations and agroforestry systems on treeless regions and, where possible, select agroforestry over plantations.”
(4) Natural forest once restored must be protected.
Restoring natural forests is the best way to remove atmospheric carbon, Lewis et al. 2019
In order to keep global warming under the 1.5C threshold, the IPCC warns that not only must we cut carbon emissions nearly in half by 2030, we must also draw massive amounts of CO2 out of the atmosphere.
The Intergovernmental Panel on Climate Change (IPCC) suggests that around 730 billion tons of CO2 (730 petagrams of CO2, or 199 petagrams of carbon, Pg C) must be taken out of the atmosphere by the end of this century. That is equivalent to all the CO2 emitted by the United States, the United Kingdom, Germany and China since the Industrial Revolution [Lewis 2019: 25].
The IPCC further advises that forests and wooded savannas could store enough carbon to get us a quarter of the way there. “In the near term, this means adding up to 24 million hectares (Mha) of forest every year from now until 2030” [Lewis 2019: 26].
Through the Bonn Challenge, 43 countries have pledged to reforest nearly 300Mha out of a goal of 350Mha by 2030. “However, plantations are the most popular restoration plan: 45% of all commitments involve planting vast monocultures of trees as profitable enterprises” [Lewis 2019: 26], which stores much less carbon than do natural forests. Agroforestry accounts for 21% of pledged land, while natural forest regeneration accounts for 34% of commitments.
While timber plantations technically fit the definition of a forest (greater than 0.5 hectares in area, trees at least five meters high and more than 10% canopy cover, according to UN FAO),
the key components of climate-change mitigation and biodiversity protection are missing. Plantations are important economically, but they should not be classified as forest restoration. That definition urgently needs an overhaul to exclude monoculture plantations [Lewis 2019: 27].
Illustrating vast differences in mitigation potential, Lewis et al. state that “if the entire 350 Mha [of the Bonn Challenge goal] is given over to natural forests, they would store an additional 42 Pg C by 2100. Giving the same area exclusively to plantations would sequester just 1 Pg C or, if used only for agroforestry, 7 Pg C” [Lewis 2019: 27].
The authors make four specific recommendations to ensure more effective climate change mitigation through conservation and restoration efforts:
(1) Countries should significantly increase the proportion of natural forest restoration in their commitments. (Natural forest restoration over an area the size of South Carolina could store 1 Pg of carbon by 2100.)
(2) Natural forest restoration should be prioritized in the tropics, where trees grow fastest and don’t risk countering the albedo effect since there’s never any reflective snow there anyway.
(3) “Target degraded forests and partly wooded areas for natural regeneration; focus plantations and agroforestry systems on treeless regions and, where possible, select agroforestry over plantations.”
(4) Natural forest once restored must be protected.
The global tree restoration potential, Bastin et al. 2019
This study models the total amount of land globally that is suitable for reforestation, finding that there is sufficient space to meet the IPCC’s recommendation of reforestation on 1 billion hectares to limit global warming to 1.5C by 2050. The potential forest land identified in this study excludes urban and agricultural land; rather, it “exists in areas that were previously degraded, dominated by sparse vegetation, grasslands, and degraded bare soils” [Bastin 2019]. Yet for reforestation efforts to meet this potential, time is of the essence. By 2050, climate change will have shrunk the additional amount of land capable of supporting forest ecosystems by about a quarter.
The restoration of trees remains among the most effective strategies for climate change mitigation. We mapped the global potential tree coverage to show that 4.4 billion hectares of canopy cover could exist under the current climate. Excluding existing trees and agricultural and urban areas, we found that there is room for an extra 0.9 billion hectares of canopy cover, which could store 205 gigatonnes of carbon in areas that would naturally support woodlands and forests.This highlights global tree restoration as our most effective climate change solution to date. However, climate change will alter this potential tree coverage.We estimate that if we cannot deviate from the current trajectory, the global potential canopy cover may shrink by ~223 million hectares by 2050, with the vast majority of losses occurring in the tropics. Our results highlight the opportunity of climate change mitigation through global tree restoration but also the urgent need for action [Bastin 2019: 1].
The restoration of trees remains among the most effective strategies for climate change mitigation [Bastin 2019: 1].
More than 50% of the tree restoration potential can be found in only six countries (in million hectares: Russia,+151; United States,+103; Canada, +78.4; Australia, +58; Brazil, +49.7; and China, +40.2), stressing the important responsibility of some of the world’s leading economies.
The significance of retention trees for survival of ectomycorrhizal fungi in clear-cut Scots pine forests, Sterkenburg et al. 2019
Industrialized forestry simplifies forest structure and harms biodiversity. To mitigate this harm, retention forestry has been adopted in places such as Sweden, where this study was conducted. “Retention forestry” avoids clearcutting and instead preserves some 5-30 percent of trees to benefit populations of birds, lichens, fungi and other types of organisms.
The authors focused on the effects of retention on ectomycorrhizal (ECM) fungi (also commonly abbreviated as “EM” fungi), an ecologically important group of species.
ECM fungi represent a large part of the biodiversity in boreal forests. They depend on carbohydrates from their host trees and are vital for forest production, as uptake of nutrients and water by the trees is mediated by the soil ECM symbiosis. ECM fungal mycelium forms a basis for soil food webs. The largely cryptic life of ECM fungi has hampered understanding of their biology and their importance for ecosystem processes, impeding adaptation of forestry to sustain ECM fungal diversity [Sterkenburg 2019: 2].
Aiming to quantify the decline in ECM fungi species abundance and richness in relation to the proportion of trees logged, the authors established an experiment with two levels of trees retained (30% and 60%), which was then compared to unlogged forest (100% retained) and clear-cut forest (0% retained). They found that ECM fungal diversity and relative abundance is preserved in proportion to the amount of retained trees.
“In clear-cuts, ECM fungal relative abundance had decreased by 95%, while ECM fungal species richness had declined by 75%, compared to unlogged plots” [Sterkenburg 2019: 1]. The latter result meant that the less common species of ECM fungi were lost, while the more dominant ones survived. The authors noted that even at the Swedish Forestry Council’s sustainability threshold of 5% tree retention (i.e., 95% logged), some 75% of ECM species are lost. In other words, there’s no significant difference between clearcutting and retaining 5% of the trees in terms of the effects on the number of fungal species lost. To preserve fungal diversity, many more trees must be retained when logging.
This study illustrates the unseen damage to forest ecosystems of intensive logging, as well as the potential challenges of re-growing forests following clear-cutting, given a likely dearth of ECM symbionts to aid sapling development.
Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics, Bennett et al. 2017
This study illustrates the important role of soil fungi in tree population dynamics of temperate forests. In general, when a particular plant species dominates an area of land, it attracts species that feed on it. In an experiment conducted in this study, the roots of surviving seedlings had 60% fewer lesions when they were planted beneath a tree species different than their own, compared to when they were planted beneath a member of their own species, “potentially because of increased root damage by antagonists” [Bennett 2017: 2].
However, seedlings inoculated by ectomycorrhizal (EM) fungi, which forms a protective sheath around its host’s roots and also efficiently transferring nitrogen to its host,
had 840% higher survival and 75% lower lesion densities than those of uninoculated seedlings when planted beneath conspecifics [members of the same species], but inoculation had no effect beneath heterospecifics [members of another species]. In contrast, AM seedlings did not benefit from pre-inoculation, nor did pre-inoculation affect lesion densities, regardless of transplant location [Bennett 2017: 2].
Mycorrhizal fungi colonize plant roots, where they transfer nutrients to hosts in exchange for sugar produced through photosynthesis. Ectomycorrhizal fungi form a sheath around tree roots, while arbuscular mycorrhizal (AM) fungi colonize tree roots by penetrating root cell walls. In this study, as noted above, trees with EM fungi symbionts appear to be better protected from pathogens than are AM trees when growing amongst others of their own species. This dynamic affects the population dynamics of the forest by facilitating larger stands of EM trees, while inhibiting the clustering of AM trees.
In summary, “These results suggest that mycorrhizal type, through effects on plant-soil feedbacks, could be an important contributor to population regulation and community structure in temperate forests” [Bennett 2017: 1].
Nitrogen-fixing red alder trees tap rock-derived nutrients, Perakisa & Pett-Ridge 2019
Red alder fix atmospheric nitrogen through a symbiosis with bacteria that colonize their roots. This study showed that when more nitrogen is produced than is needed by the plant, the resulting excess of nitric acid acts to dissolve bedrock minerals in the soil, making them available to plants.
The substantial increase in mineral weathering by N-fixing [nitrogen-fixing] alder helps explain how this species takes up 65% more P [phosphorus] and 200% more Ca [calcium] than non-fixing Douglas-fir. Enhanced access to P is most likely important to N fixers, and is used to increase photosynthetic tissue mass and N-fixing nodule production to support growth. … Ecosystem supplies of both P and Ca can limit nonfixer tree growth where N is abundant, including in our forests. Alder-enhanced uptake of rock-derived Ca and its subsequent redistribution via litterfall may especially benefit bigleaf maple and western red cedar, two nonfixers with consistently high Ca demands that have limited direct access to rock-derived nutrients [Perakisa & Pett-Ridge 2019: 5012].
This study suggests possibilities for increasing forests’ capacity to absorb carbon and mitigate climate change through the ability of red alder (and potentially other nitrogen-fixing trees) to make otherwise limiting nutrients, including nitrogen, phosphorus and calcium, available within the forest ecosystem.
Our finding that an N-fixing tree species can directly access rock-derived nutrients has implications for nutrient supplies that regulate tree growth and C uptake in forests. Inputs of fixed N can increase tree growth in N-limited forests, and could be further stimulated by access to rock-derived nutrients. Where N is already abundant and other nutrients are limiting, supplies of rock-derived nutrients can be even more important to forest growth and C uptake. It is presently unknown whether high rates of N fixation by trees are geographically widespread, and whether N fixers other than red alder can similarly access rock-derived nutrients [Perakisa & Pett-Ridge 2019: 5013].
Climatic controls of decomposition drive the global biogeography of forest-tree symbioses, Steidinger et al. 2019
This article describes three major types of microbial tree symbionts, why they matter, and maps their global distribution.
Microbial symbionts strongly influence the functioning of forest ecosystems. Root-associated microorganisms exploit inorganic, organic and/or atmospheric forms of nutrients that enable plant growth, determine how trees respond to increased concentrations of CO2, regulate the respiratory activity of soil microorganisms and affect plant species diversity by altering the strength of conspecific[5] negative density dependence [Steidinger 2019: 404].
Arbuscular mycorrhizal and ectomycorrhizal fungi and nitrogen-fixing bacteria are the focus of the study.
Plants that are involved in arbuscular mycorrhizal symbiosis comprise nearly 80% of all terrestrial plant species; these plants principally rely on arbuscular mycorrhizal fungi for enhancing mineral phosphorus uptake. In contrast to arbuscular mycorrhizal fungi, ectomycorrhizal fungi evolved from multiple lineages of saprotrophic[6] ancestors and, as a result, some ectomycorrhizal fungi are capable of directly mobilizing organic sources of soil nutrients (particularly nitrogen). Associations with ectomycorrhizal fungi—but not arbuscular mycorrhizal fungi—have previously been shown to enable trees to accelerate photosynthesis in response to increased concentrations of atmospheric CO2 when soil nitrogen is limiting, and to inhibit soil respiration by decomposer microorganisms. Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change.
In contrast to mycorrhizal fungi, which extract nutrients from the soil, symbiotic N-fixers (Rhizobia and Actinobacteria) convert atmospheric N2 to plant-usable forms. Symbiotic N-fixers are responsible for a large fraction of biological soil-nitrogen inputs, which can increase nitrogen availability in forests in which N-fixers are locally abundant [Steidinger 2019: 404].
Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change [Steidinger 2019: 404].
The study finds that climatic controls on litter breakdown determine fungi type in a given region, where colder climates favor ectomycorrhizal fungi, which are more efficient at extracting nutrients from organic material, and warmer climates favor arbuscular mycorrhizal fungi, which efficiently extract phosphorus from the soil. Warmer climates also favor nitrogen-fixing bacterial symbionts. Based on symbiosis distribution vis-a-vis existing spatial climate gradients, the authors predict changes in forest symbiosis distribution as the climate changes overtime.
To illustrate the sensitivity of global patterns of tree symbiosis to climate change, we use the relationships that we observed for current climates to project potential changes in the symbiotic status of forests in the future. Relative to our global predictions that use the most-recent climate data, model predictions that use the projected climates for 2070 suggest that the abundance of ectomycorrhizal trees will decline by as much as 10%… Our models predict that the largest declines in ectomycorrhizal abundance will occur
along the boreal–temperate ecotone, where small increases in climatic decomposition coefficients cause abrupt transitions to arbuscular mycorrhizal forests [Steidinger 2019: 407].
The authors explain that existing transitions between arbuscular and ectomycorrhizal forests are abrupt due to positive feedbacks maintaining these systems. For instance, the chemical composition of the leaves of trees forming ectomycorrhizal symbioses resists decomposition, meaning that their leaf litter reinforces the presence of ectomycorrhiza in cooler regions. Once a small temperature threshold is breached, however, climate controls on decomposition speed up litter breakdown and favor arbuscular mycorrhizal fungi, along with their tree hosts.
Hydraulic diversity of forests regulates ecosystem resilience during drought, Anderegg et al. 2018
Higher forest biodiversity (specifically plant functional diversity related to water, or hydraulic, transport) engenders greater ecosystem resilience to drought. This is because different species respond differently to water stress – some species slow down their release of water (and heat) through transpiration sooner than others do. Plants’ response to water availability in turn affects the local climate.
Water, carbon and energy exchanges from the land surface strongly influence the atmosphere and climate; these exchanges are dominated by plants in most ecosystems. Plant physiological responses to water stress influence these fluxes, and the resulting land-surface feedback effects influence local weather as well as the regional atmospheric circulation. Furthermore, changes in vegetation physiology and cover can drive shifts in sensible and latent heat fluxes that intensify droughts [Anderegg 2018: 538].
We have documented a fundamental effect of trait variation on ecosystem stability that directly influences the atmosphere and climate system. Temperate and boreal forest ecosystems with higher hydraulic diversity are more buffered to changing drought conditions [Anderegg 2018: 540].
Tree diversity regulates forest pest invasion, Guo et al. 2019
Using data from 130,210 forest plots across the US, this study examines the effects of tree diversity on pest invasions. The authors found that tree diversity increases pest diversity by increasing the variety of host species available (i.e., facilitation), while also decreasing establishment of pests by increasing the number of non-hosts for any given pest species relative to the total number of trees (i.e., dilution). In other words:
The relative proportion of component tree species (hosts vs. nonhosts) plays a key role in determining pest invasions, as indicated by our evidence that host diversity may promote pest diversity while neighboring nonhost species could enhance the associational resistance of host species to nonnative pest invasions [Guo 2019].
More specifically, the study observed a hump-shaped relationship between tree and pest diversity.
Pest diversity increases with tree diversity at low tree diversity (because of facilitation or amplification) and is reduced at higher tree diversity (as a result of dilution). Thus, tree diversity likely regulates forest pest invasion through both facilitation and dilution that operate simultaneously, but their relative strengths vary with overall diversity [Guo 2019].
Other factors that influence pest invasions in forest ecosystems include: “climate, resource availability, spatial scale, and habitat fragmentation related to human disturbances.” Furthermore, “recent analyses indicate that pest species continue to be introduced and spread around the globe. Under climate and land use changes, many tree species could expand, contract, or undergo latitudinal/elevational shifts in their geographical ranges” [Guo 2019].
These findings underscore the importance of biodiversity in maintaining healthy and stable ecosystems, while also highlighting the complexity of ecosystems (given the non-linear relationship between tree and pest diversity) and the challenges that poses for restoration.
Restoration of living environment based on vegetation ecology: theory and practice, Miyawaki 2004
Natural environments have been devastated and destroyed worldwide by recent rapid development, urbanization and industrialization. It is no exaggeration to say that the basis of human life is now threatened (Miyawaki 1982a,b).
We ecologists have been giving warnings against the devastation of nature through study results, and have produced some good effects. Besides criticism, however, we should contribute to the wholesome development of human society by active concern for nature restoration and reconstruction (Miyawaki 1975, 1981) [Miyawaki 2004: 83].
As suggested in these introductory words, Akira Miyawaki is a Japanese ecologist who has dedicated decades of his life to the study and implementation of forest restoration. He emphasizes the importance of restoring barren or degraded land more quickly than the time it takes for natural forest succession to occur, which can be 150-300 years, depending on the regional climate. By contrast, the methods he recommends can yield results within 15-20 years in terms of establishing forests mature enough to protect communities against natural disasters, such as earthquakes and storms. The principles of what has become known as the “Miyawaki Method” are based on mimicking natural forest growth patterns and thus feature: high biodiversity, preference for native species, relatively high planting densities, and healthy soil.
Communities undertaking such restoration efforts must first survey the landscape to determine the “potential vegetation” for the area based on what remains of native tree communities. Next, seeds must be gathered for some 30-50 species of native trees, and then propagated in greenhouses. After a year or two, once the seedlings have strong, well-developed roots, they can be planted. Miyawaki refers to planting events as “festivals” because the community dynamic is important for increasing public understanding of the relevance of ecological restoration and igniting a collective willingness to protect the plantings well into the future.
Miyawaki concludes with these words:
These forests of complex multilayer communities have disaster-mitigation and environmental protection functions in each region. In the Great Hanshin Earthquake, which hit the Kobe district, western Japan in January 1995, there was no damage to trees in Japanese traditional temple forests, the potential natural vegetation, however, huge structures made of non-living materials collapsed, including elevated railways, highways and tall buildings (Miyawaki 1998). On a global scale, natural forests help to avoid global warming by absorbing carbon dioxide. Restoration and regeneration of ecologically diverse forests is inevitable for citizens in every region to survive in the next century, and the next millennium [Miyawaki 2004: 89].
The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon, Maezumi et al. 2018
This study combines archaeology, archaeobotany, palaeoecology and palaeoclimate investigation to shed light on the legacy of pre-Columbian land management practices on today’s Amazon rainforest. Evidence points to a millennial-scale cultivation practice that at once maintained ecosystem integrity while sustaining a large and growing human civilization.
Here, we show that persistent anthropogenic landscapes for the past 4,500 years have had an enduring legacy on the hyperdominance of edible plants in modern forests in the eastern Amazon. We found an abrupt enrichment of edible plant species in fossil lake and terrestrial records associated with pre-Columbian occupation. Our results demonstrate that, through closed-canopy forest enrichment, limited clearing for crop cultivation and low-severity fire management, long-term food security was attained despite climate and social changes. Our results suggest that, in the eastern Amazon, the subsistence basis for the development of complex societies began ~4,500 years ago with the adoption of polyculture agroforestry, combining the cultivation of multiple annual crops with the progressive enrichment of edible forest species and the exploitation of aquatic resources. This subsistence strategy intensified with the later development of Amazonian dark earths, enabling the expansion of maize cultivation to the Belterra Plateau, providing a food production system that sustained growing human populations in the eastern Amazon. Furthermore, these millennial-scale polyculture agroforestry systems have an enduring legacy on the hyperdominance of edible plants in modern forests in the eastern Amazon. Together, our data provide a long-term example of past anthropogenic land use that can inform management and conservation efforts in modern Amazonian ecosystems [Maezumi 2018: 540].
This largely hidden history of the Amazon illuminates a path forward today as humanity grapples with the combined challenges of maintaining food production for a growing global population, while preserving and restoring forests and curbing biodiversity collapse. As suggested here, ecological restoration and agricultural productivity to sustain growing populations are not mutually exclusive enterprises, but in fact can be synergistic.
Understanding what makes forests thrive is important in light of mounting calls for reforestation and forest conservation as antidotes both to species loss and climate breakdown. Moreover, distinguishing between natural forest regeneration and timber plantations is critical to achieving intended goals.
Intact forests, and especially tropical forests, sequester twice as much carbon as planted monocultures. These findings make forest conservation a critical approach to combat global warming. Because about two-thirds of all species on Earth are found in natural forests, maintaining intact forest is vital to prevent mass extinction [Dinerstein 2019: 1].
Dozens of country signatories to the Bonn Challenge have pledged to reforest nearly 300 Mha[3] out of a goal of 350 Mha by 2030. “However, plantations are the most popular restoration plan: 45% of all commitments involve planting vast monocultures of trees as profitable enterprises” [Lewis 2019: 26]. (Agroforestry accounts for 21% of pledged land, while natural forest regeneration accounts for 34% of commitments.)
Lewis et al. [2019] warn that the trend of tree plantations standing in for reforestation commitments is extremely problematic given the ecological superiority of natural, biodiverse forests. Dinerstein et al. [2019] explain why this is so.
It is no coincidence that some of the most carbon-rich ecosystems on land—natural forests—also harbor high levels of biodiversity. Evolution has generated carbon-rich forests by packing in long-lived trees that also feed stable soil carbon storage pools. This packing effect is made possible by high levels of coexistence among diverse species and growth forms, and this coexistence has been made possible by the biotic interactions that generate competition and defense. It is the very pests, pathogens, pollinators, decomposers, and predators that comprise a tropical forest that generated the carbon- rich growth forms (in both wood and soil) that take the carbon out of the atmosphere [Dinerstein 2019: 3].
It is the very pests, pathogens, pollinators, decomposers, and predators that comprise a tropical forest that generated the carbon- rich growth forms (in both wood and soil) that take the carbon out of the atmosphere [Dinerstein 2019: 3].
Shedding light on what makes forests thrive, researchers are increasingly able to explain the mechanisms by which biodiversity drives forest dynamics. For example, red alder trees, which fix nitrogen in symbiosis with soil bacteria, increase availability not only of nitrogen, but also of potassium and calcium [Perakisa & Pett-Ridge 2019]. That’s because excess fixed nitrogen acts as a weathering agent on bits of rock, leaching minerals into the soil. This increased nutrient availability removes limits to growth not only for the red alder, but also for surrounding trees.
Similarly, ectomycorrhizal (EM) fungi, in addition to providing nitrogen in exchange for photosynthate, form protective sheaths around their tree host roots, allowing saplings to develop under the canopy of parent trees [Bennett 2017]. Absent EM fungal protection, saplings are exposed to pests drawn to the immediate area due to the presence of other members of the same tree species, which draw host-specific pests. This symbiosis allows for trees forming EM symbioses to cluster, while tree species lacking this fungal partnership survive more readily as seedlings away from members of their own species. Thus, ultimately, a key driver of forest population dynamics is the unlikely, invisible, underground EM fungi.
This particular fungal type as well as countless other powerful, yet hidden, soil microorganisms are overlooked when forests are harvested for timber, and to disastrous effects. A recent Swedish study showed that: “In clear-cuts, ECM [EM] fungal relative abundance had decreased by 95%, while ECM fungal species richness had declined by 75%, compared to unlogged plots” [Sterkenburg 2019: 1]. This means that in addition to a nearly complete loss of overall soil fungal abundance, most fungal species disappeared. The researchers tested the effects of retaining 30% or 60% of trees during a logging event and found that ECM fungal diversity and relative abundance is preserved in proportion to the amount of retained trees.
One may conclude, therefore, that natural forest regeneration following timber harvest is likely to be more successful when more trees are retained, permitting the preservation of soil microorganism biodiversity. This finding of the severe effects of logging is especially important considering the key role EM fungi in particular play in mitigating and adapting to climate change.
Associations with ectomycorrhizal fungi—but not arbuscular[4] mycorrhizal fungi—have previously been shown to enable trees to accelerate photosynthesis in response to increased concentrations of atmospheric CO2 when soil nitrogen is limiting, and to inhibit soil respiration by decomposer microorganisms. Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change [Steidinger 2019: 404].
Not only microbial diversity, but alsolarge animal diversity contributes to forest productivity and carbon sequestration/storage. Mammal diversity has been found to be positively correlated to carbon concentration in the soil due to an increase in feeding interactions, where “processing of fruits and direct biomass contributions by vertebrates and plants affect soil carbon concentration” [Sobral 2017]. Similarly (though the causality here is not explained), “forested areas that contain tigers have three times the carbon density compared to forests and degraded lands where tigers have been eradicated” [Dinerstein 2019: 11]. Even humans can contribute positively to forest and other ecosystems as the Australian Martu [Penn State 2019], ancient Amazonian [Maezumi 2018] and British Columbia coastal First Nations communities [Trant 2016] have shown.
Taking the importance of biodiversity to heart, the Miyawake Method of reforestation calls for urban and rural reforestation projects where 30-50 species to be densely planted in an area “according to the system of natural forests” [Miyawaki 2004]. In combination with compost and mulch application at planting time and diligent care in the first three years after planting, this approach can within 15-20 years generate a multi-story forest capable of shielding neighboring communities from storm damage.
Compilation of article summaries on forest dynamics
Restoring natural forests is the best way to remove atmospheric carbon, Lewis et al. 2019
In order to keep global warming under the 1.5C threshold, the IPCC warns that not only must we cut carbon emissions nearly in half by 2030, we must also draw massive amounts of CO2 out of the atmosphere.
The Intergovernmental Panel on Climate Change (IPCC) suggests that around 730 billion tons of CO2 (730 petagrams of CO2, or 199 petagrams of carbon, Pg C) must be taken out of the atmosphere by the end of this century. That is equivalent to all the CO2 emitted by the United States, the United Kingdom, Germany and China since the Industrial Revolution [Lewis 2019: 25].
The IPCC further advises that forests and wooded savannas could store enough carbon to get us a quarter of the way there. “In the near term, this means adding up to 24 million hectares (Mha) of forest every year from now until 2030” [Lewis 2019: 26].
Through the Bonn Challenge, 43 countries have pledged to reforest nearly 300Mha out of a goal of 350Mha by 2030. “However, plantations are the most popular restoration plan: 45% of all commitments involve planting vast monocultures of trees as profitable enterprises” [Lewis 2019: 26], which stores much less carbon than do natural forests. Agroforestry accounts for 21% of pledged land, while natural forest regeneration accounts for 34% of commitments.
While timber plantations technically fit the definition of a forest (greater than 0.5 hectares in area, trees at least five meters high and more than 10% canopy cover, according to UN FAO),
the key components of climate-change mitigation and biodiversity protection are missing. Plantations are important economically, but they should not be classified as forest restoration. That definition urgently needs an overhaul to exclude monoculture plantations [Lewis 2019: 27].
Illustrating vast differences in mitigation potential, Lewis et al. state that “if the entire 350 Mha [of the Bonn Challenge goal] is given over to natural forests, they would store an additional 42 Pg C by 2100. Giving the same area exclusively to plantations would sequester just 1 Pg C or, if used only for agroforestry, 7 Pg C” [Lewis 2019: 27].
The authors make four specific recommendations to ensure more effective climate change mitigation through conservation and restoration efforts:
(1) Countries should significantly increase the proportion of natural forest restoration in their commitments. (Natural forest restoration over an area the size of South Carolina could store 1 Pg of carbon by 2100.)
(2) Natural forest restoration should be prioritized in the tropics, where trees grow fastest and don’t risk countering the albedo effect since there’s never any reflective snow there anyway.
(3) “Target degraded forests and partly wooded areas for natural regeneration; focus plantations and agroforestry systems on treeless regions and, where possible, select agroforestry over plantations.”
(4) Natural forest once restored must be protected.
The global tree restoration potential, Bastin et al. 2019
This study models the total amount of land globally that is suitable for reforestation, finding that there is sufficient space to meet the IPCC’s recommendation of reforestation on 1 billion hectares to limit global warming to 1.5C by 2050. The potential forest land identified in this study excludes urban and agricultural land; rather, it “exists in areas that were previously degraded, dominated by sparse vegetation, grasslands, and degraded bare soils” [Bastin 2019]. Yet for reforestation efforts to meet this potential, time is of the essence. By 2050, climate change will have shrunk the additional amount of land capable of supporting forest ecosystems by about a quarter.
The restoration of trees remains among the most effective strategies for climate change mitigation. We mapped the global potential tree coverage to show that 4.4 billion hectares of canopy cover could exist under the current climate. Excluding existing trees and agricultural and urban areas, we found that there is room for an extra 0.9 billion hectares of canopy cover, which could store 205 gigatonnes of carbon in areas that would naturally support woodlands and forests.This highlights global tree restoration as our most effective climate change solution to date. However, climate change will alter this potential tree coverage.We estimate that if we cannot deviate from the current trajectory, the global potential canopy cover may shrink by ~223 million hectares by 2050, with the vast majority of losses occurring in the tropics. Our results highlight the opportunity of climate change mitigation through global tree restoration but also the urgent need for action [Bastin 2019: 1].
The restoration of trees remains among the most effective strategies for climate change mitigation [Bastin 2019: 1].
More than 50% of the tree restoration potential can be found in only six countries (in million hectares: Russia,+151; United States,+103; Canada, +78.4; Australia, +58; Brazil, +49.7; and China, +40.2), stressing the important responsibility of some of the world’s leading economies.
The significance of retention trees for survival of ectomycorrhizal fungi in clear-cut Scots pine forests, Sterkenburg et al. 2019
Industrialized forestry simplifies forest structure and harms biodiversity. To mitigate this harm, retention forestry has been adopted in places such as Sweden, where this study was conducted. “Retention forestry” avoids clearcutting and instead preserves some 5-30 percent of trees to benefit populations of birds, lichens, fungi and other types of organisms.
The authors focused on the effects of retention on ectomycorrhizal (ECM) fungi (also commonly abbreviated as “EM” fungi), an ecologically important group of species.
ECM fungi represent a large part of the biodiversity in boreal forests. They depend on carbohydrates from their host trees and are vital for forest production, as uptake of nutrients and water by the trees is mediated by the soil ECM symbiosis. ECM fungal mycelium forms a basis for soil food webs. The largely cryptic life of ECM fungi has hampered understanding of their biology and their importance for ecosystem processes, impeding adaptation of forestry to sustain ECM fungal diversity [Sterkenburg 2019: 2].
Aiming to quantify the decline in ECM fungi species abundance and richness in relation to the proportion of trees logged, the authors established an experiment with two levels of trees retained (30% and 60%), which was then compared to unlogged forest (100% retained) and clear-cut forest (0% retained). They found that ECM fungal diversity and relative abundance is preserved in proportion to the amount of retained trees.
“In clear-cuts, ECM fungal relative abundance had decreased by 95%, while ECM fungal species richness had declined by 75%, compared to unlogged plots” [Sterkenburg 2019: 1]. The latter result meant that the less common species of ECM fungi were lost, while the more dominant ones survived. The authors noted that even at the Swedish Forestry Council’s sustainability threshold of 5% tree retention (i.e., 95% logged), some 75% of ECM species are lost. In other words, there’s no significant difference between clearcutting and retaining 5% of the trees in terms of the effects on the number of fungal species lost. To preserve fungal diversity, many more trees must be retained when logging.
This study illustrates the unseen damage to forest ecosystems of intensive logging, as well as the potential challenges of re-growing forests following clear-cutting, given a likely dearth of ECM symbionts to aid sapling development.
Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics, Bennett et al. 2017
This study illustrates the important role of soil fungi in tree population dynamics of temperate forests. In general, when a particular plant species dominates an area of land, it attracts species that feed on it. In an experiment conducted in this study, the roots of surviving seedlings had 60% fewer lesions when they were planted beneath a tree species different than their own, compared to when they were planted beneath a member of their own species, “potentially because of increased root damage by antagonists” [Bennett 2017: 2].
However, seedlings inoculated by ectomycorrhizal (EM) fungi, which forms a protective sheath around its host’s roots and also efficiently transferring nitrogen to its host,
had 840% higher survival and 75% lower lesion densities than those of uninoculated seedlings when planted beneath conspecifics [members of the same species], but inoculation had no effect beneath heterospecifics [members of another species]. In contrast, AM seedlings did not benefit from pre-inoculation, nor did pre-inoculation affect lesion densities, regardless of transplant location [Bennett 2017: 2].
Mycorrhizal fungi colonize plant roots, where they transfer nutrients to hosts in exchange for sugar produced through photosynthesis. Ectomycorrhizal fungi form a sheath around tree roots, while arbuscular mycorrhizal (AM) fungi colonize tree roots by penetrating root cell walls. In this study, as noted above, trees with EM fungi symbionts appear to be better protected from pathogens than are AM trees when growing amongst others of their own species. This dynamic affects the population dynamics of the forest by facilitating larger stands of EM trees, while inhibiting the clustering of AM trees.
In summary, “These results suggest that mycorrhizal type, through effects on plant-soil feedbacks, could be an important contributor to population regulation and community structure in temperate forests” [Bennett 2017: 1].
Nitrogen-fixing red alder trees tap rock-derived nutrients, Perakisa & Pett-Ridge 2019
Red alder fix atmospheric nitrogen through a symbiosis with bacteria that colonize their roots. This study showed that when more nitrogen is produced than is needed by the plant, the resulting excess of nitric acid acts to dissolve bedrock minerals in the soil, making them available to plants.
The substantial increase in mineral weathering by N-fixing [nitrogen-fixing] alder helps explain how this species takes up 65% more P [phosphorus] and 200% more Ca [calcium] than non-fixing Douglas-fir. Enhanced access to P is most likely important to N fixers, and is used to increase photosynthetic tissue mass and N-fixing nodule production to support growth. … Ecosystem supplies of both P and Ca can limit nonfixer tree growth where N is abundant, including in our forests. Alder-enhanced uptake of rock-derived Ca and its subsequent redistribution via litterfall may especially benefit bigleaf maple and western red cedar, two nonfixers with consistently high Ca demands that have limited direct access to rock-derived nutrients [Perakisa & Pett-Ridge 2019: 5012].
This study suggests possibilities for increasing forests’ capacity to absorb carbon and mitigate climate change through the ability of red alder (and potentially other nitrogen-fixing trees) to make otherwise limiting nutrients, including nitrogen, phosphorus and calcium, available within the forest ecosystem.
Our finding that an N-fixing tree species can directly access rock-derived nutrients has implications for nutrient supplies that regulate tree growth and C uptake in forests. Inputs of fixed N can increase tree growth in N-limited forests, and could be further stimulated by access to rock-derived nutrients. Where N is already abundant and other nutrients are limiting, supplies of rock-derived nutrients can be even more important to forest growth and C uptake. It is presently unknown whether high rates of N fixation by trees are geographically widespread, and whether N fixers other than red alder can similarly access rock-derived nutrients [Perakisa & Pett-Ridge 2019: 5013].
Climatic controls of decomposition drive the global biogeography of forest-tree symbioses, Steidinger et al. 2019
This article describes three major types of microbial tree symbionts, why they matter, and maps their global distribution.
Microbial symbionts strongly influence the functioning of forest ecosystems. Root-associated microorganisms exploit inorganic, organic and/or atmospheric forms of nutrients that enable plant growth, determine how trees respond to increased concentrations of CO2, regulate the respiratory activity of soil microorganisms and affect plant species diversity by altering the strength of conspecific[5] negative density dependence [Steidinger 2019: 404].
Arbuscular mycorrhizal and ectomycorrhizal fungi and nitrogen-fixing bacteria are the focus of the study.
Plants that are involved in arbuscular mycorrhizal symbiosis comprise nearly 80% of all terrestrial plant species; these plants principally rely on arbuscular mycorrhizal fungi for enhancing mineral phosphorus uptake. In contrast to arbuscular mycorrhizal fungi, ectomycorrhizal fungi evolved from multiple lineages of saprotrophic[6] ancestors and, as a result, some ectomycorrhizal fungi are capable of directly mobilizing organic sources of soil nutrients (particularly nitrogen). Associations with ectomycorrhizal fungi—but not arbuscular mycorrhizal fungi—have previously been shown to enable trees to accelerate photosynthesis in response to increased concentrations of atmospheric CO2 when soil nitrogen is limiting, and to inhibit soil respiration by decomposer microorganisms. Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change.
In contrast to mycorrhizal fungi, which extract nutrients from the soil, symbiotic N-fixers (Rhizobia and Actinobacteria) convert atmospheric N2 to plant-usable forms. Symbiotic N-fixers are responsible for a large fraction of biological soil-nitrogen inputs, which can increase nitrogen availability in forests in which N-fixers are locally abundant [Steidinger 2019: 404].
Because increased plant photosynthesis and decreased soil respiration both reduce atmospheric CO2 concentrations, the ectomycorrhizal symbiosis is associated with buffering the Earth’s climate against anthropogenic change [Steidinger 2019: 404].
The study finds that climatic controls on litter breakdown determine fungi type in a given region, where colder climates favor ectomycorrhizal fungi, which are more efficient at extracting nutrients from organic material, and warmer climates favor arbuscular mycorrhizal fungi, which efficiently extract phosphorus from the soil. Warmer climates also favor nitrogen-fixing bacterial symbionts. Based on symbiosis distribution vis-a-vis existing spatial climate gradients, the authors predict changes in forest symbiosis distribution as the climate changes overtime.
To illustrate the sensitivity of global patterns of tree symbiosis to climate change, we use the relationships that we observed for current climates to project potential changes in the symbiotic status of forests in the future. Relative to our global predictions that use the most-recent climate data, model predictions that use the projected climates for 2070 suggest that the abundance of ectomycorrhizal trees will decline by as much as 10%… Our models predict that the largest declines in ectomycorrhizal abundance will occur
along the boreal–temperate ecotone, where small increases in climatic decomposition coefficients cause abrupt transitions to arbuscular mycorrhizal forests [Steidinger 2019: 407].
The authors explain that existing transitions between arbuscular and ectomycorrhizal forests are abrupt due to positive feedbacks maintaining these systems. For instance, the chemical composition of the leaves of trees forming ectomycorrhizal symbioses resists decomposition, meaning that their leaf litter reinforces the presence of ectomycorrhiza in cooler regions. Once a small temperature threshold is breached, however, climate controls on decomposition speed up litter breakdown and favor arbuscular mycorrhizal fungi, along with their tree hosts.
Hydraulic diversity of forests regulates ecosystem resilience during drought, Anderegg et al. 2018
Higher forest biodiversity (specifically plant functional diversity related to water, or hydraulic, transport) engenders greater ecosystem resilience to drought. This is because different species respond differently to water stress – some species slow down their release of water (and heat) through transpiration sooner than others do. Plants’ response to water availability in turn affects the local climate.
Water, carbon and energy exchanges from the land surface strongly influence the atmosphere and climate; these exchanges are dominated by plants in most ecosystems. Plant physiological responses to water stress influence these fluxes, and the resulting land-surface feedback effects influence local weather as well as the regional atmospheric circulation. Furthermore, changes in vegetation physiology and cover can drive shifts in sensible and latent heat fluxes that intensify droughts [Anderegg 2018: 538].
We have documented a fundamental effect of trait variation on ecosystem stability that directly influences the atmosphere and climate system. Temperate and boreal forest ecosystems with higher hydraulic diversity are more buffered to changing drought conditions [Anderegg 2018: 540].
Tree diversity regulates forest pest invasion, Guo et al. 2019
Using data from 130,210 forest plots across the US, this study examines the effects of tree diversity on pest invasions. The authors found that tree diversity increases pest diversity by increasing the variety of host species available (i.e., facilitation), while also decreasing establishment of pests by increasing the number of non-hosts for any given pest species relative to the total number of trees (i.e., dilution). In other words:
The relative proportion of component tree species (hosts vs. nonhosts) plays a key role in determining pest invasions, as indicated by our evidence that host diversity may promote pest diversity while neighboring nonhost species could enhance the associational resistance of host species to nonnative pest invasions [Guo 2019].
More specifically, the study observed a hump-shaped relationship between tree and pest diversity.
Pest diversity increases with tree diversity at low tree diversity (because of facilitation or amplification) and is reduced at higher tree diversity (as a result of dilution). Thus, tree diversity likely regulates forest pest invasion through both facilitation and dilution that operate simultaneously, but their relative strengths vary with overall diversity [Guo 2019].
Other factors that influence pest invasions in forest ecosystems include: “climate, resource availability, spatial scale, and habitat fragmentation related to human disturbances.” Furthermore, “recent analyses indicate that pest species continue to be introduced and spread around the globe. Under climate and land use changes, many tree species could expand, contract, or undergo latitudinal/elevational shifts in their geographical ranges” [Guo 2019].
These findings underscore the importance of biodiversity in maintaining healthy and stable ecosystems, while also highlighting the complexity of ecosystems (given the non-linear relationship between tree and pest diversity) and the challenges that poses for restoration.
Restoration of living environment based on vegetation ecology: theory and practice, Miyawaki 2004
Natural environments have been devastated and destroyed worldwide by recent rapid development, urbanization and industrialization. It is no exaggeration to say that the basis of human life is now threatened (Miyawaki 1982a,b).
We ecologists have been giving warnings against the devastation of nature through study results, and have produced some good effects. Besides criticism, however, we should contribute to the wholesome development of human society by active concern for nature restoration and reconstruction (Miyawaki 1975, 1981) [Miyawaki 2004: 83].
As suggested in these introductory words, Akira Miyawaki is a Japanese ecologist who has dedicated decades of his life to the study and implementation of forest restoration. He emphasizes the importance of restoring barren or degraded land more quickly than the time it takes for natural forest succession to occur, which can be 150-300 years, depending on the regional climate. By contrast, the methods he recommends can yield results within 15-20 years in terms of establishing forests mature enough to protect communities against natural disasters, such as earthquakes and storms. The principles of what has become known as the “Miyawaki Method” are based on mimicking natural forest growth patterns and thus feature: high biodiversity, preference for native species, relatively high planting densities, and healthy soil.
Communities undertaking such restoration efforts must first survey the landscape to determine the “potential vegetation” for the area based on what remains of native tree communities. Next, seeds must be gathered for some 30-50 species of native trees, and then propagated in greenhouses. After a year or two, once the seedlings have strong, well-developed roots, they can be planted. Miyawaki refers to planting events as “festivals” because the community dynamic is important for increasing public understanding of the relevance of ecological restoration and igniting a collective willingness to protect the plantings well into the future.
Miyawaki concludes with these words:
These forests of complex multilayer communities have disaster-mitigation and environmental protection functions in each region. In the Great Hanshin Earthquake, which hit the Kobe district, western Japan in January 1995, there was no damage to trees in Japanese traditional temple forests, the potential natural vegetation, however, huge structures made of non-living materials collapsed, including elevated railways, highways and tall buildings (Miyawaki 1998). On a global scale, natural forests help to avoid global warming by absorbing carbon dioxide. Restoration and regeneration of ecologically diverse forests is inevitable for citizens in every region to survive in the next century, and the next millennium [Miyawaki 2004: 89].
The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon, Maezumi et al. 2018
This study combines archaeology, archaeobotany, palaeoecology and palaeoclimate investigation to shed light on the legacy of pre-Columbian land management practices on today’s Amazon rainforest. Evidence points to a millennial-scale cultivation practice that at once maintained ecosystem integrity while sustaining a large and growing human civilization.
Here, we show that persistent anthropogenic landscapes for the past 4,500 years have had an enduring legacy on the hyperdominance of edible plants in modern forests in the eastern Amazon. We found an abrupt enrichment of edible plant species in fossil lake and terrestrial records associated with pre-Columbian occupation. Our results demonstrate that, through closed-canopy forest enrichment, limited clearing for crop cultivation and low-severity fire management, long-term food security was attained despite climate and social changes. Our results suggest that, in the eastern Amazon, the subsistence basis for the development of complex societies began ~4,500 years ago with the adoption of polyculture agroforestry, combining the cultivation of multiple annual crops with the progressive enrichment of edible forest species and the exploitation of aquatic resources. This subsistence strategy intensified with the later development of Amazonian dark earths, enabling the expansion of maize cultivation to the Belterra Plateau, providing a food production system that sustained growing human populations in the eastern Amazon. Furthermore, these millennial-scale polyculture agroforestry systems have an enduring legacy on the hyperdominance of edible plants in modern forests in the eastern Amazon. Together, our data provide a long-term example of past anthropogenic land use that can inform management and conservation efforts in modern Amazonian ecosystems [Maezumi 2018: 540].
This largely hidden history of the Amazon illuminates a path forward today as humanity grapples with the combined challenges of maintaining food production for a growing global population, while preserving and restoring forests and curbing biodiversity collapse. As suggested here, ecological restoration and agricultural productivity to sustain growing populations are not mutually exclusive enterprises, but in fact can be synergistic.
As in every edition of this compendium, here we assemble and summarize research offering evidence of the power of ecosystems to address climate breakdown. The themes presented:
forest dynamics
ecological intensification and
transformative change
were chosen based on recurrent themes of mostly recent reports and studies. Not surprisingly given its centrality to ecosystem function, the idea of biodiversity weaves through all three themes.
Forests, especially tropical forests, are considered the lungs of the world because of their vigorous breathing in of carbon dioxide and breathing out of oxygen. Protecting existing forests and regenerating previously cleared forests is widely understood as a key component of mitigating climate change. What’s not as well appreciated is the role of biodiversity in the ecological functionality of forests. For reforestation to sequester requisite amounts of CO2, slow down water during storms and provide adequate habitat for species otherwise condemned to extinction, reforestation should aim to closely mimic natural, biodiverse forests. The studies presented here examine various symbioses in natural forests that drive forest dynamics. The ecosystem simplification that comes with monoculture plantations risks losing these important interspecies partnerships.
Ecological intensification relies on biodiversity to address agriculture’s double objective of increased productivity with minimal environmental harm. Upending deeply ingrained beliefs that high yield and ecosystem regeneration are mutually exclusive outcomes, ecological intensification focuses on replacing high-tech inputs with ecosystem services capable of delivering equivalent agronomic outcomes. To put ecological intensification’s productivity potential in perspective, consider that natural ecosystems are far more productive and resistant to pest infestations than are croplands, despite no fertilizer or pesticide use. As for crop yield? A core difference between ecology-based versus high-tech/high-input farming systems is the variety of crops harvested, due to more complex rotations on ecologically managed farms. However, the total per-acre yield between the two systems is likely to be comparable. Moreover, because ecological farming practices protect and build the soil, they are likely to be more productive in the future as drought, heat waves and flooding become more frequent. In short, a global proliferation of highly productive, ecologically functional farms can contribute enormously to healing the planet.
Transformative change – how does that relate to the theme of biodiversity? First, a growing awareness of biodiversity’s sharp global decline is prompting a spate of proposals for transformative change aiming to restore not only biodiversity, but also climate stability and democracy. Second, biodiversity in a figurative sense, meaning human diversity represented by hundreds of thousands of tiny NGOs in every corner of the globe, is rising from the embers of a civilization in decay to solve the problems threatening our collective wellbeing and survival. It is within the culturally, socially and geographically diverse local communities of the world that transformative change is most readily initiated and achieved. This local action has been dubbed ‘blessed unrest’ by author Paul Hawken [Hawken 2007]. It represents a spontaneous, decentralized, leaderless global movement approximating humanity’s immune system response to the ills caused by a long-standing vacuum of integrity and goodwill among the governments of the world.
In the concluding portion of this document, we recount just a handful of the millions of untold stories of blessed unrest. Perhaps these will be the stories that future generations tell to explain what catalysed the transformative systemic change that rescued humanity from the brink of social and ecological collapse. Leading up to that, we report on relevant discoveries and analyses from scientists and others who are putting the pieces together to explain not only the extent of harm done to the planet, but also to illuminate some of the drivers of the biosphere’s vitality, and how to navigate the way forward from here.
Guest author Gerald Pollack introduces a fundamental shift in how we view water. It has the potential to significantly alter our understandings of any processes that involve water, including aspects of climate, biology, and how we approach eco-restoration.
The Fourth Phase of Water: Beyond Solid, Liquid, and Vapor
Gerald H. Pollack, PhD, Professor of Bioengineering, University of Washington
How can a Jesus Christ lizard walk on water? Why do pollen grains jitterbug in a puddle? Why do fair weather clouds form such lovely puffy white shapes? Why do your joints work without squeaking? Why do sprained ankles swell within seconds?
Answering these questions requires an understanding of water. Given water’s simplicity and pervasiveness through nature, we presume that water must be completely understood, but in fact, precious little is known about how water molecules line up — until recently.
Students learn that water has three phases: solid, liquid and vapor. But there is something more: in our laboratory at the University of Washington we have uncovered a fourth phase. This phase occurs next to water loving (hydrophilic) surfaces. It is surprisingly extensive, projecting out from surfaces by up to millions of molecular layers. And it exists almost everywhere throughout nature, including your body.
This freshly identified phase of water has been described in a recent book: The Fourth Phase of Water: Beyond Solid, Liquid and Vapor. The book documents the basic findings and presents many applications including the ones mentioned above. It also deals with water’s many anomalies, turning those anomalies into easily explained features.
The existence of a fourth phase may seem unexpected. However, it should not be entirely so: a century ago, the physical chemist Sir William Hardy argued for the existence of a fourth phase; and many authors over the years have found evidence for some kind of “ordered” or “structured” phase of water. Fresh experimental evidence not only confirms the existence of such an ordered, liquid-crystalline phase, but also details its properties. Those properties explain everyday observations and answer questions ranging from why gelatin desserts hold their water, to why teapots whistle.
The presence of the fourth phase carries many implications. Here, I outline some basic features of this phase, and then deal with several of those implications. I will touch on atmospheric science, and then focus on some biological and health applications.
Does Water Transduce Energy?
The energy for building water structure comes from the sun. Radiant energy converts ordinary bulk water into ordered water, building this ordered zone. We found that all wavelengths ranging from UV through visible to infrared can build this ordered water. Near-infrared energy is the most capable. Water absorbs infrared energy freely from the environment; it uses that energy to convert bulk water into liquid crystalline water (fourth phase water) — which we also call “exclusion zone” or “EZ” water because it profoundly excludes solutes. Hence, buildup of EZ water occurs naturally and spontaneously from environmental energy. Additional energy input creates additional EZ buildup.
Figure 1. Diagrammatic representation of EZ water, negatively charged,
and the positively charged bulk water beyond. Hydrophilic surface at left.
Of particular significance is the fourth phase’s charge: commonly negative (Figure 1). Absorbed radiant energy splits water molecules; the negative moiety constitutes the building block of the EZ, while the positive moiety binds with water molecules to form free hydronium ions, which diffuse throughout the water. Adding additional light stimulates more charge separation.
This process resembles the first step of photosynthesis. In that step, energy from the sun splits water molecules. Hydrophilic chromophores catalyze the splitting. The process considered here is similar but more generic: any hydrophilic surface may catalyze the splitting. Some surfaces work more effectively than others.
The separated charges resemble a battery. That battery can deliver energy in a manner similar to the way the separated charges in plants deliver energy. Plants, of course, comprise mostly water, and it is therefore no surprise that similar energy conversion takes place in water itself.
Figure 2. Practically incessant flow occurs through hydrophilic tubes immersed in water
The stored electrical energy in water can drive various kinds of work, including flow. An example is the axial flow through tubes. We found that immersing tubes made of hydrophilic materials into water produces flow through those tubes, similar to blood flow through blood vessels (Figure 2). The driving energy comes from the radiant energy absorbed and stored in the water. Nothing more. Flow may persist undiminished for many hours, even days. Additional incident light brings faster flow. This is not a perpetual motion machine: incident radiant energy drives the flow — in much the same way that it drives vascular flow in plants. And, we have fresh evidence (see below) that it also assists the heart in driving blood in the cardiovascular system.
Applications in Biological Flow and Atmospheric Science
The water-based energy conversion framework is rich with implication for many systems involving water. These systems may range from biology and chemistry all the way to atmospheric science and engineering. The fourth phase appears nearly everywhere: all that’s needed is water, radiant energy, and a hydrophilic surface. The latter can be as large as a slab of polymer and as small as a dissolved molecule. The liquid crystalline phase inevitably builds — and its presence plays some integral role in the system’s behavior.
Let me provide a few representative examples.
One example is…you. Two thirds of your cells are water — by volume. In terms of the molecular fraction, that fraction translates to more than 99% because so many of those diminutive molecules are required to build that two-thirds volume fraction. Modern cell biology considers that 99% fraction of your molecules as mere background carriers of the “important” molecules of life such as proteins and nucleic acids. Conventional wisdom asserts that 99% of your molecules don’t do very much.
However, EZ water envelops every macromolecule in the cell. Those macromolecules are so tightly packed that the enveloping liquid crystalline water largely fills your cells. In other words, most of your cell water is liquid crystalline, or EZ water. This water plays a central role in everything the cell does — as elaborated in my earlier book, Cells, Gels and the Engines of Life.
What’s new is the role of radiant energy: incident radiant energy powers many of those cellular functions. An example is the blood flowing through your capillaries. That blood eventually encounters high resistance: capillaries are often narrower than the red blood cells that must pass through them; to make their way through, those red cells need to bend and contort. Resistance is high. You’d anticipate the need for lots of driving pressure; yet, the pressure gradient across the capillary bed is negligible. The paradox resolves if radiant energy helps propel flow through capillaries in the same way that it propels flow through hydrophilic tubes. Radiant energy may constitute an unsuspected source of vascular drive, supplementing cardiac pressure.
Why you feel good after a sauna now seems understandable. If radiant energy drives capillary flow and ample capillary flow is important for optimal functioning, then sitting in the sauna will inevitably be a feel-good experience. The infrared energy associated with heat should help drive that flow. The same if you walk out into sunlight: we presume that the feel-good experience derives purely from the psychological realm; but the evidence above implies that sunlight may build your body’s EZs. Fully built EZs around each protein seem necessary for optimal cellular functioning.
A second example of the EZ’s central role is weather. Common understanding of weather derives from two principal variables: temperature and pressure. Those two variables are said to explain virtually everything we experience in terms of weather. However, the atmosphere also contains water: it is full of micrometer-scale droplets commonly known as aerosol droplets or aerosol particles. Those droplets make up atmospheric humidity. When the atmosphere is humid, the many droplets scatter considerable light, causing haze; you can’t see clearly through that haze. When the atmosphere contains only few droplets, you may see clearly, over long distances.
The Fourth Phase book presents evidence for the structure of those droplets. It shows that EZ water envelops each droplet, while hydronium ions occupy the droplets’ interior. Repelling one another, those internal hydronium ions create pressure, which pushes against the robust shell of EZ water. That explains why droplets tend toward roundness.
How do those aerosol droplets condense to form clouds? The droplets’ EZ shells bear negative charge. Negatively charged droplets should repel one another, precluding any condensation into clouds. Those like-charged aerosol droplets should remain widely dispersed throughout the atmosphere. However, droplets do often condense into clouds, and the question is how that can happen.
The reason they condense is because of the unlike charges that lie in between the droplets. Richard Feynman, the legendary Nobel Prize physicist of the late 20th century understood the principle, opining that: “like-likes-like because of an intermediate of unlikes.” The like-charged droplets “like” one another, so they come together; the unlike charges lying in between those droplets constitute the attractors (Figure 3).
Figure 3: Like-charged entities attract because of an intermediate of
opposite charge.
The like-likes-like principle has been widely appreciated, but also widely ignored: after all, how could like charges conceivably attract? A reason why this powerfully simple concept has been ignored is that the source of the unlike charges has been difficult to identify. We now know that the unlike charges can come from the splitting of water — the negative components building EZ shells, while the corresponding positive components provide the unlike attractors. With enough of those attractors, the negatively charged aerosol droplets may condense into clouds.
These two phenomena, radiant energy-induced biological function and like-likes-like cloud formation, provide examples of how water’s energy can account for phenomena not otherwise explained. The fourth phase is the key building block that allows for construction of an edifice of understanding.
Practical Applications
Beyond scientific, the discovery of the fourth phase has practical applications. They include flow production (already mentioned), electrical energy harvesting, and even filtration. I briefly mention the latter two applications.
Filtration occurs naturally because the liquid crystalline phase massively excludes solutes and particles in much the same way as does ice. Accordingly, fourth phase water is essentially solute free. Collecting it provides solute-free and bacteria-free water. A working prototype has confirmed this expectation. Purification by this method requires no physical filter: the fourth phase itself does the separation, and the energy comes from the sun.
Energy harvesting seems straightforward: light drives the separation of charge, and those separated charges constitute a battery. Harvesting electrical energy should be realizable with proper electrodes. This technology development is underway in our spinoff company, and has the potential to replace standard photovoltaic systems with simpler ones based on water. More detail on these practical applications can be found in the Pollack laboratory homepage: <http://faculty.washington.edu/ghp/>.
Practical applications also exist within our bodies, and I present two of them: why your joints don’t squeak: and why dislocated or sprained joints will swell within seconds.
Joints are sites at which bones press upon one another (Figure 4). The bones may also rotate, as during deep-knee bends and push-ups. You’d think that rotation under pressure might elicit squeaky frictional resistance, but joint friction remains remarkably modest. Why so?
Figure 4. Enveloping the joint, the capsule ensures that the fluid’s hydronium ions don’t disperse. The concentrated hydronium ions repel, keeping surfaces apart and assuring low frictions.
The ends of bones are lined with cartilage. Those cartilaginous materials do the actual pressing. Hence, the issue of joint friction reduces to the issue of the cartilaginous surfaces and the synovial fluid lying in between. How does this system behave under pressure?
Cartilage is made of classic gel materials: highly charged polymers and water; therefore, cartilage is a gel. Gel surfaces bear EZs, so cartilage surfaces should likewise bear EZs. The splitting of water associated with EZ buildup creates many hydronium ions in the synovial fluid between. Additional hydronium ions come from the molecules within that fluid, creating their own EZs and protons. Thus, many hydronium ions will lie in the area where two cartilaginous surfaces lie across from one another. The repulsive force coming from those hydronium ions should keep the cartilage surfaces apart — some investigators maintain that despite heavy loads, the cartilage surfaces never touch. That separation means that any rough spots, or asperities, will never come into contact as the respective surfaces shear past one another; and that in turn means low friction.
For such a mechanism to work, some kind of built-in restraint should be present to keep the repelling hydronium ions in place. Otherwise, they may be forced out of the local region, compromising lubrication. Nature provides that safety net: a structure known as the joint capsule envelops the joint. By constraining the dispersal of hydronium ions, that encapsulation ensures low friction. That’s why your joints don’t ordinarily squeak.
Figure 5. Example of post-injury swelling.
Regarding swelling, the second issue under consideration, osmosis evidently plays a role. Since the cell is packed with negatively charged proteins, the cytoplasm should generate an osmotic draw similar to the osmotic draw generated by diapers or gels. Physiologists know that it does.
A peculiar feature of cells, however, is their relatively modest water content. Compared to 20:1 or higher for many common gels, the cell’s water-to-solids ratio is only about 2:1. The many negatively charged macromolecules of the cell should generate a strong osmotic draw; yet the water content in the cell remains surprisingly low. That limited water content may come as a consequence of the macromolecular network’s stiffness: cellular networks typically comprise tubular or multi-stranded biopolymers tightly cross-linked to one another. The resultant stiffness prevents the network from expanding to its full osmotic potential.
If those cross-links were to disrupt, however, then the full power of osmotic draw would take effect; the tissue could then build many EZ layers and therefore hydrate massively, bringing huge expansion (Figure 5). That’s what happens when body tissues are injured, especially with dislocations. The injury disrupts fibrous macromolecules and cross-links, eliminating the restraining forces that keep osmosis at bay; EZ buildup can then proceed virtually unimpeded.
The reason why swelling can be so impressive is that the cross-link disruption occurs progressively. Breaking one cross-link results in higher stress on neighboring cross-links; so disruption progresses in a zipper-like fashion. When that happens, the osmotic rush of water into the tissue can continue practically without restraint, resulting in the enormous immediate swelling that is often seen. The tissue will return to normal only when cross-links repair and the matrix returns to its normally restraining configuration.
Water and Healing
During childhood illness, grandmothers and doctors will often advise: “drink more water.” In his now-classical book, sub-titled Your Body’s Many Cries for Water: You Are Not Sick, You Are Thirsty, the Iranian physician Fereydoon Batmanghelidj confirms the wisdom of this quaint advice. The author documents years of clinical practice showing reversal of diverse pathologies simply by drinking more water. Hydration is critical.
Batmanghelidj’s experience meshes with evidence of healing from special waters such as those from the Ganges and Lourdes. Those waters most often come from deep underground springs or from glacial melt. Spring waters experience pressure from above; pressure converts liquid water into EZ water because of EZ water’s higher density. So, spring water’s healing quality may arise not only from its mineral content but also from its relatively high EZ content.
The same for mountain water: it too should have high EZ content. Our studies have shown that ice formation requires an EZ intermediate; i.e., bulk water does not convert directly to ice; it converts to EZ, which then converts to ice. Similarly for melting: melting ice forms EZ, which subsequently converts to bulk water. Fresh ice melt contains abundant EZ water.
For spring water and fresh ice melt, then, the high EZ content may explain the recognized health benefits. EZ water should rehydrate tissues better than ordinary water because of its higher dipole moment. To appreciate this argument, picture a bean with positive charge localized at one end, negative at the other. The positive end of that dipole orients toward the negatively charged cell, which then strongly draws in that dipole. The larger the dipole, the stronger will be the draw. Since EZs contain masses of separated charges, or large dipoles, EZ water should hydrate cells better than ordinary water. That’s why EZ water may particularly promote good health.
Negative Charge and Anti-Oxidants
Humans are considered neutral, but I suggest that we bear net negative charge.
Physical chemists reasonably presume that all systems tend toward neutrality because positive charge attracts negative charge. The human body being one of those “systems,” we assume that the body must be neutral.
Not all systems are neutral, however. The earth bears net negative charge, while the atmosphere bears net positive charge. Water itself can bear charge: Anyone watching MIT professor Walter Lewin’s stunning demonstration of the Kelvin water dropper, where separated bodies of water eventually discharge onto one another, will immediately see that bodies of water can bear net charge. If any doubt remains, then the experience of getting an electric shock from touching certain kinds of drinking water (which my colleagues and I have personally experienced) should eliminate that doubt.
Charges can remain separated if input energy keeps them separated — something like recharging your cell phone battery and creating separated negative and positive terminals. Since we constantly absorb external energy from the environment, the theoretical possibility exists that we may bear net charge.
Consider the arithmetic. Cells make up some 60% of your body’s mass, and they are negatively charged. Extracellular tissues such as collagen and elastin are next in line, and those proteins bear negative charge and adsorb negatively charged EZ water. Only some of the smaller compartments are positively charged with protons (low pH), and they commonly expel: urine, gastrointestinal system; sweat, and expired air (containing hydrated CO2 or carbonic acid). They help rid the body of positive charge.
So, the arithmetic shows not only that our body bears net negative charge, but also that the body makes every effort to maintain that negativity by ridding itself of protons. It is as though maintaining negativity is a “goal” of life. Plants do it easily: they connect directly to the negatively charged earth; animals need to struggle a bit more to maintain their body’s charge, in exchange for greater mobility.
How does our body’s negative charge relate to the benefits of anti-oxidants?
Answering this question returns us to basic chemistry. Recall that “reduction” is the gain of electrons, while “oxidation” means electron loss. Oxidation strips molecules of their negative charge, working against the body’s attempt to maintain high negativity. To guard against that loss we employ anti-oxidants. Anti-oxidants may keep us healthy simply by maintaining proper negativity.
The Future
Water’s centrality for health is nothing new, but it has been progressively forgotten. With the various sciences laying emphasis [on] molecular, atomic, and even sub-atomic approaches, we have lost sight of what happens when the pieces come together to form the larger entity. The whole may indeed exceed the sum of its parts. 99% of those parts are water molecules. To think that 99% of our molecules merely bathe the “more important” molecules of life ignores centuries of evidence to the contrary. Water plays a central role in all features of life.
Until recently, the understanding of water’s properties has been constrained by the common misconception that water has three phases. We now know it has four. Taking into account this fourth phase allows many of water’s “anomalies” to vanish: those anomalies turn into predictable features. Water becomes more understandable, and so do entities made largely of water, such as oceans, clouds, and human beings.
Various hour-long talks describe these fresh understandings. One of them is a University of Washington public award lecture:
Water, Energy and Life: Fresh Views From the Water’s Edge]
Another was delivered more recently:
Electrically Structured Water, Part 1
A third is a recent TEDx talk:
The Fourth Phase of Water: Dr. Gerald Pollack at TEDxGuelphU
The insights described above arose out of a departure from mainstream science. They were gleaned mainly from simple observations and logical interpretations. I have purposefully ignored the “generally accepted,” with some skepticism that all accepted principles are necessarily valid. I believe this skepticism has brought us a long way.
Water Isn’t What You Think It Is: The Fourth Phase of Water by Gerald Pollack
Guest author Gerald Pollack introduces a fundamental shift in how we view water. It has the potential to significantly alter our understandings of any processes that involve water, including aspects of climate, biology, and how we approach eco-restoration.
The Fourth Phase of Water: Beyond Solid, Liquid, and Vapor
How can a Jesus Christ lizard walk on water? Why do pollen grains jitterbug in a puddle? Why do fair weather clouds form such lovely puffy white shapes? Why do your joints work without squeaking? Why do sprained ankles swell within seconds?
Answering these questions requires an understanding of water. Given water’s simplicity and pervasiveness through nature, we presume that water must be completely understood, but in fact, precious little is known about how water molecules line up — until recently.
Students learn that water has three phases: solid, liquid and vapor. But there is something more: in our laboratory at the University of Washington we have uncovered a fourth phase. This phase occurs next to water loving (hydrophilic) surfaces. It is surprisingly extensive, projecting out from surfaces by up to millions of molecular layers. And it exists almost everywhere throughout nature, including your body.
This freshly identified phase of water has been described in a recent book: The Fourth Phase of Water: Beyond Solid, Liquid and Vaporwww.ebnerandsons.com. The book documents the basic findings and presents many applications including the ones mentioned above. It also deals with water’s many anomalies, turning those anomalies into easily explained features.
The existence of a fourth phase may seem unexpected. However, it should not be entirely so: a century ago, the physical chemist Sir William Hardy argued for the existence of a fourth phase; and many authors over the years have found evidence for some kind of “ordered” or “structured” phase of water. Fresh experimental evidence not only confirms the existence of such an ordered, liquid-crystalline phase, but also details its properties. Those properties explain everyday observations and answer questions ranging from why gelatin desserts hold their water, to why teapots whistle.
The presence of the fourth phase carries many implications. Here, I outline some basic features of this phase, and then deal with several of those implications. I will touch on atmospheric science, and then focus on some biological and health applications.
Does Water Transduce Energy?
The energy for building water structure comes from the sun. Radiant energy converts ordinary bulk water into ordered water, building this ordered zone. We found that all wavelengths ranging from UV through visible to infrared can build this ordered water. Near-infrared energy is the most capable. Water absorbs infrared energy freely from the environment; it uses that energy to convert bulk water into liquid crystalline water (fourth phase water) — which we also call “exclusion zone” or “EZ” water because it profoundly excludes solutes. Hence, buildup of EZ water occurs naturally and spontaneously from environmental energy. Additional energy input creates additional EZ buildup.
Of particular significance is the fourth phase’s charge: commonly negative (Figure 1). Absorbed radiant energy splits water molecules; the negative moiety constitutes the building block of the EZ, while the positive moiety binds with water molecules to form free hydronium ions, which diffuse throughout the water. Adding additional light stimulates more charge separation.
This process resembles the first step of photosynthesis. In that step, energy from the sun splits water molecules. Hydrophilic chromophores catalyze the splitting. The process considered here is similar but more generic: any hydrophilic surface may catalyze the splitting. Some surfaces work more effectively than others.
The separated charges resemble a battery. That battery can deliver energy in a manner similar to the way the separated charges in plants deliver energy. Plants, of course, comprise mostly water, and it is therefore no surprise that similar energy conversion takes place in water itself.
The stored electrical energy in water can drive various kinds of work, including flow. An example is the axial flow through tubes. We found that immersing tubes made of hydrophilic materials into water produces flow through those tubes, similar to blood flow through blood vessels (Figure 2). The driving energy comes from the radiant energy absorbed and stored in the water. Nothing more. Flow may persist undiminished for many hours, even days. Additional incident light brings faster flow. This is not a perpetual motion machine: incident radiant energy drives the flow — in much the same way that it drives vascular flow in plants. And, we have fresh evidence (see below) that it also assists the heart in driving blood in the cardiovascular system.
Applications in Biological Flow and Atmospheric Science
The water-based energy conversion framework is rich with implication for many systems involving water. These systems may range from biology and chemistry all the way to atmospheric science and engineering. The fourth phase appears nearly everywhere: all that’s needed is water, radiant energy, and a hydrophilic surface. The latter can be as large as a slab of polymer and as small as a dissolved molecule. The liquid crystalline phase inevitably builds — and its presence plays some integral role in the system’s behavior.
Let me provide a few representative examples.
One example is…you. Two thirds of your cells are water — by volume. In terms of the molecular fraction, that fraction translates to more than 99% because so many of those diminutive molecules are required to build that two-thirds volume fraction. Modern cell biology considers that 99% fraction of your molecules as mere background carriers of the “important” molecules of life such as proteins and nucleic acids. Conventional wisdom asserts that 99% of your molecules don’t do very much.
However, EZ water envelops every macromolecule in the cell. Those macromolecules are so tightly packed that the enveloping liquid crystalline water largely fills your cells. In other words, most of your cell water is liquid crystalline, or EZ water. This water plays a central role in everything the cell does — as elaborated in my earlier book, Cells, Gels and the Engines of Lifewww.ebnerandsons.com.
What’s new is the role of radiant energy: incident radiant energy powers many of those cellular functions. An example is the blood flowing through your capillaries. That blood eventually encounters high resistance: capillaries are often narrower than the red blood cells that must pass through them; to make their way through, those red cells need to bend and contort. Resistance is high. You’d anticipate the need for lots of driving pressure; yet, the pressure gradient across the capillary bed is negligible. The paradox resolves if radiant energy helps propel flow through capillaries in the same way that it propels flow through hydrophilic tubes. Radiant energy may constitute an unsuspected source of vascular drive, supplementing cardiac pressure.
Why you feel good after a sauna now seems understandable. If radiant energy drives capillary flow and ample capillary flow is important for optimal functioning, then sitting in the sauna will inevitably be a feel-good experience. The infrared energy associated with heat should help drive that flow. The same if you walk out into sunlight: we presume that the feel-good experience derives purely from the psychological realm; but the evidence above implies that sunlight may build your body’s EZs. Fully built EZs around each protein seem necessary for optimal cellular functioning.
A second example of the EZ’s central role is weather. Common understanding of weather derives from two principal variables: temperature and pressure. Those two variables are said to explain virtually everything we experience in terms of weather. However, the atmosphere also contains water: it is full of micrometer-scale droplets commonly known as aerosol droplets or aerosol particles. Those droplets make up atmospheric humidity. When the atmosphere is humid, the many droplets scatter considerable light, causing haze; you can’t see clearly through that haze. When the atmosphere contains only few droplets, you may see clearly, over long distances.
The Fourth Phase book presents evidence for the structure of those droplets. It shows that EZ water envelops each droplet, while hydronium ions occupy the droplets’ interior. Repelling one another, those internal hydronium ions create pressure, which pushes against the robust shell of EZ water. That explains why droplets tend toward roundness.
How do those aerosol droplets condense to form clouds? The droplets’ EZ shells bear negative charge. Negatively charged droplets should repel one another, precluding any condensation into clouds. Those like-charged aerosol droplets should remain widely dispersed throughout the atmosphere. However, droplets do often condense into clouds, and the question is how that can happen.
The reason they condense is because of the unlike charges that lie in between the droplets. Richard Feynman, the legendary Nobel Prize physicist of the late 20th century understood the principle, opining that: “like-likes-like because of an intermediate of unlikes.” The like-charged droplets “like” one another, so they come together; the unlike charges lying in between those droplets constitute the attractors (Figure 3).
The like-likes-like principle has been widely appreciated, but also widely ignored: after all, how could like charges conceivably attract? A reason why this powerfully simple concept has been ignored is that the source of the unlike charges has been difficult to identify. We now know that the unlike charges can come from the splitting of water — the negative components building EZ shells, while the corresponding positive components provide the unlike attractors. With enough of those attractors, the negatively charged aerosol droplets may condense into clouds.
These two phenomena, radiant energy-induced biological function and like-likes-like cloud formation, provide examples of how water’s energy can account for phenomena not otherwise explained. The fourth phase is the key building block that allows for construction of an edifice of understanding.
Practical Applications
Beyond scientific, the discovery of the fourth phase has practical applications. They include flow production (already mentioned), electrical energy harvesting, and even filtration. I briefly mention the latter two applications.
Filtration occurs naturally because the liquid crystalline phase massively excludes solutes and particles in much the same way as does ice. Accordingly, fourth phase water is essentially solute free. Collecting it provides solute-free and bacteria-free water. A working prototype has confirmed this expectation. Purification by this method requires no physical filter: the fourth phase itself does the separation, and the energy comes from the sun.
Energy harvesting seems straightforward: light drives the separation of charge, and those separated charges constitute a battery. Harvesting electrical energy should be realizable with proper electrodes. This technology development is underway in our spinoff company, and has the potential to replace standard photovoltaic systems with simpler ones based on water. More detail on these practical applications can be found in the Pollack laboratory homepage: <http://faculty.washington.edu/ghp/>.
Practical applications also exist within our bodies, and I present two of them: why your joints don’t squeak: and why dislocated or sprained joints will swell within seconds.
Joints are sites at which bones press upon one another (Figure 4). The bones may also rotate, as during deep-knee bends and push-ups. You’d think that rotation under pressure might elicit squeaky frictional resistance, but joint friction remains remarkably modest. Why so?
The ends of bones are lined with cartilage. Those cartilaginous materials do the actual pressing. Hence, the issue of joint friction reduces to the issue of the cartilaginous surfaces and the synovial fluid lying in between. How does this system behave under pressure?
Cartilage is made of classic gel materials: highly charged polymers and water; therefore, cartilage is a gel. Gel surfaces bear EZs, so cartilage surfaces should likewise bear EZs. The splitting of water associated with EZ buildup creates many hydronium ions in the synovial fluid between. Additional hydronium ions come from the molecules within that fluid, creating their own EZs and protons. Thus, many hydronium ions will lie in the area where two cartilaginous surfaces lie across from one another. The repulsive force coming from those hydronium ions should keep the cartilage surfaces apart — some investigators maintain that despite heavy loads, the cartilage surfaces never touch. That separation means that any rough spots, or asperities, will never come into contact as the respective surfaces shear past one another; and that in turn means low friction.
For such a mechanism to work, some kind of built-in restraint should be present to keep the repelling hydronium ions in place. Otherwise, they may be forced out of the local region, compromising lubrication. Nature provides that safety net: a structure known as the joint capsule envelops the joint. By constraining the dispersal of hydronium ions, that encapsulation ensures low friction. That’s why your joints don’t ordinarily squeak.
Regarding swelling, the second issue under consideration, osmosis evidently plays a role. Since the cell is packed with negatively charged proteins, the cytoplasm should generate an osmotic draw similar to the osmotic draw generated by diapers or gels. Physiologists know that it does.
A peculiar feature of cells, however, is their relatively modest water content. Compared to 20:1 or higher for many common gels, the cell’s water-to-solids ratio is only about 2:1. The many negatively charged macromolecules of the cell should generate a strong osmotic draw; yet the water content in the cell remains surprisingly low. That limited water content may come as a consequence of the macromolecular network’s stiffness: cellular networks typically comprise tubular or multi-stranded biopolymers tightly cross-linked to one another. The resultant stiffness prevents the network from expanding to its full osmotic potential.
If those cross-links were to disrupt, however, then the full power of osmotic draw would take effect; the tissue could then build many EZ layers and therefore hydrate massively, bringing huge expansion (Figure 5). That’s what happens when body tissues are injured, especially with dislocations. The injury disrupts fibrous macromolecules and cross-links, eliminating the restraining forces that keep osmosis at bay; EZ buildup can then proceed virtually unimpeded.
The reason why swelling can be so impressive is that the cross-link disruption occurs progressively. Breaking one cross-link results in higher stress on neighboring cross-links; so disruption progresses in a zipper-like fashion. When that happens, the osmotic rush of water into the tissue can continue practically without restraint, resulting in the enormous immediate swelling that is often seen. The tissue will return to normal only when cross-links repair and the matrix returns to its normally restraining configuration.
Water and Healing
During childhood illness, grandmothers and doctors will often advise: “drink more water.” In his now-classical book, sub-titled Your Body’s Many Cries for Water: You Are Not Sick, You Are Thirsty, the Iranian physician Fereydoon Batmanghelidj confirms the wisdom of this quaint advice. The author documents years of clinical practice showing reversal of diverse pathologies simply by drinking more water. Hydration is critical.
Batmanghelidj’s experience meshes with evidence of healing from special waters such as those from the Ganges and Lourdes. Those waters most often come from deep underground springs or from glacial melt. Spring waters experience pressure from above; pressure converts liquid water into EZ water because of EZ water’s higher density. So, spring water’s healing quality may arise not only from its mineral content but also from its relatively high EZ content.
The same for mountain water: it too should have high EZ content. Our studies have shown that ice formation requires an EZ intermediate; i.e., bulk water does not convert directly to ice; it converts to EZ, which then converts to ice. Similarly for melting: melting ice forms EZ, which subsequently converts to bulk water. Fresh ice melt contains abundant EZ water.
For spring water and fresh ice melt, then, the high EZ content may explain the recognized health benefits. EZ water should rehydrate tissues better than ordinary water because of its higher dipole moment. To appreciate this argument, picture a bean with positive charge localized at one end, negative at the other. The positive end of that dipole orients toward the negatively charged cell, which then strongly draws in that dipole. The larger the dipole, the stronger will be the draw. Since EZs contain masses of separated charges, or large dipoles, EZ water should hydrate cells better than ordinary water. That’s why EZ water may particularly promote good health.
Negative Charge and Anti-Oxidants
Humans are considered neutral, but I suggest that we bear net negative charge.
Physical chemists reasonably presume that all systems tend toward neutrality because positive charge attracts negative charge. The human body being one of those “systems,” we assume that the body must be neutral.
Not all systems are neutral, however. The earth bears net negative charge, while the atmosphere bears net positive charge. Water itself can bear charge: Anyone watching MIT professor Walter Lewin’s stunning demonstration of the Kelvin water dropper, where separated bodies of water eventually discharge onto one another, will immediately see that bodies of water can bear net charge. If any doubt remains, then the experience of getting an electric shock from touching certain kinds of drinking water (which my colleagues and I have personally experienced) should eliminate that doubt.
Charges can remain separated if input energy keeps them separated — something like recharging your cell phone battery and creating separated negative and positive terminals. Since we constantly absorb external energy from the environment, the theoretical possibility exists that we may bear net charge.
Consider the arithmetic. Cells make up some 60% of your body’s mass, and they are negatively charged. Extracellular tissues such as collagen and elastin are next in line, and those proteins bear negative charge and adsorb negatively charged EZ water. Only some of the smaller compartments are positively charged with protons (low pH), and they commonly expel: urine, gastrointestinal system; sweat, and expired air (containing hydrated CO2 or carbonic acid). They help rid the body of positive charge.
So, the arithmetic shows not only that our body bears net negative charge, but also that the body makes every effort to maintain that negativity by ridding itself of protons. It is as though maintaining negativity is a “goal” of life. Plants do it easily: they connect directly to the negatively charged earth; animals need to struggle a bit more to maintain their body’s charge, in exchange for greater mobility.
How does our body’s negative charge relate to the benefits of anti-oxidants?
Answering this question returns us to basic chemistry. Recall that “reduction” is the gain of electrons, while “oxidation” means electron loss. Oxidation strips molecules of their negative charge, working against the body’s attempt to maintain high negativity. To guard against that loss we employ anti-oxidants. Anti-oxidants may keep us healthy simply by maintaining proper negativity.
The Future
Water’s centrality for health is nothing new, but it has been progressively forgotten. With the various sciences laying emphasis [on] molecular, atomic, and even sub-atomic approaches, we have lost sight of what happens when the pieces come together to form the larger entity. The whole may indeed exceed the sum of its parts. 99% of those parts are water molecules. To think that 99% of our molecules merely bathe the “more important” molecules of life ignores centuries of evidence to the contrary. Water plays a central role in all features of life.
Until recently, the understanding of water’s properties has been constrained by the common misconception that water has three phases. We now know it has four. Taking into account this fourth phase allows many of water’s “anomalies” to vanish: those anomalies turn into predictable features. Water becomes more understandable, and so do entities made largely of water, such as oceans, clouds, and human beings.
Various hour-long talks describe these fresh understandings. One of them is a University of Washington public award lecture http://www.youtube.com/watch?v=XVBEwn6iWOo [Water, Energy and Life: Fresh Views From the Water’s Edge]. Another was delivered more recently http://www.youtube.com/watch?v=JnGCMQ8TJ_g [Electrically Structured Water, Part 1]. A third is a recent TEDx talk http://youtu.be/i-T7tCMUDXU [The Fourth Phase of Water: Dr. Gerald Pollack at TEDxGuelphU].
A much fuller, well-referenced understanding of these phenomena and more appears in the above-mentioned new book, The Fourth Phase of Water: Beyond Solid, Liquid, and Vapor <www.ebnerandsons.com>.
The insights described above arose out of a departure from mainstream science. They were gleaned mainly from simple observations and logical interpretations. I have purposefully ignored the “generally accepted,” with some skepticism that all accepted principles are necessarily valid. I believe this skepticism has brought us a long way.
Picture California in the 1700s, around the time the first Spanish missions appeared. It must have looked like heaven on earth for the 100,000s of native people living there [Ecological Society of America 2014], cradled between forested mountains and sparkling ocean. Meandering streams and rivers teeming with salmon criss-cross the valley and are knit together by a latticework of beaver dams. These porous little dams spread water over floodplains stretching in every direction, topping off aquifers and creating diverse habitat for an abundance of life. Wildfires come and go, and some are even manipulated by native peoples to diversify the resources in the landscape. But fires burn through relatively small patches before being damped out by ample ambient moisture, while triggering new growth in their wake.
This old-time paradise lacks the cozy thrill of watching a good Hollywood movie with a box of buttery popcorn, or the sense of security from a Central Valley harvest bountiful enough to nourish a nation. Indeed, our current era boasts a different kind of paradise – one that features convenient access to entertainment, great food, elegantly decorated homes and exotic vacations. This modern paradise is only occasionally interrupted by massive wildfires or storms that level whole neighborhoods, taps that run dry or are tainted with poison, deadly heat waves, or by news reports of such events. Those without the means to live in modern paradise are often the ones featured in the disturbing news reports.
Now imagine California in 2050, after ecosystems have been restored and deployed to rehydrate the landscape and to give Mother Nature the elbowroom she needs to go about her business without doing quite as much damage to human habitats, in spite of ongoing climate chaos. While we can’t and may not even want to go back to a pre-colonial natural California paradise, we can shore up our human spaces against the ravages of climate change by rehabilitating ecosystem processes in all the open space inside of and surrounding our existing cities and towns.
This means preventing any further urban sprawl into rural/wilderness areas. It means relocating people out of flood and fire prone rural/wilderness edges (as the Netherlands has done in its most low-lying areas [Bentley 2016]), and moving them into refurbished urban lots, while fully supporting them through the transition. It means thoughtfully investing in cities and towns to make them affordable and beautiful to reduce people’s compulsion to move into the fringes.
It means reconnecting rivers to their floodplains so that water has somewhere to go other than surging into cities when it floods, and so that floodplains, their wild inhabitants, and the underlying aquifers can benefit from a recharge of groundwater and nutrients. It means helping farmers transition to agricultural methods that conserve water, recycle nutrients, and restore biodiversity to their fields. And it means seeking out the best in applied ecological science to restore and manage wilderness areas in a way that favors biodiversity and therefore resilience.
Like the growing global call to reduce carbon emissions to net zero by 2025, the prospect of vast and thorough ecosystems restoration sounds utopian, naïve, impossible. However, serious solutions are needed to manage ever more extreme weather. Hotter, drier conditions wrought by climate change have made the land crisp and flammable, causing wildfires to spread further and faster in recent years. A century of wildfire suppression allowing fuel build-up, urban sprawl butting up against fire-prone wilderness areas, and the spread of invasive flammable cheatgrass have further contributed to the severity of recent fires.
Moreover, the frequency of rainy seasons that are extremely dry is predicted to increase, especially in southern California, where consecutive dry years will become more common [Swain 2018]. At the same time, the frequency of major flooding events, comparable to the 1862 flood that temporarily turned the Sacramento Valley into an inland sea, is predicted to triple by the end of this century.
Water – whether too much or too little – is at the heart of California’s troubles. With a Mediterranean climate defined by a winter rainy season and long dry summers, California is inherently prone to feast or famine when it comes to water. State water managers, therefore, have long been dealt the challenge of capturing and storing winter rainfall for summer use. Water storage today is achieved with reservoirs, underground aquifers and mountain snowpack, which slowly releases water from melting snow during spring and summer.
The state’s water management system may be due for a redesign, though. For one thing, snowpack is no longer a reliable storage system. Not only is there less snowpack overall, but what there is bears a greater risk now of “rain on snow” events that melt it all at once in a giant gush. Furthermore, levees are not high enough for the scale of flooding that is predicted to become more common, and the water infrastructure overall is in need of repair to the tune of $34 billion [Mount 2017].
The state’s aquifers are a natural storage system that contribute about one third to annual water supply, but they have been unregulated and severely overdrawn during droughts, in some cases leading to land subsidence[10] [Martin 2018]. In response, however, the government passed the Sustainable Groundwater Management Act in 2014, which makes local governments responsible for monitoring and recharging the aquifers within their jurisdictions.
Localities can fulfil their legal obligations at least in part by restoring natural processes that recharge groundwater, such as giving rivers more space to meander and restoring native vegetation. Plants protect and build soil, rendering the soil more sponge-like and able to absorb and hold water, allowing water to percolate into aquifers. Vegetation also shades soil surfaces, limiting evaporation. These principles hold true for all healthy ecosystems, which is to say biodiverse ecosystems, whether in the context of wilderness or agriculture. Biodiversity fosters ecosystem stability, productivity and resilience.
At this turning point for water management in California, what if the state were to redesign its aging system in a way that places a much higher value on hydrological functionality? Improving the land’s hydrology would mean facilitating the ground’s absorption of precipitation (such as the Yolo Bypass Wilderness Area has done), slowing water down rather than isolating it from the land with straight, narrow and constrained waterways ultimately emptying into the ocean.
Slowing down water would recharge aquifers and revitalize streams, rivers, wetlands, grasslands and forests. Expansive functional forested ecosystems could in turn reactivate local water cycling through transpiration and rain recycling, cooling the land in the process. Moistening and cooling the land by enhancing its absorptive capacity could create fire breaks and reduce flammability, thus mitigating the threat of out-of-control wildfires.
Moistening and cooling the land by enhancing its absorptive capacity could create fire breaks and reduce flammability, thus mitigating the threat of out-of-control wildfires.
Disturbed, dry hydrological systems are associated with more severe wildfires. In Borneo, researchers [Taufik 2017] found that massive wildfires coincided with years characterized by large areas of hydrological drought, where surface or groundwater levels had dropped due to extended rainfall deficits. Land use changes including deforestation and canalization to drain wetlands exacerbated hydrological drought, leading to more acres burned during wildfires.
How to restore ecosystems? On a small scale, Californians are already succeeding in this by removing levees to reactivate the floodplain [Fountain 2018], introducing beavers to restore streams and surrounding habitat [Goldfarb 2018],[11] and adaptively grazing livestock in wilderness areas to thin out vegetation, thus reducing fuel loads and replacing it with soil-nourishing manure and urine [Greenwood 2018]. Farmers in the Yolo Bypass Wilderness Area have adapted their practices to allow for seasonal flooding of their fields [Sommer 2001]. Farmers throughout the state participate in the department of agriculture’s Healthy Soils Program by implementing conservation practices to build soil and enhance ecosystem processes, such as carbon sequestration.
Urban areas too are learning to manage water in a way that better withstands boom or bust rainfall patterns. While cities everywhere are normally designed to remove stormwater as quickly as possible, some cities are beginning to see stormwater not so much as a nuisance, but as a valuable resource [Shimabuku, Diringer, Cooley 2018]. For example, the small agricultural town of Gonzalez, CA, modified its municipal code to facilitate low-impact development measures, such as the removal of portions of curb to allow rainwater to drain into vegetated areas. Santa Monica set a city-wide goal of becoming water self-sufficient by 2022 and plans to meet this goal, in part, by capturing more than 500 million gallons of stormwater for treatment and reuse. San Francisco now requires new developments of a certain size to capture and reuse stormwater on site.
San Francisco also recently passed a bill to allow denser development near transit lines within the city, potentially alleviating the housing crisis that’s driving sprawl. (A similar bill was defeated at the state level, unfortunately, due in part to a NIMBY-like resistance to sharing more spacious urban neighborhoods with lower-income newcomers.)
All these initiatives are a promising start. To stave off ever-longer fire seasons, alongside more severe flooding and drought, however, these innovations cannot remain isolated examples. They need to become the state-wide norm for water management, urban design and land stewardship. Such changes are urgently needed not only in California, profiled here as an instructive example of life in the era of climate change, but everywhere.
As in every community everywhere on Earth, California must take an honest, clear look at its choices: reimagine paradise as a place where we embrace the vibrancy of human diversity in dense, walkable urban neighborhoods, and cultivate biodiversity in all open green spaces; or tempt Mother Nature to destroy everything we love as we cling to an ideal of paradise as being the accumulation of manufactured luxuries, while ignoring the environmental and social costs.
Close up on California in the era of climate change: a verdant vision for fire-prone land
Picture California in the 1700s, around the time the first Spanish missions appeared. It must have looked like heaven on earth for the 100,000s of native people living there [Ecological Society of America 2014], cradled between forested mountains and sparkling ocean. Meandering streams and rivers teeming with salmon criss-cross the valley and are knit together by a latticework of beaver dams. These porous little dams spread water over floodplains stretching in every direction, topping off aquifers and creating diverse habitat for an abundance of life. Wildfires come and go, and some are even manipulated by native peoples to diversify the resources in the landscape. But fires burn through relatively small patches before being damped out by ample ambient moisture, while triggering new growth in their wake.
This old-time paradise lacks the cozy thrill of watching a good Hollywood movie with a box of buttery popcorn, or the sense of security from a Central Valley harvest bountiful enough to nourish a nation. Indeed, our current era boasts a different kind of paradise – one that features convenient access to entertainment, great food, elegantly decorated homes and exotic vacations. This modern paradise is only occasionally interrupted by massive wildfires or storms that level whole neighborhoods, taps that run dry or are tainted with poison, deadly heat waves, or by news reports of such events. Those without the means to live in modern paradise are often the ones featured in the disturbing news reports.
Now imagine California in 2050, after ecosystems have been restored and deployed to rehydrate the landscape and to give Mother Nature the elbowroom she needs to go about her business without doing quite as much damage to human habitats, in spite of ongoing climate chaos. While we can’t and may not even want to go back to a pre-colonial natural California paradise, we can shore up our human spaces against the ravages of climate change by rehabilitating ecosystem processes in all the open space inside of and surrounding our existing cities and towns.
This means preventing any further urban sprawl into rural/wilderness areas. It means relocating people out of flood and fire prone rural/wilderness edges (as the Netherlands has done in its most low-lying areas [Bentley 2016]), and moving them into refurbished urban lots, while fully supporting them through the transition. It means thoughtfully investing in cities and towns to make them affordable and beautiful to reduce people’s compulsion to move into the fringes.
It means reconnecting rivers to their floodplains so that water has somewhere to go other than surging into cities when it floods, and so that floodplains, their wild inhabitants, and the underlying aquifers can benefit from a recharge of groundwater and nutrients. It means helping farmers transition to agricultural methods that conserve water, recycle nutrients, and restore biodiversity to their fields. And it means seeking out the best in applied ecological science to restore and manage wilderness areas in a way that favors biodiversity and therefore resilience.
Like the growing global call to reduce carbon emissions to net zero by 2025, the prospect of vast and thorough ecosystems restoration sounds utopian, naïve, impossible. However, serious solutions are needed to manage ever more extreme weather. Hotter, drier conditions wrought by climate change have made the land crisp and flammable, causing wildfires to spread further and faster in recent years. A century of wildfire suppression allowing fuel build-up, urban sprawl butting up against fire-prone wilderness areas, and the spread of invasive flammable cheatgrass have further contributed to the severity of recent fires.
Moreover, the frequency of rainy seasons that are extremely dry is predicted to increase, especially in southern California, where consecutive dry years will become more common [Swain 2018]. At the same time, the frequency of major flooding events, comparable to the 1862 flood that temporarily turned the Sacramento Valley into an inland sea, is predicted to triple by the end of this century.
Water – whether too much or too little – is at the heart of California’s troubles. With a Mediterranean climate defined by a winter rainy season and long dry summers, California is inherently prone to feast or famine when it comes to water. State water managers, therefore, have long been dealt the challenge of capturing and storing winter rainfall for summer use. Water storage today is achieved with reservoirs, underground aquifers and mountain snowpack, which slowly releases water from melting snow during spring and summer.
The state’s water management system may be due for a redesign, though. For one thing, snowpack is no longer a reliable storage system. Not only is there less snowpack overall, but what there is bears a greater risk now of “rain on snow” events that melt it all at once in a giant gush. Furthermore, levees are not high enough for the scale of flooding that is predicted to become more common, and the water infrastructure overall is in need of repair to the tune of $34 billion [Mount 2017].
The state’s aquifers are a natural storage system that contribute about one third to annual water supply, but they have been unregulated and severely overdrawn during droughts, in some cases leading to land subsidence[10] [Martin 2018]. In response, however, the government passed the Sustainable Groundwater Management Act in 2014, which makes local governments responsible for monitoring and recharging the aquifers within their jurisdictions.
Localities can fulfil their legal obligations at least in part by restoring natural processes that recharge groundwater, such as giving rivers more space to meander and restoring native vegetation. Plants protect and build soil, rendering the soil more sponge-like and able to absorb and hold water, allowing water to percolate into aquifers. Vegetation also shades soil surfaces, limiting evaporation. These principles hold true for all healthy ecosystems, which is to say biodiverse ecosystems, whether in the context of wilderness or agriculture. Biodiversity fosters ecosystem stability, productivity and resilience.
At this turning point for water management in California, what if the state were to redesign its aging system in a way that places a much higher value on hydrological functionality? Improving the land’s hydrology would mean facilitating the ground’s absorption of precipitation (such as the Yolo Bypass Wilderness Area has done), slowing water down rather than isolating it from the land with straight, narrow and constrained waterways ultimately emptying into the ocean.
Slowing down water would recharge aquifers and revitalize streams, rivers, wetlands, grasslands and forests. Expansive functional forested ecosystems could in turn reactivate local water cycling through transpiration and rain recycling, cooling the land in the process. Moistening and cooling the land by enhancing its absorptive capacity could create fire breaks and reduce flammability, thus mitigating the threat of out-of-control wildfires.
Moistening and cooling the land by enhancing its absorptive capacity could create fire breaks and reduce flammability, thus mitigating the threat of out-of-control wildfires.
Disturbed, dry hydrological systems are associated with more severe wildfires. In Borneo, researchers [Taufik 2017] found that massive wildfires coincided with years characterized by large areas of hydrological drought, where surface or groundwater levels had dropped due to extended rainfall deficits. Land use changes including deforestation and canalization to drain wetlands exacerbated hydrological drought, leading to more acres burned during wildfires.
How to restore ecosystems? On a small scale, Californians are already succeeding in this by removing levees to reactivate the floodplain [Fountain 2018], introducing beavers to restore streams and surrounding habitat [Goldfarb 2018],[11] and adaptively grazing livestock in wilderness areas to thin out vegetation, thus reducing fuel loads and replacing it with soil-nourishing manure and urine [Greenwood 2018]. Farmers in the Yolo Bypass Wilderness Area have adapted their practices to allow for seasonal flooding of their fields [Sommer 2001]. Farmers throughout the state participate in the department of agriculture’s Healthy Soils Program by implementing conservation practices to build soil and enhance ecosystem processes, such as carbon sequestration.
Urban areas too are learning to manage water in a way that better withstands boom or bust rainfall patterns. While cities everywhere are normally designed to remove stormwater as quickly as possible, some cities are beginning to see stormwater not so much as a nuisance, but as a valuable resource [Shimabuku, Diringer, Cooley 2018]. For example, the small agricultural town of Gonzalez, CA, modified its municipal code to facilitate low-impact development measures, such as the removal of portions of curb to allow rainwater to drain into vegetated areas. Santa Monica set a city-wide goal of becoming water self-sufficient by 2022 and plans to meet this goal, in part, by capturing more than 500 million gallons of stormwater for treatment and reuse. San Francisco now requires new developments of a certain size to capture and reuse stormwater on site.
San Francisco also recently passed a bill to allow denser development near transit lines within the city, potentially alleviating the housing crisis that’s driving sprawl. (A similar bill was defeated at the state level, unfortunately, due in part to a NIMBY-like resistance to sharing more spacious urban neighborhoods with lower-income newcomers.)
All these initiatives are a promising start. To stave off ever-longer fire seasons, alongside more severe flooding and drought, however, these innovations cannot remain isolated examples. They need to become the state-wide norm for water management, urban design and land stewardship. Such changes are urgently needed not only in California, profiled here as an instructive example of life in the era of climate change, but everywhere.
As in every community everywhere on Earth, California must take an honest, clear look at its choices: reimagine paradise as a place where we embrace the vibrancy of human diversity in dense, walkable urban neighborhoods, and cultivate biodiversity in all open green spaces; or tempt Mother Nature to destroy everything we love as we cling to an ideal of paradise as being the accumulation of manufactured luxuries, while ignoring the environmental and social costs.
This article highlights the relevance of the concept of “sustainable intensification” (SI), wherein farming practices are improved to produce more crops (intensification) while doing no harm to – and possibly even enhancing – the environment (sustainable).
The combination of the two terms was an attempt to indicate that desirable outcomes, such as more food and better ecosystem services, need not be mutually exclusive. Both could be achieved by making better use of land, water, biodiversity, labour, knowledge, and technologies [Pretty 2018: 441].
Having screened hundreds of SI projects/programs worldwide and selected those implemented at a large enough scale to benefit ecosystem services and agricultural objectives across whole landscapes, the authors estimate that 29% of all farms worldwide and 9% of agricultural land have crossed a transformative threshold in their use of SI methods.
Sustainable intensification can be achieved in three non-linear stages of transition: (1) improved efficiency (such as through precision agriculture, which uses sensors to target fertilizer application, thereby wasting less), (2) substitution (such as substituting biological control agents in the place of synthetic inputs), or (3) system redesign. The authors state that of these three stages, only system redesign is capable of “maximizing co-production of both favorable agricultural and environmental outcomes at regional and continental scales” [Pretty 2018: 442].
The authors describe system redesign as follows:
The third stage is a fundamental prerequisite for SI to achieve impact at scale. Redesign centres on the composition and structure of agro-ecosystems to deliver sustainability across all dimensions to facilitate food, fibre and fuel production at increased rates. Redesign harnesses predation, parasitism, allelopathy, herbivory, nitrogen fixation, pollination, trophic dependencies and other agro-ecological processes to develop components that deliver beneficial services for the production of crops and livestock. A prime aim is to influence the impacts of agroecosystem management on externalities (negative and positive), such as greenhouse gas emissions, clean water, carbon sequestration, biodiversity and dispersal of pests, pathogens and weeds. Whereas efficiency and substitution tend to be additive and incremental within current production systems, redesign brings the most transformative changes across systems [Pretty 2018: 442].
We analysed transitions towards redesign in agricultural systems worldwide. We reviewed literature on SI, including meta-analyses and practices, to produce a typology of seven system types that we classify as redesign: (i) integrated pest management, (ii) conservation agriculture, (iii) integrated crop and biodiversity, (iv) pasture and forage, (v) trees in agricultural systems, (vi) irrigation water management and (vii) intensive small and patch systems [Pretty 2018: 443].
By showing that “the expansion of SI has begun to occur at scale across a wide range of agroecosystems” [Pretty 2018: 444], this analysis offers a roadmap for how to transition global agriculture toward systemic ecosystem and community resilience in the face of global warming. The authors emphasize the importance both of social networks and cooperation for the co-creation and sharing of agricultural knowledge, and of state policies to support or at least not undermine SI expansion. The authors suggest that agricultural management may be at a crucial tipping point, where “a further small increase in the number of farms successfully operating re-designed agricultural systems could lead rapidly to re-design of agriculture on a global scale” [Pretty 2018: 445].
Although the overall contribution of decaying plants, available substrate, and microbes to the buildup of soil organic matter (SOM) is well recognized, their individual contributions are not as clearly understood. Analytical shortcomings have constrained a thorough study that can distinguish the amount of SOM attributable to plants and the amount attributable to microbes. Using pyrolysis-GC/MS, the authors investigated the chemistry of carbon and microbe-depleted soils after 18 months, after inoculation by various substrates and with two clays (montmorillonite and kaolinite).
By six months, active microbial communities were present in all inocula save one, and SOM molecular diversity increased across all model soil systems. Soils treated with either sugar or syringol (a structural component of cell walls) contained substantial concentrations of lipids and proteins after 18 months. Soil organic carbon (SOC) also increased over time. Syringol-treated montmorillonite soils accumulated the most carbon, and had lower bacterial and higher fungal abundance than sugar-treated soils. Higher fungal abundance was positively correlated with carbon use efficiency (CUE) across treatments.
The authors concluded that the microbial community may be a stronger driver of SOM development than the soil’s mineralogy. They also found that their sugar- and syringol-treated samples provided a chemical diversity that was as rich as natural soil. This article contributes to an expanding scientific knowledge base regarding soil ecosystems and the critical role of soil microorganisms vis a vis the carbon cycle.
Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls, Kallenbach et al. 2016
Although the overall contribution of decaying plants, available substrate, and microbes to the buildup of soil organic matter (SOM) is well recognized, their individual contributions are not as clearly understood. Analytical shortcomings have constrained a thorough study that can distinguish the amount of SOM attributable to plants and the amount attributable to microbes. Using pyrolysis-GC/MS, the authors investigated the chemistry of carbon and microbe-depleted soils after 18 months, after inoculation by various substrates and with two clays (montmorillonite and kaolinite).
By six months, active microbial communities were present in all inocula save one, and SOM molecular diversity increased across all model soil systems. Soils treated with either sugar or syringol (a structural component of cell walls) contained substantial concentrations of lipids and proteins after 18 months. Soil organic carbon (SOC) also increased over time. Syringol-treated montmorillonite soils accumulated the most carbon, and had lower bacterial and higher fungal abundance than sugar-treated soils. Higher fungal abundance was positively correlated with carbon use efficiency (CUE) across treatments.
The authors concluded that the microbial community may be a stronger driver of SOM development than the soil’s mineralogy. They also found that their sugar- and syringol-treated samples provided a chemical diversity that was as rich as natural soil. This article contributes to an expanding scientific knowledge base regarding soil ecosystems and the critical role of soil microorganisms vis a vis the carbon cycle.
Global assessment of agricultural system redesign for sustainable intensification, Pretty et al. 2018
This article highlights the relevance of the concept of “sustainable intensification” (SI), wherein farming practices are improved to produce more crops (intensification) while doing no harm to – and possibly even enhancing – the environment (sustainable).
The combination of the two terms was an attempt to indicate that desirable outcomes, such as more food and better ecosystem services, need not be mutually exclusive. Both could be achieved by making better use of land, water, biodiversity, labour, knowledge, and technologies [Pretty 2018: 441].
Having screened hundreds of SI projects/programs worldwide and selected those implemented at a large enough scale to benefit ecosystem services and agricultural objectives across whole landscapes, the authors estimate that 29% of all farms worldwide and 9% of agricultural land have crossed a transformative threshold in their use of SI methods.
Sustainable intensification can be achieved in three non-linear stages of transition: (1) improved efficiency (such as through precision agriculture, which uses sensors to target fertilizer application, thereby wasting less), (2) substitution (such as substituting biological control agents in the place of synthetic inputs), or (3) system redesign. The authors state that of these three stages, only system redesign is capable of “maximizing co-production of both favorable agricultural and environmental outcomes at regional and continental scales” [Pretty 2018: 442].
The authors describe system redesign as follows:
The third stage is a fundamental prerequisite for SI to achieve impact at scale. Redesign centres on the composition and structure of agro-ecosystems to deliver sustainability across all dimensions to facilitate food, fibre and fuel production at increased rates. Redesign harnesses predation, parasitism, allelopathy, herbivory, nitrogen fixation, pollination, trophic dependencies and other agro-ecological processes to develop components that deliver beneficial services for the production of crops and livestock. A prime aim is to influence the impacts of agroecosystem management on externalities (negative and positive), such as greenhouse gas emissions, clean water, carbon sequestration, biodiversity and dispersal of pests, pathogens and weeds. Whereas efficiency and substitution tend to be additive and incremental within current production systems, redesign brings the most transformative changes across systems [Pretty 2018: 442].
We analysed transitions towards redesign in agricultural systems worldwide. We reviewed literature on SI, including meta-analyses and practices, to produce a typology of seven system types that we classify as redesign: (i) integrated pest management, (ii) conservation agriculture, (iii) integrated crop and biodiversity, (iv) pasture and forage, (v) trees in agricultural systems, (vi) irrigation water management and (vii) intensive small and patch systems [Pretty 2018: 443].
By showing that “the expansion of SI has begun to occur at scale across a wide range of agroecosystems” [Pretty 2018: 444], this analysis offers a roadmap for how to transition global agriculture toward systemic ecosystem and community resilience in the face of global warming. The authors emphasize the importance both of social networks and cooperation for the co-creation and sharing of agricultural knowledge, and of state policies to support or at least not undermine SI expansion. The authors suggest that agricultural management may be at a crucial tipping point, where “a further small increase in the number of farms successfully operating re-designed agricultural systems could lead rapidly to re-design of agriculture on a global scale” [Pretty 2018: 445].
This study examines how subordinate species[8] influence community insurance against drought in semi-natural grasslands of the Swiss Jura.The insurance hypothesis proposes that an increase in community diversity corresponds to an increase in the range of potential species responses to environmental stress. The authors tested the role of subordinate species in community resistance to drought, recovery and resilience, and on productivity. They induced summer drought conditions for two months by covering the test plants with raincovers.
The drought simulation reduced soil water content by 67%, relative to comparable watered land plots. Drought, removal of subordinate species, and their interaction, all had dramatic adverse impacts on community resistance. In contrast to dominant and transient species, subordinate species showed significantly stronger resistance in drought plots than in control plots. Additional findings supported the conclusion that the plant community was more resistant and produced more biomass after drought when containing high biomass of subordinate plants.
Plant community resilience was not affected by drought but was decreased by the subordinate removal treatment. Species composition was also affected by drought and removal conditions; most dominant and transient species[9] were associated with watered plots. Some transient species (such as the ox-eye daisy) were associated with plots in which subordinate removal had occurred.
The authors conclude that, in general, dominant species fared poorly in response to drought, whereas the proportion of subordinate and transient species increased under these conditions. They also noted that the decline in resistance was about 10 times higher in plots where subordinates had been removed than in plots without removal. Thus, the subordinates facilitated the regrowth of dominants and transients during drought. They proposed that the reduced competition among dominants during drought conditions afforded the subordinates the opportunity to accumulate more biomass.
The authors demonstrate that: “in species-rich grassland communities, subordinate species, a key component of plant diversity, are a main driver of community resistance to drought. Our findings show the importance of ecosystem-level impacts of these low abundant plants” [Mariotte 2013: 771]. They further speculated that the role of subordinates in resisting drought for the whole community may lie in their ability to increase water availability through greater interaction with the soil microbial community, such as mycorrhizal fungi. This article adds credence and specificity to our understanding of the key role of biodiversity in ecosystem functioning.
Our results demonstrate that in the Amazon, forest height and age regulate photosynthesis interannual variability and are as relevant as mean precipitation. In particular, tall, old and dense forests are more resistant to precipitation variability. Tree size and age directly impact forest structure and thus the carbon cycle in the Amazon. This is especially significant given the importance of the Amazon rainforest, not only for the global carbon cycle, but also for global atmospheric circulation, which is closely connected to the evapotranspiration process of this area. Forest height, age and biomass have a role equivalent to mean precipitation in the regulation of forest photosynthesis response to interannual climate variability [Giardina 2018: 4].
This study distinguishes between meteorological droughts (lower than average rainfall) and hydrological droughts, where rainfall shortage has eventually led to surface or groundwater levels falling, to predict area burnt from wildfires. By contrast, most studies consider only climate data when predicting wildfire, yet “these overlook subsurface processes leading to hydrological drought, an important driver” [Taufik 2017: 428].
The authors hypothesize that periods with low groundwater recharge will create conditions for a greater area burnt. They found that massive wildfires in Borneo over the past two decades coincided with years when there were large areas of hydrological drought.
Statistical modelling evidence shows amplifying wildfires and greater area burnt in response to El Niño/Southern Oscillation (ENSO) strength, when hydrology is considered. [Taufik 2017: 428]
Hydrological drought stems from a lack of rain, but also depends on the ability of the land to store water. Thus, land use can exacerbate a hydrological drought.
Human activities through land-use change and associated drainage and land clearing immediately following deforestation or long fallow periods create favourable conditions for the fires and amplify the hydrological drying processes in the aboveground fuels and the underlying organic soil [Taufik 2017: 428].
Human activities through land-use change and associated drainage and land clearing immediately following deforestation or long fallow periods create favourable conditions for the fires and amplify the hydrological drying processes in the aboveground fuels and the underlying organic soil[Taufik 2017: 428].
Wildfires in the West have become larger and more frequent over the past three decades (globally, the length of the fire season increased by 19% from 1979 to 2013) and this trend will continue with global warming. Typical fire prevention strategy, centering on fuel reduction and fire suppression, has proved inadequate. Instead, society must accept the inevitability of fires and reorganize itself accordingly, according to this study. Specifically, an adaptive resilience approach would mean:
(i) recognizing that fuels reduction cannot alter regional wildfire trends; (ii) targeting fuels reduction to increase adaptation by some ecosystems and residential communities to more frequent fire; (iii) actively managing more wild and prescribed fires with a range of severities; and (iv) incentivizing and planning residential development to withstand inevitable wildfire [Schoennagel 2017: 4582].
Between 1990 and 2010, almost 2 million homes were added in the 11 states of the western United States, increasing the WUI [wild-urban interface] area by 24%. Currently, most homes in the WUI are in California (4.5 million), Arizona (1.4 million), and Washington (1 million). Since 1990, the average annual number of structures lost to wildfire has increased by 300%, with a significant step up since 2000. About 15% of the area burned in the western United States since 2000 was within the WUI, including a 2.4-km community protection zone, with the largest proportion of wildfires burning in the WUI zone in California (35%), Colorado (30%), and Washington (24%). Additionally, almost 900,000 residential properties in the western United States, representing a total property value more than $237 billion, are currently at high risk of wildfire damage. Because of the people and property values at risk, WUI fires fundamentally change the tactics and cost of fire suppression as compared with fighting remote fires and account for as much as 95% of suppression costs [Schoennagel 2017: 4583].
There often is a lack of political will to implement policies that incur short-term costs despite their long-term value or to change long-standing policies that are ineffective. For example, few jurisdictions have the will or means to restrict further residential development in the WUI, although modifying and curtailing residential growth in fire-prone lands now would reduce the costs and risks from wildfire in the long term. [Schoennagel 2017: 4585].
…modifying and curtailing residential growth in fire-prone lands now would reduce the costs and risks from wildfire in the long term [Schoennagel 2017: 4585].
Cheatgrass is an introduced annual grass that has spread everywhere throughout the western USA. It is among the first plants to emerge in the spring, after which it completes its life cycle, drying out in summer and thus creating a continuous, dry, fine fuel load across the landscape. This study examined the cheatgrass invasion’s effect on the fire regime of the Great Basin region of the western USA, finding that:
Fires were more likely to start in cheatgrass than in other vegetation types and that cheatgrass is associated with increased fire frequency, size, and duration [Balch 2013: 179-180].
Here, we have documented that cheatgrass-dominated areas, which currently cover ~40,000 km2, sustain increased fire probability compared with native vegetation types. As sites burn, more and more of them are likely to become cheatgrass grasslands thus increasing their future probability of burning. If future climate scenarios hold true, the combination of warmer temperatures and high water availability[7] could yield larger fire events that are carried between forested or shrubland areas by invasive grasses, thus perpetuating a novel grass-fire cycle across the western United States and ultimately reducing cover of woody species [Balch 2013: 182].
In native shrub and grassland ecosystems of the arid western United States, high antecedent precipitation has been shown to be one of the strongest predictors of government-registered burned area (1977–2003), even more so than current-year temperature or drought conditions. The oscillation between wet years that enable substantial grass growth and dry years that desiccate those built-up fuels may create ideal conditions for high fire years, but this hypothesis remains untested for cheatgrass rangelands [Balch 2013: 174].
Fire-driven conversion of shrubland to grassland has been linked to a loss of carbon storage and available soil water [Balch 2013: 174].
In a project funded by National Oceanic and Atmospheric Association (NOAA), about two dozen citizen scientists measured temperatures in Baltimore and Washington DC on two of the hottest days of 2018. By measuring temperatures second by second with thermal sensors while driving prescribed routes through each city, the data collectors revealed a 17-degree temperature gap between the coolest and hottest parts of DC on the same day. The difference? Trees. The well-wooded areas of Natural Arboretum and Rock Creek Park were the coolest parts of the city. The results were similar in Baltimore, where the hottest places were neighborhoods covered in concrete and asphalt with little vegetation. These hotspots were 103 degrees, compared to areas with lots of big trees and parks, which were 16 degrees cooler on the same day.
“Major roadways and dense urban pockets are some of the warmest landscapes in both cities” [NOAA 2018], according to Jeremy Hoffman of the Science Museum of Virginia, one of the lead researchers on the study. “These are areas with little or no vegetation, more asphalt and concrete buildings, which can amplify a heat wave” [NOAA 2018].
Researchers used the data to create heat maps of both cities, which can pinpoint the neighborhoods most vulnerable to dangerous heat waves, and to help city officials identify cooling and resiliency strategies, namely bolstering the quantity and quality of green space, planting new trees and protecting existing trees.
Hot days in the city? It’s all about location, NOAA 2018
In a project funded by National Oceanic and Atmospheric Association (NOAA), about two dozen citizen scientists measured temperatures in Baltimore and Washington DC on two of the hottest days of 2018. By measuring temperatures second by second with thermal sensors while driving prescribed routes through each city, the data collectors revealed a 17-degree temperature gap between the coolest and hottest parts of DC on the same day. The difference? Trees. The well-wooded areas of Natural Arboretum and Rock Creek Park were the coolest parts of the city. The results were similar in Baltimore, where the hottest places were neighborhoods covered in concrete and asphalt with little vegetation. These hotspots were 103 degrees, compared to areas with lots of big trees and parks, which were 16 degrees cooler on the same day.
“Major roadways and dense urban pockets are some of the warmest landscapes in both cities” [NOAA 2018], according to Jeremy Hoffman of the Science Museum of Virginia, one of the lead researchers on the study. “These are areas with little or no vegetation, more asphalt and concrete buildings, which can amplify a heat wave” [NOAA 2018].
Researchers used the data to create heat maps of both cities, which can pinpoint the neighborhoods most vulnerable to dangerous heat waves, and to help city officials identify cooling and resiliency strategies, namely bolstering the quantity and quality of green space, planting new trees and protecting existing trees.
Introduced annual grass increases regional fire activity across the arid western USA (1980–2009), Balch et al. 2013
Cheatgrass is an introduced annual grass that has spread everywhere throughout the western USA. It is among the first plants to emerge in the spring, after which it completes its life cycle, drying out in summer and thus creating a continuous, dry, fine fuel load across the landscape. This study examined the cheatgrass invasion’s effect on the fire regime of the Great Basin region of the western USA, finding that:
Fires were more likely to start in cheatgrass than in other vegetation types and that cheatgrass is associated with increased fire frequency, size, and duration [Balch 2013: 179-180].
Here, we have documented that cheatgrass-dominated areas, which currently cover ~40,000 km2, sustain increased fire probability compared with native vegetation types. As sites burn, more and more of them are likely to become cheatgrass grasslands thus increasing their future probability of burning. If future climate scenarios hold true, the combination of warmer temperatures and high water availability[7] could yield larger fire events that are carried between forested or shrubland areas by invasive grasses, thus perpetuating a novel grass-fire cycle across the western United States and ultimately reducing cover of woody species [Balch 2013: 182].
In native shrub and grassland ecosystems of the arid western United States, high antecedent precipitation has been shown to be one of the strongest predictors of government-registered burned area (1977–2003), even more so than current-year temperature or drought conditions. The oscillation between wet years that enable substantial grass growth and dry years that desiccate those built-up fuels may create ideal conditions for high fire years, but this hypothesis remains untested for cheatgrass rangelands [Balch 2013: 174].
Fire-driven conversion of shrubland to grassland has been linked to a loss of carbon storage and available soil water [Balch 2013: 174].
Adapt to more wildfire in western North American forests as climate changes, Schoennagel et al. 2017
Wildfires in the West have become larger and more frequent over the past three decades (globally, the length of the fire season increased by 19% from 1979 to 2013) and this trend will continue with global warming. Typical fire prevention strategy, centering on fuel reduction and fire suppression, has proved inadequate. Instead, society must accept the inevitability of fires and reorganize itself accordingly, according to this study. Specifically, an adaptive resilience approach would mean:
(i) recognizing that fuels reduction cannot alter regional wildfire trends; (ii) targeting fuels reduction to increase adaptation by some ecosystems and residential communities to more frequent fire; (iii) actively managing more wild and prescribed fires with a range of severities; and (iv) incentivizing and planning residential development to withstand inevitable wildfire [Schoennagel 2017: 4582].
Between 1990 and 2010, almost 2 million homes were added in the 11 states of the western United States, increasing the WUI [wild-urban interface] area by 24%. Currently, most homes in the WUI are in California (4.5 million), Arizona (1.4 million), and Washington (1 million). Since 1990, the average annual number of structures lost to wildfire has increased by 300%, with a significant step up since 2000. About 15% of the area burned in the western United States since 2000 was within the WUI, including a 2.4-km community protection zone, with the largest proportion of wildfires burning in the WUI zone in California (35%), Colorado (30%), and Washington (24%). Additionally, almost 900,000 residential properties in the western United States, representing a total property value more than $237 billion, are currently at high risk of wildfire damage. Because of the people and property values at risk, WUI fires fundamentally change the tactics and cost of fire suppression as compared with fighting remote fires and account for as much as 95% of suppression costs [Schoennagel 2017: 4583].
There often is a lack of political will to implement policies that incur short-term costs despite their long-term value or to change long-standing policies that are ineffective. For example, few jurisdictions have the will or means to restrict further residential development in the WUI, although modifying and curtailing residential growth in fire-prone lands now would reduce the costs and risks from wildfire in the long term. [Schoennagel 2017: 4585].
…modifying and curtailing residential growth in fire-prone lands now would reduce the costs and risks from wildfire in the long term [Schoennagel 2017: 4585].
Amplification of wildfire area burnt by hydrological drought in the humid tropics, Taufik et al. 2017
This study distinguishes between meteorological droughts (lower than average rainfall) and hydrological droughts, where rainfall shortage has eventually led to surface or groundwater levels falling, to predict area burnt from wildfires. By contrast, most studies consider only climate data when predicting wildfire, yet “these overlook subsurface processes leading to hydrological drought, an important driver” [Taufik 2017: 428].
The authors hypothesize that periods with low groundwater recharge will create conditions for a greater area burnt. They found that massive wildfires in Borneo over the past two decades coincided with years when there were large areas of hydrological drought.
Statistical modelling evidence shows amplifying wildfires and greater area burnt in response to El Niño/Southern Oscillation (ENSO) strength, when hydrology is considered. [Taufik 2017: 428]
Hydrological drought stems from a lack of rain, but also depends on the ability of the land to store water. Thus, land use can exacerbate a hydrological drought.
Human activities through land-use change and associated drainage and land clearing immediately following deforestation or long fallow periods create favourable conditions for the fires and amplify the hydrological drying processes in the aboveground fuels and the underlying organic soil [Taufik 2017: 428].
Human activities through land-use change and associated drainage and land clearing immediately following deforestation or long fallow periods create favourable conditions for the fires and amplify the hydrological drying processes in the aboveground fuels and the underlying organic soil[Taufik 2017: 428].
Tall Amazonian forests are less sensitive to precipitation variability, Giardina et al. 2018
Our results demonstrate that in the Amazon, forest height and age regulate photosynthesis interannual variability and are as relevant as mean precipitation. In particular, tall, old and dense forests are more resistant to precipitation variability. Tree size and age directly impact forest structure and thus the carbon cycle in the Amazon. This is especially significant given the importance of the Amazon rainforest, not only for the global carbon cycle, but also for global atmospheric circulation, which is closely connected to the evapotranspiration process of this area. Forest height, age and biomass have a role equivalent to mean precipitation in the regulation of forest photosynthesis response to interannual climate variability [Giardina 2018: 4].
Subordinate plant species enhance community resistance against drought in semi-natural grasslands, Mariotte et al. 2013
This study examines how subordinate species[8] influence community insurance against drought in semi-natural grasslands of the Swiss Jura.The insurance hypothesis proposes that an increase in community diversity corresponds to an increase in the range of potential species responses to environmental stress. The authors tested the role of subordinate species in community resistance to drought, recovery and resilience, and on productivity. They induced summer drought conditions for two months by covering the test plants with raincovers.
The drought simulation reduced soil water content by 67%, relative to comparable watered land plots. Drought, removal of subordinate species, and their interaction, all had dramatic adverse impacts on community resistance. In contrast to dominant and transient species, subordinate species showed significantly stronger resistance in drought plots than in control plots. Additional findings supported the conclusion that the plant community was more resistant and produced more biomass after drought when containing high biomass of subordinate plants.
Plant community resilience was not affected by drought but was decreased by the subordinate removal treatment. Species composition was also affected by drought and removal conditions; most dominant and transient species[9] were associated with watered plots. Some transient species (such as the ox-eye daisy) were associated with plots in which subordinate removal had occurred.
The authors conclude that, in general, dominant species fared poorly in response to drought, whereas the proportion of subordinate and transient species increased under these conditions. They also noted that the decline in resistance was about 10 times higher in plots where subordinates had been removed than in plots without removal. Thus, the subordinates facilitated the regrowth of dominants and transients during drought. They proposed that the reduced competition among dominants during drought conditions afforded the subordinates the opportunity to accumulate more biomass.
The authors demonstrate that: “in species-rich grassland communities, subordinate species, a key component of plant diversity, are a main driver of community resistance to drought. Our findings show the importance of ecosystem-level impacts of these low abundant plants” [Mariotte 2013: 771]. They further speculated that the role of subordinates in resisting drought for the whole community may lie in their ability to increase water availability through greater interaction with the soil microbial community, such as mycorrhizal fungi. This article adds credence and specificity to our understanding of the key role of biodiversity in ecosystem functioning.
Politicians often call for logging and fuel reduction to prevent future wildfires. However, it’s not good logging trees that are burning in such fires so much as cheatgrass, annual weed, dry brush and dead weeds. Reintroducing beaver to create ponds could raise the water table, increase humidity in the drainage area (thus reducing burn intensity) and provide a refuge for animals during a fire.
Beavers are recognized for their ability to restore floodplain hydrology and biological function, yet finding suitable places for their reintroduction remains a conservation challenge. The goal of this study was to identify places in the Snohomish River basin of Washington state suitable for beaver reintroduction.
Because of their abilities to modify streams and floodplains, beavers have the potential to play a critical role in shaping how riparian and stream ecosystems respond to climate change. The Pacific Northwest of the United States is experiencing increases in annual air temperature and decreases in snow pack and summer precipitation, resulting in lower base flows, particularly in streams that rely on late season snowmelt. Climate shifts have altered stream-temperature regimes to the detriment of cold-water fishes, including Pacific salmon. Recent increases in winter precipitation and storm magnitude have increased the potential for stream scour, channel incision, and floodplain disconnection, thereby promoting the drying of adjacent riparian areas [Dittbrenner 2018: 2].
By damming streams, beavers create pond and wetland complexes that increase spatial heterogeneity and geomorphic complexity, species and habitat diversity, and therefore ecosystem resilience to climate-induced environmental change. Beaver impoundments slow stream velocity allowing sediment suspended in the water column to settle, aggrading incised stream systems, and reconnecting streams with their floodplains. The increase in surface water promotes groundwater recharge, storage, and supplementation during base flows. The increased geomorphic complexity also promotes higher thermal variability and coldwater refugia in deeper waters and in areas of downstream upwelling [Dittbrenner 2018: 2].
To qualify as a suitable site for beaver reintroduction, a site needs to be intrinsically suitable beaver habitat and clear of competing human interests.
Of 5,019 stream km assessed in this study, just 33% had moderate or high intrinsic potential for beaver habitat. “Of the riparian areas around streams with high intrinsic potential for beaver, 38% are on public lands and 17% are on large tracts of privately-owned timber land” [Dittbrenner 2018: 1], while the rest was on human-dominated landscapes (agricultural, industrial, residential, etc.). Thus, the areas available for beaver reintroduction are limited. Even so, the authors argue that beavers can play a critical role in adapting to climate change, and they propose that watersheds dominated by public ownership, “provide ample opportunities to test how beavers can be reintroduced into landscapes where they are absent or at low population levels” [Dittbrenner 2018: 11].
This study provides empirical evidence that beavers influence hydrologic processes in riparian areas. Conducted at the headwaters of the Colorado River in the Rocky Mountains, the study examines patterns from two beaver dams of surface inundation, groundwater flow, and groundwater level dynamics. The authors observe that :
Beaver dams on the Colorado River caused river water to move around them as surface runoff and groundwater seepage during both high- and low-flow periods. The beaver dams attenuated the expected water table decline in the drier summer months for 9 and 12 ha of the 58 ha study area [Westbrook 2006: 1] … by providing a constant supply of water to the riparian area via surface and subsurface flow paths [Westbrook 2006: 10].
In both cases [both dams], water left the Colorado River, flowed across the floodplain and terrace, and then back to the river far downstream of the dams [Westbrook 2006: 11].
Noting that the current beaver population is but a small fraction of what it was before Europeans settled the west, the authors state that:
If the results of our intensive study were extrapolated to a time of more abundant beaver then the magnitude of their hydrologic effects may have encompassed nearly the entire study area. It is easy to visualize abundant beaver as key drivers of hydrologic processes in mountain valleys and other unconfined stream valleys throughout North America [Westbrook 2006: 10].
The significance of this study is that beaver dams can maintain the water table in forests, creating resilience to drought. Beaver dams do this by causing water to overflow the banks of the river and spread over a greater surface area. More effective even than any given rain event, “overbank flood events have generally been regarded as the main hydrologic mechanism for replenishing groundwater and soil water in riparian areas” [Westbrook 2006: 8].
The significance of this study is that beaver dams can maintain the water table in forests, creating resilience to drought. Beaver dams do this by causing water to overflow the banks of the river and spread over a greater surface area.
Beaver dams and overbank floods influence groundwater–surface water interactions of a Rocky Mountain riparian area, Westbrook et al. 2006
This study provides empirical evidence that beavers influence hydrologic processes in riparian areas. Conducted at the headwaters of the Colorado River in the Rocky Mountains, the study examines patterns from two beaver dams of surface inundation, groundwater flow, and groundwater level dynamics. The authors observe that :
Beaver dams on the Colorado River caused river water to move around them as surface runoff and groundwater seepage during both high- and low-flow periods. The beaver dams attenuated the expected water table decline in the drier summer months for 9 and 12 ha of the 58 ha study area [Westbrook 2006: 1] … by providing a constant supply of water to the riparian area via surface and subsurface flow paths [Westbrook 2006: 10].
In both cases [both dams], water left the Colorado River, flowed across the floodplain and terrace, and then back to the river far downstream of the dams [Westbrook 2006: 11].
Noting that the current beaver population is but a small fraction of what it was before Europeans settled the west, the authors state that:
If the results of our intensive study were extrapolated to a time of more abundant beaver then the magnitude of their hydrologic effects may have encompassed nearly the entire study area. It is easy to visualize abundant beaver as key drivers of hydrologic processes in mountain valleys and other unconfined stream valleys throughout North America [Westbrook 2006: 10].
The significance of this study is that beaver dams can maintain the water table in forests, creating resilience to drought. Beaver dams do this by causing water to overflow the banks of the river and spread over a greater surface area. More effective even than any given rain event, “overbank flood events have generally been regarded as the main hydrologic mechanism for replenishing groundwater and soil water in riparian areas” [Westbrook 2006: 8].
The significance of this study is that beaver dams can maintain the water table in forests, creating resilience to drought. Beaver dams do this by causing water to overflow the banks of the river and spread over a greater surface area.
Modeling intrinsic potential for beaver (Castor canadensis) habitat to inform restoration and climate change adaptation, Dittbrenner et al. 2018
Beavers are recognized for their ability to restore floodplain hydrology and biological function, yet finding suitable places for their reintroduction remains a conservation challenge. The goal of this study was to identify places in the Snohomish River basin of Washington state suitable for beaver reintroduction.
Because of their abilities to modify streams and floodplains, beavers have the potential to play a critical role in shaping how riparian and stream ecosystems respond to climate change. The Pacific Northwest of the United States is experiencing increases in annual air temperature and decreases in snow pack and summer precipitation, resulting in lower base flows, particularly in streams that rely on late season snowmelt. Climate shifts have altered stream-temperature regimes to the detriment of cold-water fishes, including Pacific salmon. Recent increases in winter precipitation and storm magnitude have increased the potential for stream scour, channel incision, and floodplain disconnection, thereby promoting the drying of adjacent riparian areas [Dittbrenner 2018: 2].
By damming streams, beavers create pond and wetland complexes that increase spatial heterogeneity and geomorphic complexity, species and habitat diversity, and therefore ecosystem resilience to climate-induced environmental change. Beaver impoundments slow stream velocity allowing sediment suspended in the water column to settle, aggrading incised stream systems, and reconnecting streams with their floodplains. The increase in surface water promotes groundwater recharge, storage, and supplementation during base flows. The increased geomorphic complexity also promotes higher thermal variability and coldwater refugia in deeper waters and in areas of downstream upwelling [Dittbrenner 2018: 2].
To qualify as a suitable site for beaver reintroduction, a site needs to be intrinsically suitable beaver habitat and clear of competing human interests.
Of 5,019 stream km assessed in this study, just 33% had moderate or high intrinsic potential for beaver habitat. “Of the riparian areas around streams with high intrinsic potential for beaver, 38% are on public lands and 17% are on large tracts of privately-owned timber land” [Dittbrenner 2018: 1], while the rest was on human-dominated landscapes (agricultural, industrial, residential, etc.). Thus, the areas available for beaver reintroduction are limited. Even so, the authors argue that beavers can play a critical role in adapting to climate change, and they propose that watersheds dominated by public ownership, “provide ample opportunities to test how beavers can be reintroduced into landscapes where they are absent or at low population levels” [Dittbrenner 2018: 11].
Beaver restoration would reduce wildfires, Maughan 2013
Politicians often call for logging and fuel reduction to prevent future wildfires. However, it’s not good logging trees that are burning in such fires so much as cheatgrass, annual weed, dry brush and dead weeds. Reintroducing beaver to create ponds could raise the water table, increase humidity in the drainage area (thus reducing burn intensity) and provide a refuge for animals during a fire.
The vulnerability of coastal wetlands to sea-level rise is disputed, with some researchers predicting most will be flooded out of existence by the end of the 21st Century. Coastal wetlands provide critical ecosystem services, including protection from storm surges, water quality improvement, fisheries habitat and carbon sequestration. By accounting for the enhancement of sediment build-up when storms are more frequent and more severe and for the possibility of “accommodation space” for coastal wetlands to move inland, however, these authors reach a more optimistic conclusion. (Sediment build-up, or accretion, allows coastal wetlands to grow vertically, potentially remaining at a higher elevation than sea-level.) They estimate that:
Rather than losses, wetland gains of up to 60 per cent of the current area are possible, if more than 37 per cent (our upper estimate for current accommodation space) of coastal wetlands have sufficient accommodation space, and sediment supply remains at present levels [Schuerch 2018: 231].
This is an important ecosystems restoration message because it means humans can directly influence the persistence of coastal wetlands, and thus the continuation of the essential ecosystem services they provide.
This is an important ecosystems restoration message because it means humans can directly influence the persistence of coastal wetlands, and thus the continuation of the essential ecosystem services they provide.
Our simulations suggest that the resilience of global wetlands is primarily driven by the availability of accommodation space, which is strongly influenced by the building of anthropogenic infrastructure in the coastal zone and such infrastructure is expected to change over the twenty-first century. Rather than being an inevitable consequence of global sea-level rise, our findings indicate that large-scale loss of coastal wetlands might be avoidable, if sufficient additional accommodation space can be created through careful nature-based adaptation solutions to coastal management [Schuerch 2018: 231].
The authors describe specific solutions to protect coastal wetlands, which they recommend be implemented at a large, regional or landscape scale.
Existing nature-based adaptation solutions that allow coastal wetlands to migrate inland include the inland displacement of coastal flood defenses (typically along highly engineered coastlines) or the designation of nature reserve buffers in upland areas surrounding coastal wetlands. These schemes, however, are currently implemented as local-scale projects only; strategically upscaling such projects, for example, as suggested by the shoreline management plans in England and Wales or the coastal master plan in Louisiana, may help coastal wetlands adapt to SLR [sea level rise] at the landscape scale and protect rapidly increasing global coastal populations [Schuerch 2018: 234].
This article emphasizes the global importance of protecting and restoring wetlands in the context of climate change and outlines policy strategies for wetland protection and restoration.
Wetlands play a major though under-appreciated role in climate change mitigation and adaptation. Wetlands enhance local resilience to climate change by providing: “flood storage, buffering of storm damage, protecting water quality by filtering pollutants and sediment out of runoff generated by severe storm events, groundwater recharge and provision of water supply during drought, provision of wildlife refuges and corridors and maintenance of biodiversity” [Moomaw 2018: 192], as well as “direct harvests of fish, animals, and plants” [Moomaw 2018: 188]. Furthermore, wetlands/peatlands store massive amounts of carbon, drawing it out of the atmosphere.
Peatlands and vegetated coastal wetlands are among the most carbon rich sinks on the planet sequestering approximately as much carbon as do global forest ecosystems [Moormaw 2018: 183].
Wetland conditions are critical for C accumulation and storage since decomposition in these systems is limited by a lack of oxygen due to water saturation. Therefore, when plant productivity exceeds decomposition there is an accumulation of soil C. This process eventually develops deep peat deposits, which may accumulate for thousands of years [Moomaw 2018: 187].
By the same token wetlands can become major GHG sources when damaged or destroyed by land use change, fire or climate change.
Altering wetlands can increase the vulnerability of the organic C pool by weakening the self-regulating feedbacks that exist in many peatland systems. Land use change that affects wetland hydrology has had substantial impacts on wetland structure and function. Draining wetlands decreases CO2 uptake and increases rates of microbial decomposition and CO2 release. Soil C is also lost by peat extraction, drainage and other disturbance. The hydrologic changes can be so large that they result in massive losses of C to the atmosphere, such as occurred during the fires in tropical peatlands in Southeast Asia [Moomaw 2018: 187].
Many land-use practices in or near wetlands reduce wetlands’ resilience to any further stress, such as hotter, drier weather wrought by climate change.
Unfortunately, many of the world’s freshwater wetlands are already stressed by increased land-use pressure, so that additional hydrological alteration can contribute to an overall decrease in resilience to climate change. Human alteration is commonplace throughout river corridors, challenging management as the impacts of upstream alterations accumulate along the waterway. As demands for river resources increase, such problems are expected to worsen. Flowing water is compromised by river re-engineering practices, even though moving water generally improves oxygenation and plant health. Also, upriver freshwater extraction in tidal freshwater wetlands coupled with sea level rise can cause the salinification of surface and ground water, with accompanying stress and even the collapse of tidal vegetation in the freshwater reaches of estuaries [Moomaw 2018: 188].
On the other hand, wetland resilience can be bolstered through proper land management.
The effects of climate changes on wetland C storage will be determined largely by the extent to which the wetlands have been modified through land-use change [Moomaw 2018: 187].
One opportunity to decrease the amount of saltmarsh loss that is likely to occur with sea level rise is to actively plan for future inland marsh migration now [Moomaw 2018: 191].
The authors express concern that wetlands are overlooked in policy discussions on climate change, noting that climate scientists tend to sideline the role of wetlands, while wetlands science and management have often failed to acknowledge the outsized role of wetlands as a carbon sink. Thus:
To play a more effective role in climate change mitigation and adaptation/resiliency, wetland scientists need to clearly communicate the significance of wetlands to the wellbeing of society and the economy. Communicating with policy makers and the public requires aligning wetland science and specific climate mitigation and adaptation/resiliency ecosystem services with the concerns and mindset of the audience [Moomaw 2018: 198].
A handful of policy structures at international, national and subnational levels aim to better account for and protect wetlands. For example, the International Panel on Climate Change (IPCC) has since 2013 provided guidance (through the Wetlands Supplement) to countries about including wetlands in national GHG inventories, thus moving “closer to requiring countries to account for the substantial emissions from these ecosystems when they are disturbed or destroyed” [193]. The 1975 Ramsar Agreement establishes an international framework for wetland management, but lacks adequate guidance on how to best protect wetlands from the stressors of climate change. At the local level, decisions about wetlands are often made by land managers.
Thinking globally and acting locally, wetland managers can incorporate carbon management and climate resiliency science into project-level work (including developing a body of climate-related Best Management Practices), whether or not governing policies and regulations exist. As noted earlier in this article, avoidance of impacts to wetlands, and associated carbon stocks and processes, is likely to be the most effective management practice for preventing increases in GHG emissions from wetlands, protecting climate resiliency functions, and protecting traditional wetland ecosystem services, and it is therefore important for managers to understand the underlying science [Moomaw 2018: 197].
The authors of this article note that prior agreements to halt wetland degradation, such as the Ramsar Convention of 1971, have been largely unsuccessful. They advocate for both a re-emphasis on how wetlands help mitigate climate change, and how to protect existing wetlands from the damaging effects of climate change. They had previously authored the Second Warning to Humanity and Wetlands, which urged the Society for Wetland Scientists (SS) and other organizations to raise the profile of wetlands. Doing so can lead to policy changes which would attenuate the deleterious actions that humans currently apply to wetlands.
The authors then provide 11 recommendations for preserving and renewing wetlands. These recommendations include halting the conversion of wetlands to other land uses, rewilding wetlands with native species, and reducing the wastage of wetland-derived foods. Other recommendations are increasing wetland education, adopting renewable energy sources that don’t impact wetlands, prioritizing the enactment of connected, well-funded and well-managed networks of protected wetland areas, and supporting ecologically sound financial investments.
The authors address the lack of high-resolution, large-scale assessments of the value of coastal wetlands for reducing property damages from flooding. In the first part of this paper, they assess Hurricane Sandy-induced damages to wetlands. The second part examines the risk reduction benefits of salt marshes in Ocean County, NJ, in terms of average annual economic flood losses. This study involved over 2000 synthetic storm events in Ocean County. The storm events were matched in frequency with actual storms that occurred between 1900 and 2011.
Wetland extent was positively correlated with damage reduction in all but one of 12 states impacted by Hurricane Sandy. The authors used a hydrodynamic model that calculated the propagation of storm surges from the coastal shelf on to land. The average amount of damage reduction was slightly over 1%; however, four states with extensive wetlands experienced flood damage reduction of 20-30%.
Losses were less for areas with salt marshes than for those without. On average, salt marshes reduced flood-related damages by 18%. Higher elevations were also correlated with damage reductions.
The authors noted that damage reduction was also apparent at locations several kilometers upstream of affected wetlands. A few areas, however, showed increased storm damage because of their proximity to wetlands. These areas often were dammed, or had their stream channel redirected. Based on their findings, the authors advocate for the increased use of flood risk models by the insurance industry and small businesses.
In their natural state, rivers are not separate or separable from surrounding lands. Rather, a river channel is just one integral part of a larger river-floodplain ecosystem. Annual flood pulses and larger flooding events connect river channels to their floodplains, driving the cycles of life for the particularly diverse ensemble of species that live in floodplain ecosystems. For example, fish use floodplain lakes and backwaters for spawning, shelter, feeding and nurseries. Plants on the floodplain depend on nutrients supplied by sediment deposited during flooding. Due to their geological age, size, habitat complexity, and variability, large river ecosystems – such as the Amazon basin – are among the more biodiverse ecosystems on Earth.
Building levees to contain river water eliminates annual flood pulses, thereby fracturing an ecosystem dependent on these processes. Therefore, for example, “in both tropical and temperate rivers, fish yield per acre is considerably greater in rivers with flood pulses and floodplains than in nearby impoundments where flood pulses are reduced or absent” [Sparks 1995: 172]. In addition,
On land, the natural nutrient-replenishment system once provided by the flood must be replaced with commercial fertilizer. Some societies practice a flood-adapted form of agriculture or harvest both fish and a compatible crop, such as rice, but intensive, high-yield agriculture often conflicts with fisheries, particularly if pesticides are used that can contaminate fish through biomagnification [Sparks 1995: 172].
To at least partially reconnect rivers with floodplains, the author recommends modifying existing structures to divert some flow to create or maintain side channels into the floodplains and restore the annual flood pulse.
Human societies tend to value the potential benefits that a landscape might provide in a limited way, adjusting management practices towards desired outputs by maximizing the benefits gained from one or some of the services (often the provision of goods) leading to the loss of multifunctionality and the degradation of natural capital at the expense of human welfare. As a result of this biased valuation, the opportunity costs of biodiversity conservation have been perceived as too high [Schindler 2014: 230].
Using a lens of landscape multifunctionality, this study evaluates 38 potential interventions (ranging from mining, agriculture and residential development to dam removal, natural habitat creation and hiking trail maintenance) in European floodplain ecosystems for their potential to provide multiple ecosystem services (ESS). “Most ESS arise from living organisms and the interaction of biotic and abiotic processes, and refer specifically to the ‘final’ outputs from landscapes that provide benefits to humans” [Schindler 2014: 230].
Each intervention was evaluated to determine whether its effect on a given ecosystem service was positive, negative or neutral. The more ecosystem services an intervention was considered to positively affect (such as pollination, water purification, flood mitigation, providing for farming, fishing, drinking water, or recreation), the greater its contribution to landscape multifunctionality.
Interventions with the most positive effects were related to the creation of natural habitat, dike relocation, lateral floodplain reconnection, creation of channels, oxbows and ponds, whereas the interventions [related to] terrestrial settlement and transportation infrastructure, navigational infrastructure, and intensive forms of agriculture, forestry and fisheries are rather problematic when preserving multifunctionality in floodplains [Schindler 2014: 238].
Thus the authors found that:
Restoration and rehabilitation measures strongly improved the multifunctionality of the landscape and caused win–win situations for enhancing overall ESS supply for all regulation/maintenance and cultural services, but also for provisioning services [Schindler 2014: 242].
In short, a multifunctional approach allows for ecosystem services and goods that we depend on yet often take for granted, such as clean, abundant drinking water, clean air, pollination, and productive wild fish populations, for example, to be considered in economic evaluations of sites and landscapes such as floodplains.
The area of floodplains allowed to perform the natural function of storing and conveying floodwaters must be expanded by strategically removing levees or setting them back from the river. Floodplain reconnection will accomplish three primary objectives: flood-risk reduction, an increase in floodplain goods and services, and resiliency to potential climate change impacts [Opperman 2009: 1487].
Floodplain reconnection reduces flood risk by: (1) replacing vulnerable land uses with flood-tolerant uses, thereby reducing damages, and (2) giving the water somewhere to go, thereby sparing downstream and other nearby communities. Furthermore, by storing and conveying water, floodplains alleviate pressure on upstream dams/reservoirs for flood control and water supply, increasing the resilience of this infrastructure. Finally, by restoring biological activity and diversity, floodplain restoration activates ecosystem services, including carbon sequestration and water quality improvement and groundwater recharge.
The authors note that agricultural lands would be less expensive than densely populated residential areas to reconnect and should be prioritized. Furthermore, agricultural land could remain as such by switching to production of flood-tolerant crops, such as timber and pasture. Furthermore, floodplain reconnection has proven popular among farmers, who requested more than ten times the amount of land be enrolled in a one-time floodplain easement program than the USDA could afford to support with American Recovery and Reinvestment Act funds.
Sustainable floodplains through large-scale reconnection to rivers, Opperman et al. 2009
The area of floodplains allowed to perform the natural function of storing and conveying floodwaters must be expanded by strategically removing levees or setting them back from the river. Floodplain reconnection will accomplish three primary objectives: flood-risk reduction, an increase in floodplain goods and services, and resiliency to potential climate change impacts [Opperman 2009: 1487].
Floodplain reconnection reduces flood risk by: (1) replacing vulnerable land uses with flood-tolerant uses, thereby reducing damages, and (2) giving the water somewhere to go, thereby sparing downstream and other nearby communities. Furthermore, by storing and conveying water, floodplains alleviate pressure on upstream dams/reservoirs for flood control and water supply, increasing the resilience of this infrastructure. Finally, by restoring biological activity and diversity, floodplain restoration activates ecosystem services, including carbon sequestration and water quality improvement and groundwater recharge.
The authors note that agricultural lands would be less expensive than densely populated residential areas to reconnect and should be prioritized. Furthermore, agricultural land could remain as such by switching to production of flood-tolerant crops, such as timber and pasture. Furthermore, floodplain reconnection has proven popular among farmers, who requested more than ten times the amount of land be enrolled in a one-time floodplain easement program than the USDA could afford to support with American Recovery and Reinvestment Act funds.
Multifunctionality of floodplain landscapes: relating management options to ecosystem services, Schindler et al. 2014
Human societies tend to value the potential benefits that a landscape might provide in a limited way, adjusting management practices towards desired outputs by maximizing the benefits gained from one or some of the services (often the provision of goods) leading to the loss of multifunctionality and the degradation of natural capital at the expense of human welfare. As a result of this biased valuation, the opportunity costs of biodiversity conservation have been perceived as too high [Schindler 2014: 230].
Using a lens of landscape multifunctionality, this study evaluates 38 potential interventions (ranging from mining, agriculture and residential development to dam removal, natural habitat creation and hiking trail maintenance) in European floodplain ecosystems for their potential to provide multiple ecosystem services (ESS). “Most ESS arise from living organisms and the interaction of biotic and abiotic processes, and refer specifically to the ‘final’ outputs from landscapes that provide benefits to humans” [Schindler 2014: 230].
Each intervention was evaluated to determine whether its effect on a given ecosystem service was positive, negative or neutral. The more ecosystem services an intervention was considered to positively affect (such as pollination, water purification, flood mitigation, providing for farming, fishing, drinking water, or recreation), the greater its contribution to landscape multifunctionality.
Interventions with the most positive effects were related to the creation of natural habitat, dike relocation, lateral floodplain reconnection, creation of channels, oxbows and ponds, whereas the interventions [related to] terrestrial settlement and transportation infrastructure, navigational infrastructure, and intensive forms of agriculture, forestry and fisheries are rather problematic when preserving multifunctionality in floodplains [Schindler 2014: 238].
Thus the authors found that:
Restoration and rehabilitation measures strongly improved the multifunctionality of the landscape and caused win–win situations for enhancing overall ESS supply for all regulation/maintenance and cultural services, but also for provisioning services [Schindler 2014: 242].
In short, a multifunctional approach allows for ecosystem services and goods that we depend on yet often take for granted, such as clean, abundant drinking water, clean air, pollination, and productive wild fish populations, for example, to be considered in economic evaluations of sites and landscapes such as floodplains.
Need for ecosystem management of large rivers and their floodplains: these phenomenally productive ecosystems produce fish and wildlife and preserve species, Sparks 1995
In their natural state, rivers are not separate or separable from surrounding lands. Rather, a river channel is just one integral part of a larger river-floodplain ecosystem. Annual flood pulses and larger flooding events connect river channels to their floodplains, driving the cycles of life for the particularly diverse ensemble of species that live in floodplain ecosystems. For example, fish use floodplain lakes and backwaters for spawning, shelter, feeding and nurseries. Plants on the floodplain depend on nutrients supplied by sediment deposited during flooding. Due to their geological age, size, habitat complexity, and variability, large river ecosystems – such as the Amazon basin – are among the more biodiverse ecosystems on Earth.
Building levees to contain river water eliminates annual flood pulses, thereby fracturing an ecosystem dependent on these processes. Therefore, for example, “in both tropical and temperate rivers, fish yield per acre is considerably greater in rivers with flood pulses and floodplains than in nearby impoundments where flood pulses are reduced or absent” [Sparks 1995: 172]. In addition,
On land, the natural nutrient-replenishment system once provided by the flood must be replaced with commercial fertilizer. Some societies practice a flood-adapted form of agriculture or harvest both fish and a compatible crop, such as rice, but intensive, high-yield agriculture often conflicts with fisheries, particularly if pesticides are used that can contaminate fish through biomagnification [Sparks 1995: 172].
To at least partially reconnect rivers with floodplains, the author recommends modifying existing structures to divert some flow to create or maintain side channels into the floodplains and restore the annual flood pulse.
The Value of Coastal Wetlands for Flood Damage Reduction in the Northeastern USA, Narayan et al. 2017
The authors address the lack of high-resolution, large-scale assessments of the value of coastal wetlands for reducing property damages from flooding. In the first part of this paper, they assess Hurricane Sandy-induced damages to wetlands. The second part examines the risk reduction benefits of salt marshes in Ocean County, NJ, in terms of average annual economic flood losses. This study involved over 2000 synthetic storm events in Ocean County. The storm events were matched in frequency with actual storms that occurred between 1900 and 2011.
Wetland extent was positively correlated with damage reduction in all but one of 12 states impacted by Hurricane Sandy. The authors used a hydrodynamic model that calculated the propagation of storm surges from the coastal shelf on to land. The average amount of damage reduction was slightly over 1%; however, four states with extensive wetlands experienced flood damage reduction of 20-30%.
Losses were less for areas with salt marshes than for those without. On average, salt marshes reduced flood-related damages by 18%. Higher elevations were also correlated with damage reductions.
The authors noted that damage reduction was also apparent at locations several kilometers upstream of affected wetlands. A few areas, however, showed increased storm damage because of their proximity to wetlands. These areas often were dammed, or had their stream channel redirected. Based on their findings, the authors advocate for the increased use of flood risk models by the insurance industry and small businesses.
The second warning to humanity – providing a context for wetland management and policy, Finlayson et al. 2018
The authors of this article note that prior agreements to halt wetland degradation, such as the Ramsar Convention of 1971, have been largely unsuccessful. They advocate for both a re-emphasis on how wetlands help mitigate climate change, and how to protect existing wetlands from the damaging effects of climate change. They had previously authored the Second Warning to Humanity and Wetlands, which urged the Society for Wetland Scientists (SS) and other organizations to raise the profile of wetlands. Doing so can lead to policy changes which would attenuate the deleterious actions that humans currently apply to wetlands.
The authors then provide 11 recommendations for preserving and renewing wetlands. These recommendations include halting the conversion of wetlands to other land uses, rewilding wetlands with native species, and reducing the wastage of wetland-derived foods. Other recommendations are increasing wetland education, adopting renewable energy sources that don’t impact wetlands, prioritizing the enactment of connected, well-funded and well-managed networks of protected wetland areas, and supporting ecologically sound financial investments.
Wetlands in a changing climate: science, policy and management, Moomaw et al. 2018
This article emphasizes the global importance of protecting and restoring wetlands in the context of climate change and outlines policy strategies for wetland protection and restoration.
Wetlands play a major though under-appreciated role in climate change mitigation and adaptation. Wetlands enhance local resilience to climate change by providing: “flood storage, buffering of storm damage, protecting water quality by filtering pollutants and sediment out of runoff generated by severe storm events, groundwater recharge and provision of water supply during drought, provision of wildlife refuges and corridors and maintenance of biodiversity” [Moomaw 2018: 192], as well as “direct harvests of fish, animals, and plants” [Moomaw 2018: 188]. Furthermore, wetlands/peatlands store massive amounts of carbon, drawing it out of the atmosphere.
Peatlands and vegetated coastal wetlands are among the most carbon rich sinks on the planet sequestering approximately as much carbon as do global forest ecosystems [Moormaw 2018: 183].
Wetland conditions are critical for C accumulation and storage since decomposition in these systems is limited by a lack of oxygen due to water saturation. Therefore, when plant productivity exceeds decomposition there is an accumulation of soil C. This process eventually develops deep peat deposits, which may accumulate for thousands of years [Moomaw 2018: 187].
By the same token wetlands can become major GHG sources when damaged or destroyed by land use change, fire or climate change.
Altering wetlands can increase the vulnerability of the organic C pool by weakening the self-regulating feedbacks that exist in many peatland systems. Land use change that affects wetland hydrology has had substantial impacts on wetland structure and function. Draining wetlands decreases CO2 uptake and increases rates of microbial decomposition and CO2 release. Soil C is also lost by peat extraction, drainage and other disturbance. The hydrologic changes can be so large that they result in massive losses of C to the atmosphere, such as occurred during the fires in tropical peatlands in Southeast Asia [Moomaw 2018: 187].
Many land-use practices in or near wetlands reduce wetlands’ resilience to any further stress, such as hotter, drier weather wrought by climate change.
Unfortunately, many of the world’s freshwater wetlands are already stressed by increased land-use pressure, so that additional hydrological alteration can contribute to an overall decrease in resilience to climate change. Human alteration is commonplace throughout river corridors, challenging management as the impacts of upstream alterations accumulate along the waterway. As demands for river resources increase, such problems are expected to worsen. Flowing water is compromised by river re-engineering practices, even though moving water generally improves oxygenation and plant health. Also, upriver freshwater extraction in tidal freshwater wetlands coupled with sea level rise can cause the salinification of surface and ground water, with accompanying stress and even the collapse of tidal vegetation in the freshwater reaches of estuaries [Moomaw 2018: 188].
On the other hand, wetland resilience can be bolstered through proper land management.
The effects of climate changes on wetland C storage will be determined largely by the extent to which the wetlands have been modified through land-use change [Moomaw 2018: 187].
One opportunity to decrease the amount of saltmarsh loss that is likely to occur with sea level rise is to actively plan for future inland marsh migration now [Moomaw 2018: 191].
The authors express concern that wetlands are overlooked in policy discussions on climate change, noting that climate scientists tend to sideline the role of wetlands, while wetlands science and management have often failed to acknowledge the outsized role of wetlands as a carbon sink. Thus:
To play a more effective role in climate change mitigation and adaptation/resiliency, wetland scientists need to clearly communicate the significance of wetlands to the wellbeing of society and the economy. Communicating with policy makers and the public requires aligning wetland science and specific climate mitigation and adaptation/resiliency ecosystem services with the concerns and mindset of the audience [Moomaw 2018: 198].
A handful of policy structures at international, national and subnational levels aim to better account for and protect wetlands. For example, the International Panel on Climate Change (IPCC) has since 2013 provided guidance (through the Wetlands Supplement) to countries about including wetlands in national GHG inventories, thus moving “closer to requiring countries to account for the substantial emissions from these ecosystems when they are disturbed or destroyed” [193]. The 1975 Ramsar Agreement establishes an international framework for wetland management, but lacks adequate guidance on how to best protect wetlands from the stressors of climate change. At the local level, decisions about wetlands are often made by land managers.
Thinking globally and acting locally, wetland managers can incorporate carbon management and climate resiliency science into project-level work (including developing a body of climate-related Best Management Practices), whether or not governing policies and regulations exist. As noted earlier in this article, avoidance of impacts to wetlands, and associated carbon stocks and processes, is likely to be the most effective management practice for preventing increases in GHG emissions from wetlands, protecting climate resiliency functions, and protecting traditional wetland ecosystem services, and it is therefore important for managers to understand the underlying science [Moomaw 2018: 197].
Future response of global coastal wetlands to sea-level rise, Schuerch et al. 2018
The vulnerability of coastal wetlands to sea-level rise is disputed, with some researchers predicting most will be flooded out of existence by the end of the 21st Century. Coastal wetlands provide critical ecosystem services, including protection from storm surges, water quality improvement, fisheries habitat and carbon sequestration. By accounting for the enhancement of sediment build-up when storms are more frequent and more severe and for the possibility of “accommodation space” for coastal wetlands to move inland, however, these authors reach a more optimistic conclusion. (Sediment build-up, or accretion, allows coastal wetlands to grow vertically, potentially remaining at a higher elevation than sea-level.) They estimate that:
Rather than losses, wetland gains of up to 60 per cent of the current area are possible, if more than 37 per cent (our upper estimate for current accommodation space) of coastal wetlands have sufficient accommodation space, and sediment supply remains at present levels [Schuerch 2018: 231].
This is an important ecosystems restoration message because it means humans can directly influence the persistence of coastal wetlands, and thus the continuation of the essential ecosystem services they provide.
This is an important ecosystems restoration message because it means humans can directly influence the persistence of coastal wetlands, and thus the continuation of the essential ecosystem services they provide.
Our simulations suggest that the resilience of global wetlands is primarily driven by the availability of accommodation space, which is strongly influenced by the building of anthropogenic infrastructure in the coastal zone and such infrastructure is expected to change over the twenty-first century. Rather than being an inevitable consequence of global sea-level rise, our findings indicate that large-scale loss of coastal wetlands might be avoidable, if sufficient additional accommodation space can be created through careful nature-based adaptation solutions to coastal management [Schuerch 2018: 231].
The authors describe specific solutions to protect coastal wetlands, which they recommend be implemented at a large, regional or landscape scale.
Existing nature-based adaptation solutions that allow coastal wetlands to migrate inland include the inland displacement of coastal flood defenses (typically along highly engineered coastlines) or the designation of nature reserve buffers in upland areas surrounding coastal wetlands. These schemes, however, are currently implemented as local-scale projects only; strategically upscaling such projects, for example, as suggested by the shoreline management plans in England and Wales or the coastal master plan in Louisiana, may help coastal wetlands adapt to SLR [sea level rise] at the landscape scale and protect rapidly increasing global coastal populations [Schuerch 2018: 234].
The following articles were selected and summarized by Bio4Climate’s Compendium editors and writers. The purpose of this collection is to highlight the scientific evidence and argumentation showing healthy restored and protected ecosystems as a powerful (albeit under-recognized) tool for managing the weather extremes wrought by climate change.
Floodplains and wetlands: making space for water
Sustainable floodplains through large-scale reconnection to rivers, Opperman et al. 2009
The area of floodplains allowed to perform the natural function of storing and conveying floodwaters must be expanded by strategically removing levees or setting them back from the river. Floodplain reconnection will accomplish three primary objectives: flood-risk reduction, an increase in floodplain goods and services, and resiliency to potential climate change impacts [Opperman 2009: 1487].
Floodplain reconnection reduces flood risk by: (1) replacing vulnerable land uses with flood-tolerant uses, thereby reducing damages, and (2) giving the water somewhere to go, thereby sparing downstream and other nearby communities. Furthermore, by storing and conveying water, floodplains alleviate pressure on upstream dams/reservoirs for flood control and water supply, increasing the resilience of this infrastructure. Finally, by restoring biological activity and diversity, floodplain restoration activates ecosystem services, including carbon sequestration and water quality improvement and groundwater recharge.
The authors note that agricultural lands would be less expensive than densely populated residential areas to reconnect and should be prioritized. Furthermore, agricultural land could remain as such by switching to production of flood-tolerant crops, such as timber and pasture. Furthermore, floodplain reconnection has proven popular among farmers, who requested more than ten times the amount of land be enrolled in a one-time floodplain easement program than the USDA could afford to support with American Recovery and Reinvestment Act funds.
Multifunctionality of floodplain landscapes: relating management options to ecosystem services, Schindler et al. 2014
Human societies tend to value the potential benefits that a landscape might provide in a limited way, adjusting management practices towards desired outputs by maximizing the benefits gained from one or some of the services (often the provision of goods) leading to the loss of multifunctionality and the degradation of natural capital at the expense of human welfare. As a result of this biased valuation, the opportunity costs of biodiversity conservation have been perceived as too high [Schindler 2014: 230].
Using a lens of landscape multifunctionality, this study evaluates 38 potential interventions (ranging from mining, agriculture and residential development to dam removal, natural habitat creation and hiking trail maintenance) in European floodplain ecosystems for their potential to provide multiple ecosystem services (ESS). “Most ESS arise from living organisms and the interaction of biotic and abiotic processes, and refer specifically to the ‘final’ outputs from landscapes that provide benefits to humans” [Schindler 2014: 230].
Each intervention was evaluated to determine whether its effect on a given ecosystem service was positive, negative or neutral. The more ecosystem services an intervention was considered to positively affect (such as pollination, water purification, flood mitigation, providing for farming, fishing, drinking water, or recreation), the greater its contribution to landscape multifunctionality.
Interventions with the most positive effects were related to the creation of natural habitat, dike relocation, lateral floodplain reconnection, creation of channels, oxbows and ponds, whereas the interventions [related to] terrestrial settlement and transportation infrastructure, navigational infrastructure, and intensive forms of agriculture, forestry and fisheries are rather problematic when preserving multifunctionality in floodplains [Schindler 2014: 238].
Thus the authors found that:
Restoration and rehabilitation measures strongly improved the multifunctionality of the landscape and caused win–win situations for enhancing overall ESS supply for all regulation/maintenance and cultural services, but also for provisioning services [Schindler 2014: 242].
In short, a multifunctional approach allows for ecosystem services and goods that we depend on yet often take for granted, such as clean, abundant drinking water, clean air, pollination, and productive wild fish populations, for example, to be considered in economic evaluations of sites and landscapes such as floodplains.
Need for ecosystem management of large rivers and their floodplains: these phenomenally productive ecosystems produce fish and wildlife and preserve species, Sparks 1995
In their natural state, rivers are not separate or separable from surrounding lands. Rather, a river channel is just one integral part of a larger river-floodplain ecosystem. Annual flood pulses and larger flooding events connect river channels to their floodplains, driving the cycles of life for the particularly diverse ensemble of species that live in floodplain ecosystems. For example, fish use floodplain lakes and backwaters for spawning, shelter, feeding and nurseries. Plants on the floodplain depend on nutrients supplied by sediment deposited during flooding. Due to their geological age, size, habitat complexity, and variability, large river ecosystems – such as the Amazon basin – are among the more biodiverse ecosystems on Earth.
Building levees to contain river water eliminates annual flood pulses, thereby fracturing an ecosystem dependent on these processes. Therefore, for example, “in both tropical and temperate rivers, fish yield per acre is considerably greater in rivers with flood pulses and floodplains than in nearby impoundments where flood pulses are reduced or absent” [Sparks 1995: 172]. In addition,
On land, the natural nutrient-replenishment system once provided by the flood must be replaced with commercial fertilizer. Some societies practice a flood-adapted form of agriculture or harvest both fish and a compatible crop, such as rice, but intensive, high-yield agriculture often conflicts with fisheries, particularly if pesticides are used that can contaminate fish through biomagnification [Sparks 1995: 172].
To at least partially reconnect rivers with floodplains, the author recommends modifying existing structures to divert some flow to create or maintain side channels into the floodplains and restore the annual flood pulse.
The Value of Coastal Wetlands for Flood Damage Reduction in the Northeastern USA, Narayan et al. 2017
The authors address the lack of high-resolution, large-scale assessments of the value of coastal wetlands for reducing property damages from flooding. In the first part of this paper, they assess Hurricane Sandy-induced damages to wetlands. The second part examines the risk reduction benefits of salt marshes in Ocean County, NJ, in terms of average annual economic flood losses. This study involved over 2000 synthetic storm events in Ocean County. The storm events were matched in frequency with actual storms that occurred between 1900 and 2011.
Wetland extent was positively correlated with damage reduction in all but one of 12 states impacted by Hurricane Sandy. The authors used a hydrodynamic model that calculated the propagation of storm surges from the coastal shelf on to land. The average amount of damage reduction was slightly over 1%; however, four states with extensive wetlands experienced flood damage reduction of 20-30%.
Losses were less for areas with salt marshes than for those without. On average, salt marshes reduced flood-related damages by 18%. Higher elevations were also correlated with damage reductions.
The authors noted that damage reduction was also apparent at locations several kilometers upstream of affected wetlands. A few areas, however, showed increased storm damage because of their proximity to wetlands. These areas often were dammed, or had their stream channel redirected. Based on their findings, the authors advocate for the increased use of flood risk models by the insurance industry and small businesses.
The second warning to humanity – providing a context for wetland management and policy, Finlayson et al. 2018
The authors of this article note that prior agreements to halt wetland degradation, such as the Ramsar Convention of 1971, have been largely unsuccessful. They advocate for both a re-emphasis on how wetlands help mitigate climate change, and how to protect existing wetlands from the damaging effects of climate change. They had previously authored the Second Warning to Humanity and Wetlands, which urged the Society for Wetland Scientists (SS) and other organizations to raise the profile of wetlands. Doing so can lead to policy changes which would attenuate the deleterious actions that humans currently apply to wetlands.
The authors then provide 11 recommendations for preserving and renewing wetlands. These recommendations include halting the conversion of wetlands to other land uses, rewilding wetlands with native species, and reducing the wastage of wetland-derived foods. Other recommendations are increasing wetland education, adopting renewable energy sources that don’t impact wetlands, prioritizing the enactment of connected, well-funded and well-managed networks of protected wetland areas, and supporting ecologically sound financial investments.
Wetlands in a changing climate: science, policy and management, Moomaw et al. 2018
This article emphasizes the global importance of protecting and restoring wetlands in the context of climate change and outlines policy strategies for wetland protection and restoration.
Wetlands play a major though under-appreciated role in climate change mitigation and adaptation. Wetlands enhance local resilience to climate change by providing: “flood storage, buffering of storm damage, protecting water quality by filtering pollutants and sediment out of runoff generated by severe storm events, groundwater recharge and provision of water supply during drought, provision of wildlife refuges and corridors and maintenance of biodiversity” [Moomaw 2018: 192], as well as “direct harvests of fish, animals, and plants” [Moomaw 2018: 188]. Furthermore, wetlands/peatlands store massive amounts of carbon, drawing it out of the atmosphere.
Peatlands and vegetated coastal wetlands are among the most carbon rich sinks on the planet sequestering approximately as much carbon as do global forest ecosystems [Moormaw 2018: 183].
Wetland conditions are critical for C accumulation and storage since decomposition in these systems is limited by a lack of oxygen due to water saturation. Therefore, when plant productivity exceeds decomposition there is an accumulation of soil C. This process eventually develops deep peat deposits, which may accumulate for thousands of years [Moomaw 2018: 187].
By the same token wetlands can become major GHG sources when damaged or destroyed by land use change, fire or climate change.
Altering wetlands can increase the vulnerability of the organic C pool by weakening the self-regulating feedbacks that exist in many peatland systems. Land use change that affects wetland hydrology has had substantial impacts on wetland structure and function. Draining wetlands decreases CO2 uptake and increases rates of microbial decomposition and CO2 release. Soil C is also lost by peat extraction, drainage and other disturbance. The hydrologic changes can be so large that they result in massive losses of C to the atmosphere, such as occurred during the fires in tropical peatlands in Southeast Asia [Moomaw 2018: 187].
Many land-use practices in or near wetlands reduce wetlands’ resilience to any further stress, such as hotter, drier weather wrought by climate change.
Unfortunately, many of the world’s freshwater wetlands are already stressed by increased land-use pressure, so that additional hydrological alteration can contribute to an overall decrease in resilience to climate change. Human alteration is commonplace throughout river corridors, challenging management as the impacts of upstream alterations accumulate along the waterway. As demands for river resources increase, such problems are expected to worsen. Flowing water is compromised by river re-engineering practices, even though moving water generally improves oxygenation and plant health. Also, upriver freshwater extraction in tidal freshwater wetlands coupled with sea level rise can cause the salinification of surface and ground water, with accompanying stress and even the collapse of tidal vegetation in the freshwater reaches of estuaries [Moomaw 2018: 188].
On the other hand, wetland resilience can be bolstered through proper land management.
The effects of climate changes on wetland C storage will be determined largely by the extent to which the wetlands have been modified through land-use change [Moomaw 2018: 187].
One opportunity to decrease the amount of saltmarsh loss that is likely to occur with sea level rise is to actively plan for future inland marsh migration now [Moomaw 2018: 191].
The authors express concern that wetlands are overlooked in policy discussions on climate change, noting that climate scientists tend to sideline the role of wetlands, while wetlands science and management have often failed to acknowledge the outsized role of wetlands as a carbon sink. Thus:
To play a more effective role in climate change mitigation and adaptation/resiliency, wetland scientists need to clearly communicate the significance of wetlands to the wellbeing of society and the economy. Communicating with policy makers and the public requires aligning wetland science and specific climate mitigation and adaptation/resiliency ecosystem services with the concerns and mindset of the audience [Moomaw 2018: 198].
A handful of policy structures at international, national and subnational levels aim to better account for and protect wetlands. For example, the International Panel on Climate Change (IPCC) has since 2013 provided guidance (through the Wetlands Supplement) to countries about including wetlands in national GHG inventories, thus moving “closer to requiring countries to account for the substantial emissions from these ecosystems when they are disturbed or destroyed” [193]. The 1975 Ramsar Agreement establishes an international framework for wetland management, but lacks adequate guidance on how to best protect wetlands from the stressors of climate change. At the local level, decisions about wetlands are often made by land managers.
Thinking globally and acting locally, wetland managers can incorporate carbon management and climate resiliency science into project-level work (including developing a body of climate-related Best Management Practices), whether or not governing policies and regulations exist. As noted earlier in this article, avoidance of impacts to wetlands, and associated carbon stocks and processes, is likely to be the most effective management practice for preventing increases in GHG emissions from wetlands, protecting climate resiliency functions, and protecting traditional wetland ecosystem services, and it is therefore important for managers to understand the underlying science [Moomaw 2018: 197].
Future response of global coastal wetlands to sea-level rise, Schuerch et al. 2018
The vulnerability of coastal wetlands to sea-level rise is disputed, with some researchers predicting most will be flooded out of existence by the end of the 21st Century. Coastal wetlands provide critical ecosystem services, including protection from storm surges, water quality improvement, fisheries habitat and carbon sequestration. By accounting for the enhancement of sediment build-up when storms are more frequent and more severe and for the possibility of “accommodation space” for coastal wetlands to move inland, however, these authors reach a more optimistic conclusion. (Sediment build-up, or accretion, allows coastal wetlands to grow vertically, potentially remaining at a higher elevation than sea-level.) They estimate that:
Rather than losses, wetland gains of up to 60 per cent of the current area are possible, if more than 37 per cent (our upper estimate for current accommodation space) of coastal wetlands have sufficient accommodation space, and sediment supply remains at present levels [Schuerch 2018: 231].
This is an important ecosystems restoration message because it means humans can directly influence the persistence of coastal wetlands, and thus the continuation of the essential ecosystem services they provide.
This is an important ecosystems restoration message because it means humans can directly influence the persistence of coastal wetlands, and thus the continuation of the essential ecosystem services they provide.
Our simulations suggest that the resilience of global wetlands is primarily driven by the availability of accommodation space, which is strongly influenced by the building of anthropogenic infrastructure in the coastal zone and such infrastructure is expected to change over the twenty-first century. Rather than being an inevitable consequence of global sea-level rise, our findings indicate that large-scale loss of coastal wetlands might be avoidable, if sufficient additional accommodation space can be created through careful nature-based adaptation solutions to coastal management [Schuerch 2018: 231].
The authors describe specific solutions to protect coastal wetlands, which they recommend be implemented at a large, regional or landscape scale.
Existing nature-based adaptation solutions that allow coastal wetlands to migrate inland include the inland displacement of coastal flood defenses (typically along highly engineered coastlines) or the designation of nature reserve buffers in upland areas surrounding coastal wetlands. These schemes, however, are currently implemented as local-scale projects only; strategically upscaling such projects, for example, as suggested by the shoreline management plans in England and Wales or the coastal master plan in Louisiana, may help coastal wetlands adapt to SLR [sea level rise] at the landscape scale and protect rapidly increasing global coastal populations [Schuerch 2018: 234].
Partnering with beavers to restore ecosystems
Beaver dams and overbank floods influence groundwater–surface water interactions of a Rocky Mountain riparian area, Westbrook et al. 2006
This study provides empirical evidence that beavers influence hydrologic processes in riparian areas. Conducted at the headwaters of the Colorado River in the Rocky Mountains, the study examines patterns from two beaver dams of surface inundation, groundwater flow, and groundwater level dynamics. The authors observe that :
Beaver dams on the Colorado River caused river water to move around them as surface runoff and groundwater seepage during both high- and low-flow periods. The beaver dams attenuated the expected water table decline in the drier summer months for 9 and 12 ha of the 58 ha study area [Westbrook 2006: 1] … by providing a constant supply of water to the riparian area via surface and subsurface flow paths [Westbrook 2006: 10].
In both cases [both dams], water left the Colorado River, flowed across the floodplain and terrace, and then back to the river far downstream of the dams [Westbrook 2006: 11].
Noting that the current beaver population is but a small fraction of what it was before Europeans settled the west, the authors state that:
If the results of our intensive study were extrapolated to a time of more abundant beaver then the magnitude of their hydrologic effects may have encompassed nearly the entire study area. It is easy to visualize abundant beaver as key drivers of hydrologic processes in mountain valleys and other unconfined stream valleys throughout North America [Westbrook 2006: 10].
The significance of this study is that beaver dams can maintain the water table in forests, creating resilience to drought. Beaver dams do this by causing water to overflow the banks of the river and spread over a greater surface area. More effective even than any given rain event, “overbank flood events have generally been regarded as the main hydrologic mechanism for replenishing groundwater and soil water in riparian areas” [Westbrook 2006: 8].
The significance of this study is that beaver dams can maintain the water table in forests, creating resilience to drought. Beaver dams do this by causing water to overflow the banks of the river and spread over a greater surface area.
Modeling intrinsic potential for beaver (Castor canadensis) habitat to inform restoration and climate change adaptation, Dittbrenner et al. 2018
Beavers are recognized for their ability to restore floodplain hydrology and biological function, yet finding suitable places for their reintroduction remains a conservation challenge. The goal of this study was to identify places in the Snohomish River basin of Washington state suitable for beaver reintroduction.
Because of their abilities to modify streams and floodplains, beavers have the potential to play a critical role in shaping how riparian and stream ecosystems respond to climate change. The Pacific Northwest of the United States is experiencing increases in annual air temperature and decreases in snow pack and summer precipitation, resulting in lower base flows, particularly in streams that rely on late season snowmelt. Climate shifts have altered stream-temperature regimes to the detriment of cold-water fishes, including Pacific salmon. Recent increases in winter precipitation and storm magnitude have increased the potential for stream scour, channel incision, and floodplain disconnection, thereby promoting the drying of adjacent riparian areas [Dittbrenner 2018: 2].
By damming streams, beavers create pond and wetland complexes that increase spatial heterogeneity and geomorphic complexity, species and habitat diversity, and therefore ecosystem resilience to climate-induced environmental change. Beaver impoundments slow stream velocity allowing sediment suspended in the water column to settle, aggrading incised stream systems, and reconnecting streams with their floodplains. The increase in surface water promotes groundwater recharge, storage, and supplementation during base flows. The increased geomorphic complexity also promotes higher thermal variability and coldwater refugia in deeper waters and in areas of downstream upwelling [Dittbrenner 2018: 2].
To qualify as a suitable site for beaver reintroduction, a site needs to be intrinsically suitable beaver habitat and clear of competing human interests.
Of 5,019 stream km assessed in this study, just 33% had moderate or high intrinsic potential for beaver habitat. “Of the riparian areas around streams with high intrinsic potential for beaver, 38% are on public lands and 17% are on large tracts of privately-owned timber land” [Dittbrenner 2018: 1], while the rest was on human-dominated landscapes (agricultural, industrial, residential, etc.). Thus, the areas available for beaver reintroduction are limited. Even so, the authors argue that beavers can play a critical role in adapting to climate change, and they propose that watersheds dominated by public ownership, “provide ample opportunities to test how beavers can be reintroduced into landscapes where they are absent or at low population levels” [Dittbrenner 2018: 11].
Beaver restoration would reduce wildfires, Maughan 2013
Politicians often call for logging and fuel reduction to prevent future wildfires. However, it’s not good logging trees that are burning in such fires so much as cheatgrass, annual weed, dry brush and dead weeds. Reintroducing beaver to create ponds could raise the water table, increase humidity in the drainage area (thus reducing burn intensity) and provide a refuge for animals during a fire.
More ecosystem-oriented considerations for heat wave, drought, flood and fire resilience
Hot days in the city? It’s all about location, NOAA 2018
In a project funded by National Oceanic and Atmospheric Association (NOAA), about two dozen citizen scientists measured temperatures in Baltimore and Washington DC on two of the hottest days of 2018. By measuring temperatures second by second with thermal sensors while driving prescribed routes through each city, the data collectors revealed a 17-degree temperature gap between the coolest and hottest parts of DC on the same day. The difference? Trees. The well-wooded areas of Natural Arboretum and Rock Creek Park were the coolest parts of the city. The results were similar in Baltimore, where the hottest places were neighborhoods covered in concrete and asphalt with little vegetation. These hotspots were 103 degrees, compared to areas with lots of big trees and parks, which were 16 degrees cooler on the same day.
“Major roadways and dense urban pockets are some of the warmest landscapes in both cities” [NOAA 2018], according to Jeremy Hoffman of the Science Museum of Virginia, one of the lead researchers on the study. “These are areas with little or no vegetation, more asphalt and concrete buildings, which can amplify a heat wave” [NOAA 2018].
Researchers used the data to create heat maps of both cities, which can pinpoint the neighborhoods most vulnerable to dangerous heat waves, and to help city officials identify cooling and resiliency strategies, namely bolstering the quantity and quality of green space, planting new trees and protecting existing trees.
Introduced annual grass increases regional fire activity across the arid western USA (1980–2009), Balch et al. 2013
Cheatgrass is an introduced annual grass that has spread everywhere throughout the western USA. It is among the first plants to emerge in the spring, after which it completes its life cycle, drying out in summer and thus creating a continuous, dry, fine fuel load across the landscape. This study examined the cheatgrass invasion’s effect on the fire regime of the Great Basin region of the western USA, finding that:
Fires were more likely to start in cheatgrass than in other vegetation types and that cheatgrass is associated with increased fire frequency, size, and duration [Balch 2013: 179-180].
Here, we have documented that cheatgrass-dominated areas, which currently cover ~40,000 km2, sustain increased fire probability compared with native vegetation types. As sites burn, more and more of them are likely to become cheatgrass grasslands thus increasing their future probability of burning. If future climate scenarios hold true, the combination of warmer temperatures and high water availability[7] could yield larger fire events that are carried between forested or shrubland areas by invasive grasses, thus perpetuating a novel grass-fire cycle across the western United States and ultimately reducing cover of woody species [Balch 2013: 182].
In native shrub and grassland ecosystems of the arid western United States, high antecedent precipitation has been shown to be one of the strongest predictors of government-registered burned area (1977–2003), even more so than current-year temperature or drought conditions. The oscillation between wet years that enable substantial grass growth and dry years that desiccate those built-up fuels may create ideal conditions for high fire years, but this hypothesis remains untested for cheatgrass rangelands [Balch 2013: 174].
Fire-driven conversion of shrubland to grassland has been linked to a loss of carbon storage and available soil water [Balch 2013: 174].
Adapt to more wildfire in western North American forests as climate changes, Schoennagel et al. 2017
Wildfires in the West have become larger and more frequent over the past three decades (globally, the length of the fire season increased by 19% from 1979 to 2013) and this trend will continue with global warming. Typical fire prevention strategy, centering on fuel reduction and fire suppression, has proved inadequate. Instead, society must accept the inevitability of fires and reorganize itself accordingly, according to this study. Specifically, an adaptive resilience approach would mean:
(i) recognizing that fuels reduction cannot alter regional wildfire trends; (ii) targeting fuels reduction to increase adaptation by some ecosystems and residential communities to more frequent fire; (iii) actively managing more wild and prescribed fires with a range of severities; and (iv) incentivizing and planning residential development to withstand inevitable wildfire [Schoennagel 2017: 4582].
Between 1990 and 2010, almost 2 million homes were added in the 11 states of the western United States, increasing the WUI [wild-urban interface] area by 24%. Currently, most homes in the WUI are in California (4.5 million), Arizona (1.4 million), and Washington (1 million). Since 1990, the average annual number of structures lost to wildfire has increased by 300%, with a significant step up since 2000. About 15% of the area burned in the western United States since 2000 was within the WUI, including a 2.4-km community protection zone, with the largest proportion of wildfires burning in the WUI zone in California (35%), Colorado (30%), and Washington (24%). Additionally, almost 900,000 residential properties in the western United States, representing a total property value more than $237 billion, are currently at high risk of wildfire damage. Because of the people and property values at risk, WUI fires fundamentally change the tactics and cost of fire suppression as compared with fighting remote fires and account for as much as 95% of suppression costs [Schoennagel 2017: 4583].
There often is a lack of political will to implement policies that incur short-term costs despite their long-term value or to change long-standing policies that are ineffective. For example, few jurisdictions have the will or means to restrict further residential development in the WUI, although modifying and curtailing residential growth in fire-prone lands now would reduce the costs and risks from wildfire in the long term. [Schoennagel 2017: 4585].
…modifying and curtailing residential growth in fire-prone lands now would reduce the costs and risks from wildfire in the long term [Schoennagel 2017: 4585].
Amplification of wildfire area burnt by hydrological drought in the humid tropics, Taufik et al. 2017
This study distinguishes between meteorological droughts (lower than average rainfall) and hydrological droughts, where rainfall shortage has eventually led to surface or groundwater levels falling, to predict area burnt from wildfires. By contrast, most studies consider only climate data when predicting wildfire, yet “these overlook subsurface processes leading to hydrological drought, an important driver” [Taufik 2017: 428].
The authors hypothesize that periods with low groundwater recharge will create conditions for a greater area burnt. They found that massive wildfires in Borneo over the past two decades coincided with years when there were large areas of hydrological drought.
Statistical modelling evidence shows amplifying wildfires and greater area burnt in response to El Niño/Southern Oscillation (ENSO) strength, when hydrology is considered. [Taufik 2017: 428]
Hydrological drought stems from a lack of rain, but also depends on the ability of the land to store water. Thus, land use can exacerbate a hydrological drought.
Human activities through land-use change and associated drainage and land clearing immediately following deforestation or long fallow periods create favourable conditions for the fires and amplify the hydrological drying processes in the aboveground fuels and the underlying organic soil [Taufik 2017: 428].
Human activities through land-use change and associated drainage and land clearing immediately following deforestation or long fallow periods create favourable conditions for the fires and amplify the hydrological drying processes in the aboveground fuels and the underlying organic soil[Taufik 2017: 428].
Tall Amazonian forests are less sensitive to precipitation variability, Giardina et al. 2018
Our results demonstrate that in the Amazon, forest height and age regulate photosynthesis interannual variability and are as relevant as mean precipitation. In particular, tall, old and dense forests are more resistant to precipitation variability. Tree size and age directly impact forest structure and thus the carbon cycle in the Amazon. This is especially significant given the importance of the Amazon rainforest, not only for the global carbon cycle, but also for global atmospheric circulation, which is closely connected to the evapotranspiration process of this area. Forest height, age and biomass have a role equivalent to mean precipitation in the regulation of forest photosynthesis response to interannual climate variability [Giardina 2018: 4].
Subordinate plant species enhance community resistance against drought in semi-natural grasslands, Mariotte et al. 2013
This study examines how subordinate species[8] influence community insurance against drought in semi-natural grasslands of the Swiss Jura.The insurance hypothesis proposes that an increase in community diversity corresponds to an increase in the range of potential species responses to environmental stress. The authors tested the role of subordinate species in community resistance to drought, recovery and resilience, and on productivity. They induced summer drought conditions for two months by covering the test plants with raincovers.
The drought simulation reduced soil water content by 67%, relative to comparable watered land plots. Drought, removal of subordinate species, and their interaction, all had dramatic adverse impacts on community resistance. In contrast to dominant and transient species, subordinate species showed significantly stronger resistance in drought plots than in control plots. Additional findings supported the conclusion that the plant community was more resistant and produced more biomass after drought when containing high biomass of subordinate plants.
Plant community resilience was not affected by drought but was decreased by the subordinate removal treatment. Species composition was also affected by drought and removal conditions; most dominant and transient species[9] were associated with watered plots. Some transient species (such as the ox-eye daisy) were associated with plots in which subordinate removal had occurred.
The authors conclude that, in general, dominant species fared poorly in response to drought, whereas the proportion of subordinate and transient species increased under these conditions. They also noted that the decline in resistance was about 10 times higher in plots where subordinates had been removed than in plots without removal. Thus, the subordinates facilitated the regrowth of dominants and transients during drought. They proposed that the reduced competition among dominants during drought conditions afforded the subordinates the opportunity to accumulate more biomass.
The authors demonstrate that: “in species-rich grassland communities, subordinate species, a key component of plant diversity, are a main driver of community resistance to drought. Our findings show the importance of ecosystem-level impacts of these low abundant plants” [Mariotte 2013: 771]. They further speculated that the role of subordinates in resisting drought for the whole community may lie in their ability to increase water availability through greater interaction with the soil microbial community, such as mycorrhizal fungi. This article adds credence and specificity to our understanding of the key role of biodiversity in ecosystem functioning.
The 2011 drought in Texas was the worst in recorded history and it lasted until 2015. The ground was so dry that Jonathan Cobb, a 4th generation farmer in Blackland Prairie of central Texas, couldn’t even get crops planted. His 2,500-acre conventional row crop operation was already struggling financially through a treadmill of increasingly more inputs and long hours with little or no yield improvement. So he made the hard decision to leave farming and move to the city.
But before he left, he attended a workshop with renowned soil scientist Ray Archuleta, who focused Jonathan’s attention for the first time on soil health. Archuleta demonstrated the water-holding capacity of healthy, biologically active aggregated soils compared to a typical compacted soil that crumbled and eroded when water was poured on it. This demonstration opened Cobb’s mind to a whole new way of approaching agriculture. It gave him hope that farming even through extreme drought was possible.
There was hope that nature actually did exist on its own before man started cultivating it … things that seem obvious, like a forest ecology that nobody fertilizes a forest … of course it works, it can be very abundant. And so there was a hope in that message, but there was still this big chasm to cross between how do we get there because nobody is doing it here [NRCS 2015: 5:20min].
Jonathan and his wife Kaylyn crossed that chasm by downsizing the farm to 450 acres, getting rid of the tillage equipment, learning about Holistic Management grazing, and replacing row crops with cover crops and multi-species grazing systems, including beef, lamb, pork and poultry. Now their focus is on building soil health, and they have lush pastures to show for it. Texas still gets hot, but cover crops cool down the soil:
I’ve measured [the hot days] since I’ve gotten into soil health, and on a 103-degree day of ambient temperature, the surface of a bare Blackland soil gets to 155 degrees. You could cook a steak to a safe level. Obviously your soil bacteria are not going to be living at that stage, not the ones you want anyway. Where we had cover residue from no-till and cover crops, my soil surface was 77 degrees on the same day less than a mile away. It’s a drastic change in the environment that you’re creating out there [Acres USA 2015].
On a 103-degree day of ambient temperature, the surface of a bare Blackland soil gets to 155 degrees. You could cook a steak to a safe level. Obviously your soil bacteria are not going to be living at that stage, not the ones you want anyway. Where we had cover residue from no-till and cover crops, my soil surface was 77 degrees on the same day less than a mile away. – Johnathan Cobb [Acres USA 2015]
They direct-market their products online and deliver it to designated pick-up locations in the area. And they enjoy what they do: “If we can make a living and stay here then we couldn’t ask for anything more,” Jonathan says [Voth 2018].
An old-school Italian festival celebrating the work of grazing animals and their faithful herders has taken root in Petaluma, CA. “Transhumance” is the act of moving grazing animals from one grassy site to another. The festival bearing this name takes place in the city or town centers through which the animals traverse en route to fresh paddocks. People gather there to celebrate the cultural tradition of livestock grazing, to trade, and to make merry.
A Petaluma transhumance festival was begun for practical purposes. Sweetgrass Grazing, a local contract grazing business whose client list has expanded in the wake of the recent California wildfires, needed a practical way to move livestock from one client’s site to another. A well-recognized approach for controlling the severity of wildfires is fuel reduction by means of removing vegetation. Yet, in contrast to mechanical or herbicide removal, grazing livestock herds remove vegetation in a way that builds the soil and creates conditions for healthier compositions of grassland species. Furthermore, nimble sheep, goats and herders can also access higher elevations that are inaccessible to machines.
In addition to conditioning townspeople to the idea of herding livestock through town, the festival seeks to sensitize the community at large to the wellbeing of surrounding landscape and possibilities for taking care of that land.
China’s Loess Plateau, roughly the size of France, lies between Tibet and Beijing just south of Mongolia, and is traversed by the Yellow River. Once covered in forest and grassland and the center of Chinese power and wealth, this area eventually became severely degraded by agriculture and unmanaged grazing. The fragile loess soils, composed of glacially deposited fine sediments, were prone to serious erosion when denuded of vegetation. By the 20th Century, the Loess Plateau’s barren landscape was regularly ravaged by dust storms and cycles of flooding, drought and famine. When rain fell, it left the land as quickly as it had come. Some 95% of rainfall simply washed off into gullies, flooding the river and choking it with sediment from extreme erosion.
In the mid-1990s, the Loess Watershed Rehabilitation Project began. The Chinese government working with the World Bank assembled a team of hydrologists, agronomists, and soil and forest specialists to evaluate the problem and what it would take to regreen the region. Apparently engaging local people every step of the way, they identified ecologically destabilizing land management practices, established land management policies (banning agriculture on steep slopes, tree cutting and unmanaged grazing), and developed implementation strategies. Each village was asked to work together to determine how land would be divided fairly among households, each of which received a long-term land use contract for a particular parcel for which they were responsible. Local people were hired to implement ecosystem restoration measures, such as terracing, building small earthen dams to capture rainfall, and planting vegetation. The results have been positive overall, with vegetation and biodiversity returning to a previously desert-like landscape.
More than 224,000 ha of drylands in the Tigray region of northern Ethiopia that had previously succumbed to devastating cycles of drought and flood have been restored. As a result, the hillsides are green again, previously dry wells are recharged, and fruit trees now grow in the valleys. To remedy the problem of severe land degradation, locals throughout the region started in the 1990s to dig small pits and built terraces and bunds (small walls) to capture rainfall and keep it from running off slopes, while also planting millions of tree and bush seedlings. In addition, tree cutting and livestock grazing[6] were banned from degraded lands to allow natural regeneration of vegetation.
“You must have had a lot more rain because how else can water appear where it has not existed before?” asked Zimbabwe Minister of Water Development Sam Nkomo when he saw a clear water-lily-covered pool that had only come to exist in the upper river catchment two years prior [Savory 2009].
Two herders and their employer Allan Savory explained that “the water had come and stayed through the dry season higher up in the river system than it had ever been known [to] before” [Savory 2009]. But this was not due to more rain than usual. Rather, it was because the ranch had increased its cattle and goat numbers 400% and kept them in one herd, which they constantly moved to fresh grazing land according to the needs of the grasses and plants. Managing grazing this way meant the vegetation got quick, strong periodic treatments of trampling, urine and manure, following which it had sufficient time to recover and regrow. This stimulated thicker vegetation cover and better water absorption into the soil, thus increasing groundwater and streamflow.
Faced with drying wells and silt-filled dams nationwide, Minister Nkomo responded positively to his discovery of Savory’s “holistic planned grazing” for restoring rivers and biodiversity and combating drought. In 2009 when this article was published, plans were underway to replicate this grazing management approach in other Zimbabwe watersheds.
Dimbangombe: Success in Africa, stories and photos by Allan Savory:
The Atlantic coast of Canada has started seeing damages related to sea-level rise and storm surges, including flooding, landslides, and shoreline recession. Some communities fear dikes will fail. As a result, people are looking to restoration of native coastal ecosystems as a defense against rising waters. When flooded, coastal marshes often receive large sediment loads that raise their elevation, potentially keeping pace with higher ocean levels. A recent study [Schuerch 2018] showed that making space for inland marsh migration can allow marshes even to thrive in the face of sea-level rise, which is expected to be 1 to 2 feet by 2100 on much of the Canadian Atlantic coast.
In 2010, the Canadian government funded a project in the Bay of Fundy on the Atlantic Coast to restore 16 hectares of saltmarsh land, long ago dried for agriculture, back to its original marshy state. Ducks Unlimited Canada, who led the project, initiated the restoration process by breaching a 150 year-old dike to allow the tide to flow back onto the land. The purpose of this project was to preserve agricultural land further inland from the eroding coastline and rising seas, so a new dike was built just behind the restored marsh.[5]
As noted, marshes can adapt to sea level rise by accumulating sediment. These sediment deposits also enable marshes to quickly bury (or sequester) large amounts of carbon. During the six years after the restoration process began, annual carbon accumulation at the site averaged 13.29 Mg/ha [~5.5 t/ac], mainly due to the sediment deposit that would otherwise likely have mineralized and been released as CO2.
If not deposited in marshes, the organic C in the suspended sediments in the upper Bay of Fundy is likely to be deposited in nearby mudflats. Unlike salt marshes, which tend to be stable or accreting, mudflats are highly dynamic systems subject to frequent erosion events, with scouring to depths of 20 cm or more. … Sediment and associated organic C is also more likely to be preserved in marshes compared with mudflats due to the stabilization effect of macrophyte roots and the associated erosion protection [Wollenburg 2018: 10].
In addition to quickly burying large amounts of carbon, the restored marsh showed signs of success when vegetation (cordgrass) re-established itself in 2012. By 2016, although patches of bare mud were still present, cordgrass was covering most of the marsh area.
This success story is likely to be repeated several times over since the Canadian government announced $75 million for coastal restoration as part of a $1.5 billion Coastal Protection Plan. This includes a project to restore another 75 hectares of salt marshes in the same Bay of Fundy. The coastal restoration fund prioritizes coastal watersheds, estuaries, saltgrass marshes, eel-bed marshes, and migratory corridors for salmon and other species.
The arid San Joaquin Valley of California is intensively farmed and dependent on irrigation. The San Joaquin River, once teeming with migrating fish and other wildlife, is surrounded by farmland and has become warm, muddy, and nearly devoid of aquatic life. In 2012 and 2014, River Partners, a California non-profit dedicated to restoring riparian habitat and river connectivity, partnered with state and federal agencies to buy 2,100 acres of farmland adjacent to the river. The farmers were ready to let the land go because of its proneness to flooding.
To reconnect and restore the floodplain, Dos Rios project workers are breaching berms and levees and planting native trees and shrubs tolerant of ephemeral flooding. This is one of several similar projects managed by River Partners, as well as others led by the state. Further funding for such floodplain restoration efforts is likely to continue given that California voters have passed Proposition 68 or “California Drought, Water, Parks, Climate, Coastal Protection, and Outdoor Access For All Act of 2018.” This legislation to invest $4 billion in park and ecosystem conservation and restoration and climate resiliency includes $300 million for floodplain projects in the Central Valley.
As the hydrological benefits that beaver dams bring to streams and surrounding landscapes becomes better known, ranchers, wildlife managers and researchers are increasingly working together to repair streams by building Beaver Dam Analogs (BDAs). This method is attractive to ranchers searching for ways to manage drought and to irrigate their pastures reliably. In the spring, snowmelt or heavy rainfall can happen quickly, leaving parched landscapes where it’s needed as it rushes downstream into a river and out of sight. Beaver dams slow water down. The human-made BDAs create pools and rehydrate the landscape, ultimately attracting beavers to return, recolonize the streams, and keep the dams in good repair.
“The longer that we can keep that [water] on the landscape, we increase the productivity of those plants. And [that] ultimately leads to more drought resilience, right? These sponges fill up with water. It’s like putting money in the piggy bank for those lean times,” said Jeremy Maestas, an ecologist with the department of agriculture’s Natural Resources Conservation Service. As part of the Sage Grouse Initiative to repair sagebrush habitat, NRCS hosts training workshops throughout the region from Oregon to Montana to Utah on how build the small, porous, beaver-inspired dams in streams.
To reduce the severity of flooding in Lydbrook, Gloucestershire, England, where a 2012 flood did extensive damage, the UK Ministry of Environment released a family of beavers upstream of the village in a 6.5 ha enclosure in a publicly-owned forest. Scientists who have studied the stream believe the beaver dams could hold back some 6,000 cubic meters of water, which might otherwise gush into the village during a heavy storm. If successful, this landfill-tax-funded project is intended to be replicated elsewhere in the UK. Beavers have already been successfully reintroduced elsewhere in England and Scotland, resulting in the Scottish government listing beavers as a protected native species.
Beavers released in Forest of Dean to prevent flooding:
From California to Minnesota, Pennsylvania, Maryland, and Washington DC, people are coming together in their communities to learn what river their watershed drains into, how urban stormwater management has impaired that river, and how to restore river-floodplain ecosystems through a grassroots approach.
A watershed is an area of land over which any rain that falls drains into the same river or water body. For example, all waters falling onto the eastern half of Washington DC flows into the notoriously polluted Anacostia River, while the western half of the nation’s capital drains into the Potomac River. Thus, the city is split into two watersheds.
The Anacostia River was once surrounded by forests, meadows and wetlands, which absorbed, filtered and slowed water on its way downhill to the river. Over time, urban development and industrial processes paved over these natural sponges. The area of tidal wetlands surrounding the Anacostia has shrunk from 2,500 acres in the 1800s to 150 acres today.
Today’s stormwater catchment made up of asphalt streets, parking lots and rooftops leaves water nowhere to go but into storm gutters, gushing out to the river, sometimes flooding over its banks. Furthermore, in many of the older parts of Washington, DC, the infrastructure uses CSOs (“combined sewer overflow”) — where storm drains share pipes with the sewer system — and therefore the stormwater exacerbates water treatment issues. There is thus an added incentive to reduce stormwater runoff.
The Anacostia Watershed Society (AWS), a D.C. non-profit with a mission to make the river “fishable and swimmable by 2025,” engages school children and other community members in wetland restoration along the river. In addition, as in several other communities around the country, AWS partners with the DC Dept of Environment to train community members to be ambassadors for the river. Over the course of a several-week training program, Watershed Stewards learn how individual houses and buildings contribute to the problem with impervious surfaces and gutter downspouts directing rain water directly into storm sewers. Then they learn about absorptive green rooftops, and the possibility of redirecting water from a downspout to a rain garden or a deep-rooted perennial bed, where the water can percolate into healthy spongy soil, ultimately recharging groundwater.
Primed with knowledge, enthusiasm, and the camaraderie of fellow stewards, participants are expected to implement a project of their own, to teach their neighbors what they’ve learned, and to volunteer in related community projects focusing on watershed restoration. In Minnesota, watershed steward projects redirect rainfall from gutters into gardens, where it can hydrate plants and recharge groundwater, at a rate of more than 1 million gallons per year. According to the program website, this outcome is due to the efforts initiated in 2013 which now include 141 stewards working in partnership with seven watershed districts and one municipality. An Anne Arundel, Maryland, program started in 2009 boasts having planted nearly 100,000 native plants, trees and shrubs, led by some 200 stewards in 100 communities engaging 134,000 of their neighbors in watershed restoration efforts.
“In the past, humans have taken the land away from the water; now we need to give the land back.” – Professor Hui Li [Guardian 2017]
Faced with severe flooding in many cities across China, such as a major 2012 Beijing flood, the Chinese government announced the Sponge Cities Initiative in 2014 as a remedy. The national government identified 16 cities as pilot sites, soon adding another 14 cities, including Beijing. The sponge city concept represents a paradigm shift in flood management away from impervious surfaces and chutes meant to swiftly drain a city after a storm. By contrast, a sponge city aims to manage stormwaters by vastly increasing the amount of soft, permeable surfaces that can absorb water where it falls, filter it, and store it in vegetation, ponds and aquifers.
A sponge city aims to manage stormwaters by vastly increasing the amount of soft, permeable surfaces that can absorb water where it falls, filter it, and store it in vegetation, ponds and aquifers.
According to the “Guideline of Sponge City Construction”, the target of the approach is to increase the area of urban land able to absorb surface water discharges by approximately 20%, and to retain or reuse approximately 70% of urban stormwater by 2020; and further reuse up to 80% of stormwater by 2030s. This means that the ideology of the Sponge City concept is not only addressing urban flood risk, but also taking a proactive approach to collection, purification and reuse of urban stormwater in Chinese cities to address future climatic extremes (floods and droughts) [Chan 2018: 3-4].
One of China’s new sponge cities, the Lingang/Nanhui district of Shanghai, is buildIng streets with permeable pavement, which allows water to percolate into the ground beneath, and planting rain gardens between lanes of traffic as well as some 400,000 square meters of rooftop gardens. In another city, the 84-acre Qunli Stormwater Park consists of a wetland surrounded by newly constructed filtering ponds that collect and filter the city’s stormwater runoff before slowly releasing it into the wetland, which in turn recharges groundwater. Earthen mounds are planted in cottonwood trees, and a system of elevated pathways through the park allows people to enjoy the space.
Inspiration for Qunli Stormwater Park and several other sponge city landscape design features comes from the ancient water management practices of Chinese peasants. According to landscape architect and sponge city advocate Yu Kongjian, peasants constructed simple terraces in combination with ponds to regulate flood and drought. “On sloping ground in monsoon regions these water catchments are critical,” Yu explains in an essay explaining the sponge city concept [Yu 2017: 29]. “Peasants also employed crop rotation to maximize yield, beautifully sustaining humanity for thousands of years” [Yu 2017: 29]. He continues :
Ironically, these centuries-old productive landscapes have given way to urbanization. Fine terraces are leveled into … planes called developable land; small ponds are drained and replaced with underground drainage systems; ponds and dikes give way to mechanical farming. The centuries-old ecosystem balance is broken, leading to flood, drought and habitat loss. Grey infrastructure haunts Chinese cities, while high maintenance landscapes with ornamental planting make broad scale landscape change unaffordable. [Yu 2017: 29]
We know how to enhance resilience to extreme weather where we live and work. Communities throughout the world are utilizing these approaches, and here we highlight several initiatives in a variety of habitats to illustrate potential paths forward. More information is included just below each project description. Following this section is a collection of summaries of scientific articles that provide evidence for eco-restoration to enhance resilience to a chaotic climate.
Sponge cities, China
“In the past, humans have taken the land away from the water; now we need to give the land back.” – Professor Hui Li [Guardian 2017]
Faced with severe flooding in many cities across China, such as a major 2012 Beijing flood, the Chinese government announced the Sponge Cities Initiative in 2014 as a remedy. The national government identified 16 cities as pilot sites, soon adding another 14 cities, including Beijing. The sponge city concept represents a paradigm shift in flood management away from impervious surfaces and chutes meant to swiftly drain a city after a storm. By contrast, a sponge city aims to manage stormwaters by vastly increasing the amount of soft, permeable surfaces that can absorb water where it falls, filter it, and store it in vegetation, ponds and aquifers.
A sponge city aims to manage stormwaters by vastly increasing the amount of soft, permeable surfaces that can absorb water where it falls, filter it, and store it in vegetation, ponds and aquifers.
According to the “Guideline of Sponge City Construction”, the target of the approach is to increase the area of urban land able to absorb surface water discharges by approximately 20%, and to retain or reuse approximately 70% of urban stormwater by 2020; and further reuse up to 80% of stormwater by 2030s. This means that the ideology of the Sponge City concept is not only addressing urban flood risk, but also taking a proactive approach to collection, purification and reuse of urban stormwater in Chinese cities to address future climatic extremes (floods and droughts) [Chan 2018: 3-4].
One of China’s new sponge cities, the Lingang/Nanhui district of Shanghai, is buildIng streets with permeable pavement, which allows water to percolate into the ground beneath, and planting rain gardens between lanes of traffic as well as some 400,000 square meters of rooftop gardens. In another city, the 84-acre Qunli Stormwater Park consists of a wetland surrounded by newly constructed filtering ponds that collect and filter the city’s stormwater runoff before slowly releasing it into the wetland, which in turn recharges groundwater. Earthen mounds are planted in cottonwood trees, and a system of elevated pathways through the park allows people to enjoy the space.
Inspiration for Qunli Stormwater Park and several other sponge city landscape design features comes from the ancient water management practices of Chinese peasants. According to landscape architect and sponge city advocate Yu Kongjian, peasants constructed simple terraces in combination with ponds to regulate flood and drought. “On sloping ground in monsoon regions these water catchments are critical,” Yu explains in an essay explaining the sponge city concept [Yu 2017: 29]. “Peasants also employed crop rotation to maximize yield, beautifully sustaining humanity for thousands of years” [Yu 2017: 29]. He continues :
Ironically, these centuries-old productive landscapes have given way to urbanization. Fine terraces are leveled into … planes called developable land; small ponds are drained and replaced with underground drainage systems; ponds and dikes give way to mechanical farming. The centuries-old ecosystem balance is broken, leading to flood, drought and habitat loss. Grey infrastructure haunts Chinese cities, while high maintenance landscapes with ornamental planting make broad scale landscape change unaffordable. [Yu 2017: 29]
Community-based watershed stewardship programs, USA
From California to Minnesota, Pennsylvania, Maryland, and Washington DC, people are coming together in their communities to learn what river their watershed drains into, how urban stormwater management has impaired that river, and how to restore river-floodplain ecosystems through a grassroots approach.
A watershed is an area of land over which any rain that falls drains into the same river or water body. For example, all waters falling onto the eastern half of Washington DC flows into the notoriously polluted Anacostia River, while the western half of the nation’s capital drains into the Potomac River. Thus, the city is split into two watersheds.
The Anacostia River was once surrounded by forests, meadows and wetlands, which absorbed, filtered and slowed water on its way downhill to the river. Over time, urban development and industrial processes paved over these natural sponges. The area of tidal wetlands surrounding the Anacostia has shrunk from 2,500 acres in the 1800s to 150 acres today.
Today’s stormwater catchment made up of asphalt streets, parking lots and rooftops leaves water nowhere to go but into storm gutters, gushing out to the river, sometimes flooding over its banks. Furthermore, in many of the older parts of Washington, DC, the infrastructure uses CSOs (“combined sewer overflow”) — where storm drains share pipes with the sewer system — and therefore the stormwater exacerbates water treatment issues. There is thus an added incentive to reduce stormwater runoff.
The Anacostia Watershed Society (AWS), a D.C. non-profit with a mission to make the river “fishable and swimmable by 2025,” engages school children and other community members in wetland restoration along the river. In addition, as in several other communities around the country, AWS partners with the DC Dept of Environment to train community members to be ambassadors for the river. Over the course of a several-week training program, Watershed Stewards learn how individual houses and buildings contribute to the problem with impervious surfaces and gutter downspouts directing rain water directly into storm sewers. Then they learn about absorptive green rooftops, and the possibility of redirecting water from a downspout to a rain garden or a deep-rooted perennial bed, where the water can percolate into healthy spongy soil, ultimately recharging groundwater.
Primed with knowledge, enthusiasm, and the camaraderie of fellow stewards, participants are expected to implement a project of their own, to teach their neighbors what they’ve learned, and to volunteer in related community projects focusing on watershed restoration. In Minnesota, watershed steward projects redirect rainfall from gutters into gardens, where it can hydrate plants and recharge groundwater, at a rate of more than 1 million gallons per year. According to the program website, this outcome is due to the efforts initiated in 2013 which now include 141 stewards working in partnership with seven watershed districts and one municipality. An Anne Arundel, Maryland, program started in 2009 boasts having planted nearly 100,000 native plants, trees and shrubs, led by some 200 stewards in 100 communities engaging 134,000 of their neighbors in watershed restoration efforts.
To reduce the severity of flooding in Lydbrook, Gloucestershire, England, where a 2012 flood did extensive damage, the UK Ministry of Environment released a family of beavers upstream of the village in a 6.5 ha enclosure in a publicly-owned forest. Scientists who have studied the stream believe the beaver dams could hold back some 6,000 cubic meters of water, which might otherwise gush into the village during a heavy storm. If successful, this landfill-tax-funded project is intended to be replicated elsewhere in the UK. Beavers have already been successfully reintroduced elsewhere in England and Scotland, resulting in the Scottish government listing beavers as a protected native species.
Beavers released in Forest of Dean to prevent flooding:
Low-tech stream repair for drought resilience: western USA
As the hydrological benefits that beaver dams bring to streams and surrounding landscapes becomes better known, ranchers, wildlife managers and researchers are increasingly working together to repair streams by building Beaver Dam Analogs (BDAs). This method is attractive to ranchers searching for ways to manage drought and to irrigate their pastures reliably. In the spring, snowmelt or heavy rainfall can happen quickly, leaving parched landscapes where it’s needed as it rushes downstream into a river and out of sight. Beaver dams slow water down. The human-made BDAs create pools and rehydrate the landscape, ultimately attracting beavers to return, recolonize the streams, and keep the dams in good repair.
“The longer that we can keep that [water] on the landscape, we increase the productivity of those plants. And [that] ultimately leads to more drought resilience, right? These sponges fill up with water. It’s like putting money in the piggy bank for those lean times,” said Jeremy Maestas, an ecologist with the department of agriculture’s Natural Resources Conservation Service. As part of the Sage Grouse Initiative to repair sagebrush habitat, NRCS hosts training workshops throughout the region from Oregon to Montana to Utah on how build the small, porous, beaver-inspired dams in streams.
The arid San Joaquin Valley of California is intensively farmed and dependent on irrigation. The San Joaquin River, once teeming with migrating fish and other wildlife, is surrounded by farmland and has become warm, muddy, and nearly devoid of aquatic life. In 2012 and 2014, River Partners, a California non-profit dedicated to restoring riparian habitat and river connectivity, partnered with state and federal agencies to buy 2,100 acres of farmland adjacent to the river. The farmers were ready to let the land go because of its proneness to flooding.
To reconnect and restore the floodplain, Dos Rios project workers are breaching berms and levees and planting native trees and shrubs tolerant of ephemeral flooding. This is one of several similar projects managed by River Partners, as well as others led by the state. Further funding for such floodplain restoration efforts is likely to continue given that California voters have passed Proposition 68 or “California Drought, Water, Parks, Climate, Coastal Protection, and Outdoor Access For All Act of 2018.” This legislation to invest $4 billion in park and ecosystem conservation and restoration and climate resiliency includes $300 million for floodplain projects in the Central Valley.
The Atlantic coast of Canada has started seeing damages related to sea-level rise and storm surges, including flooding, landslides, and shoreline recession. Some communities fear dikes will fail. As a result, people are looking to restoration of native coastal ecosystems as a defense against rising waters. When flooded, coastal marshes often receive large sediment loads that raise their elevation, potentially keeping pace with higher ocean levels. A recent study [Schuerch 2018] showed that making space for inland marsh migration can allow marshes even to thrive in the face of sea-level rise, which is expected to be 1 to 2 feet by 2100 on much of the Canadian Atlantic coast.
In 2010, the Canadian government funded a project in the Bay of Fundy on the Atlantic Coast to restore 16 hectares of saltmarsh land, long ago dried for agriculture, back to its original marshy state. Ducks Unlimited Canada, who led the project, initiated the restoration process by breaching a 150 year-old dike to allow the tide to flow back onto the land. The purpose of this project was to preserve agricultural land further inland from the eroding coastline and rising seas, so a new dike was built just behind the restored marsh.[5]
As noted, marshes can adapt to sea level rise by accumulating sediment. These sediment deposits also enable marshes to quickly bury (or sequester) large amounts of carbon. During the six years after the restoration process began, annual carbon accumulation at the site averaged 13.29 Mg/ha [~5.5 t/ac], mainly due to the sediment deposit that would otherwise likely have mineralized and been released as CO2.
If not deposited in marshes, the organic C in the suspended sediments in the upper Bay of Fundy is likely to be deposited in nearby mudflats. Unlike salt marshes, which tend to be stable or accreting, mudflats are highly dynamic systems subject to frequent erosion events, with scouring to depths of 20 cm or more. … Sediment and associated organic C is also more likely to be preserved in marshes compared with mudflats due to the stabilization effect of macrophyte roots and the associated erosion protection [Wollenburg 2018: 10].
In addition to quickly burying large amounts of carbon, the restored marsh showed signs of success when vegetation (cordgrass) re-established itself in 2012. By 2016, although patches of bare mud were still present, cordgrass was covering most of the marsh area.
This success story is likely to be repeated several times over since the Canadian government announced $75 million for coastal restoration as part of a $1.5 billion Coastal Protection Plan. This includes a project to restore another 75 hectares of salt marshes in the same Bay of Fundy. The coastal restoration fund prioritizes coastal watersheds, estuaries, saltgrass marshes, eel-bed marshes, and migratory corridors for salmon and other species.
Holistic planned grazing for drought relief, Zimbabwe
“You must have had a lot more rain because how else can water appear where it has not existed before?” asked Zimbabwe Minister of Water Development Sam Nkomo when he saw a clear water-lily-covered pool that had only come to exist in the upper river catchment two years prior [Savory 2009].
Two herders and their employer Allan Savory explained that “the water had come and stayed through the dry season higher up in the river system than it had ever been known [to] before” [Savory 2009]. But this was not due to more rain than usual. Rather, it was because the ranch had increased its cattle and goat numbers 400% and kept them in one herd, which they constantly moved to fresh grazing land according to the needs of the grasses and plants. Managing grazing this way meant the vegetation got quick, strong periodic treatments of trampling, urine and manure, following which it had sufficient time to recover and regrow. This stimulated thicker vegetation cover and better water absorption into the soil, thus increasing groundwater and streamflow.
Faced with drying wells and silt-filled dams nationwide, Minister Nkomo responded positively to his discovery of Savory’s “holistic planned grazing” for restoring rivers and biodiversity and combating drought. In 2009 when this article was published, plans were underway to replicate this grazing management approach in other Zimbabwe watersheds.
Dimbangombe: Success in Africa, stories and photos by Allan Savory:
More than 224,000 ha of drylands in the Tigray region of northern Ethiopia that had previously succumbed to devastating cycles of drought and flood have been restored. As a result, the hillsides are green again, previously dry wells are recharged, and fruit trees now grow in the valleys. To remedy the problem of severe land degradation, locals throughout the region started in the 1990s to dig small pits and built terraces and bunds (small walls) to capture rainfall and keep it from running off slopes, while also planting millions of tree and bush seedlings. In addition, tree cutting and livestock grazing[6] were banned from degraded lands to allow natural regeneration of vegetation.
China’s Loess Plateau, roughly the size of France, lies between Tibet and Beijing just south of Mongolia, and is traversed by the Yellow River. Once covered in forest and grassland and the center of Chinese power and wealth, this area eventually became severely degraded by agriculture and unmanaged grazing. The fragile loess soils, composed of glacially deposited fine sediments, were prone to serious erosion when denuded of vegetation. By the 20th Century, the Loess Plateau’s barren landscape was regularly ravaged by dust storms and cycles of flooding, drought and famine. When rain fell, it left the land as quickly as it had come. Some 95% of rainfall simply washed off into gullies, flooding the river and choking it with sediment from extreme erosion.
In the mid-1990s, the Loess Watershed Rehabilitation Project began. The Chinese government working with the World Bank assembled a team of hydrologists, agronomists, and soil and forest specialists to evaluate the problem and what it would take to regreen the region. Apparently engaging local people every step of the way, they identified ecologically destabilizing land management practices, established land management policies (banning agriculture on steep slopes, tree cutting and unmanaged grazing), and developed implementation strategies. Each village was asked to work together to determine how land would be divided fairly among households, each of which received a long-term land use contract for a particular parcel for which they were responsible. Local people were hired to implement ecosystem restoration measures, such as terracing, building small earthen dams to capture rainfall, and planting vegetation. The results have been positive overall, with vegetation and biodiversity returning to a previously desert-like landscape.
Culture revival of livestock grazing for wildfire management, California
An old-school Italian festival celebrating the work of grazing animals and their faithful herders has taken root in Petaluma, CA. “Transhumance” is the act of moving grazing animals from one grassy site to another. The festival bearing this name takes place in the city or town centers through which the animals traverse en route to fresh paddocks. People gather there to celebrate the cultural tradition of livestock grazing, to trade, and to make merry.
A Petaluma transhumance festival was begun for practical purposes. Sweetgrass Grazing, a local contract grazing business whose client list has expanded in the wake of the recent California wildfires, needed a practical way to move livestock from one client’s site to another. A well-recognized approach for controlling the severity of wildfires is fuel reduction by means of removing vegetation. Yet, in contrast to mechanical or herbicide removal, grazing livestock herds remove vegetation in a way that builds the soil and creates conditions for healthier compositions of grassland species. Furthermore, nimble sheep, goats and herders can also access higher elevations that are inaccessible to machines.
In addition to conditioning townspeople to the idea of herding livestock through town, the festival seeks to sensitize the community at large to the wellbeing of surrounding landscape and possibilities for taking care of that land.
Diverse cover crops and livestock for drought relief, Texas
The 2011 drought in Texas was the worst in recorded history and it lasted until 2015. The ground was so dry that Jonathan Cobb, a 4th generation farmer in Blackland Prairie of central Texas, couldn’t even get crops planted. His 2,500-acre conventional row crop operation was already struggling financially through a treadmill of increasingly more inputs and long hours with little or no yield improvement. So he made the hard decision to leave farming and move to the city.
But before he left, he attended a workshop with renowned soil scientist Ray Archuleta, who focused Jonathan’s attention for the first time on soil health. Archuleta demonstrated the water-holding capacity of healthy, biologically active aggregated soils compared to a typical compacted soil that crumbled and eroded when water was poured on it. This demonstration opened Cobb’s mind to a whole new way of approaching agriculture. It gave him hope that farming even through extreme drought was possible.
There was hope that nature actually did exist on its own before man started cultivating it … things that seem obvious, like a forest ecology that nobody fertilizes a forest … of course it works, it can be very abundant. And so there was a hope in that message, but there was still this big chasm to cross between how do we get there because nobody is doing it here [NRCS 2015: 5:20min].
Jonathan and his wife Kaylyn crossed that chasm by downsizing the farm to 450 acres, getting rid of the tillage equipment, learning about Holistic Management grazing, and replacing row crops with cover crops and multi-species grazing systems, including beef, lamb, pork and poultry. Now their focus is on building soil health, and they have lush pastures to show for it. Texas still gets hot, but cover crops cool down the soil:
I’ve measured [the hot days] since I’ve gotten into soil health, and on a 103-degree day of ambient temperature, the surface of a bare Blackland soil gets to 155 degrees. You could cook a steak to a safe level. Obviously your soil bacteria are not going to be living at that stage, not the ones you want anyway. Where we had cover residue from no-till and cover crops, my soil surface was 77 degrees on the same day less than a mile away. It’s a drastic change in the environment that you’re creating out there [Acres USA 2015].
On a 103-degree day of ambient temperature, the surface of a bare Blackland soil gets to 155 degrees. You could cook a steak to a safe level. Obviously your soil bacteria are not going to be living at that stage, not the ones you want anyway. Where we had cover residue from no-till and cover crops, my soil surface was 77 degrees on the same day less than a mile away. – Johnathan Cobb [Acres USA 2015]
They direct-market their products online and deliver it to designated pick-up locations in the area. And they enjoy what they do: “If we can make a living and stay here then we couldn’t ask for anything more,” Jonathan says [Voth 2018].
Given competing interests for floodplain property, some have argued for strategic partial reconnection of floodplains to the river by allowing portions of floodplain to flood, so that pressure elsewhere along the river during a flood may be alleviated [Opperman 2009].
For example, California’s Yolo Bypass was created in the early 1900s after the Sacramento River flooded several times and levees proved inadequate to protect the city [Sommer 2001]. The site of the bypass was historically the vast wetland floodplain of the Sacramento River and other nearby rivers and streams that had since been converted to agriculture. Today, the bypass reconnects the river to its floodplains.
Since1997, Yolo Bypass also features more than 16,000 acres of wildlife area, including seasonal and perennial wetlands, riparian forest, pasture, and seasonal crop production where fields are allowed to flood during winter. This solution has not only kept Sacramento dry during numerous high water events, it has also restored diverse populations of fish, bird, snake, mammal and other species to the area, while providing recreational and educational opportunities to nearby communities.
While the Yolo Bypass Wilderness Area exemplifies the large-scale engineered reconnection of a major river to its floodplain, many smaller stream floodplains benefit from the work of non-human engineers. Long considered the nemesis of ranchers and farmers alike, beavers caught damming irrigation ditches or flooding fields are often summarily trapped and killed. Yet in Elko, Nevada, beavers and altered livestock grazing regimes have brought stream beds back to life [Goldfarb 2018, Evans & Griggs 2015].
One rancher’s management change began by excluding cattle from grazing along Elko’s Suzie Creek – just during the hot season when plants are vulnerable. This allowed rushes, sedges and other vegetation to grow back, slowing water down enough for sediment to fill in the gouged out gully and raise the streambed back up to the level of its floodplain. Once beavers discovered their favorite food – willow – growing again at Suzie Creek, they moved in and have since built 139 dams there. These dams, in turn, raised the water table by about two feet, becoming the natural irrigation system for the ranch’s now lush riparian pastureland. The beaver dams proved vital in 2012-2015 when Suzie Creek kept flowing despite several summers of drought that left the rest of the region parched.
The beaver dams proved vital in 2012-2015 when Suzie Creek kept flowing despite several summers of drought that left the rest of the region parched.
At around the same time as Suzie Creek’s revitalization, across the Atlantic in Devon, England, a pair of Eurasian beavers were introduced to a wooded stream at the headwaters of the Tamar River [Puttock 2016]. A few years and 13 dams later, the beavers’ activity was filtering pollutants out of water passing through the dam sites and slowing the flow so as to minimize downstream flooding during storms. As in North America, beavers were once abundant in Europe, but by the 16th Century were wiped out in the UK. In recognition of beavers’ beneficial effects on hydrological systems, multiple reintroduction programs have begun establishing colonies across Europe and North America.
“Because of their abilities to modify streams and floodplains, beavers have the potential to play a critical role in shaping how riparian and stream ecosystems respond to climate change,” explain the authors of a recent study of potentially suitable beaver reintroduction sites [Dittbrenner 2018: 2]. The authors continue:
By damming streams, beavers create pond and wetland complexes that increase … species and habitat diversity, and therefore ecosystem resilience to climate-induced environmental change. Beaver impoundments slow stream velocity allowing sediment suspended in the water column to settle, aggrading incised stream systems, and reconnecting streams with their floodplains. The increase in surface water promotes groundwater recharge, storage, and supplementation during base flows. The increased geomorphic complexity also promotes higher thermal variability and cold-water refugia in deeper waters and in areas of downstream upwelling [Dittbrenner 2018: 2].
Furthermore, by repairing hydrological functioning and increasing a landscape’s overall level of moisture, beaver populations could literally dampen conditions for wildfire, which is intensified by drought [Maughan 2013].
Such vital ecosystem services provided by a keystone species like beaver in the era of climate change are nothing to scoff at. And while beavers are still trapped and killed for sport and by hunters or land managers whoconsider them a nuisance [Goldfarb 2018], castor canadensis is also increasingly accepted, as evidenced by more pro-beaver attitudes within the fish and wildlife departments of western states that had previously considered them mainly as pests. Beavers are discussed on state wildlife agency websites in terms of “living with wildlife,”[4] where beaver life histories are described, along with explanations of the benefits of beaver dams for landowners and for the landscape overall.
Similarly, Holland is learning new ways to live with water. In the Netherlands, literally “low country” due to much of its land area being at or below sea level, there is an age-old struggle with water and flooding, notably through the use of dikes. However, alarmed by recent flooding and the prospects of sea-level rise from climate change, the nation is undergoing a paradigm shift wherein the guiding principle for water management has become “make room for the river” [Pahl-Wostl 2006]. Among other tactics, certain areas are being “depoldered,” meaning dikes removed from low-lying areas and the land returned to wetland; people living in those areas are assisted in relocating [Bentley 2016].
Similarly, several cities in China are striving to “make friends with water” through adoption of the concept of “sponge cities” that aim to “retain, adapt, slow down and reuse” stormwater by increasing the porosity of urban surfaces, including increasing the amount of ecologically functional urban green space [Guardian 2018].
In our panic over increasing numbers of extreme weather events, we may grasp at familiar solutions – extra air-conditioning to shelter from the heat, higher levees to hold back floodwaters, more irrigation to combat drought, or logging forests to reduce wildfire fuel. While these measures may (or may not) temporarily bandage the situation, they increase the fragility of our built environment and usher us further down the path of climate chaos though unrelenting energy consumption and increasingly hobbled ecosystems. For our own sake, it’s time to make friends with nature and to acknowledge her superior power by partnering with instead of continuously fighting her.
The concept of hydrological drought (as distinct from meteorological drought) helps explain the success of these age-old techniques to enhance surface and groundwater supply. Meteorological drought is the occurence of abnormally low rainfall for a given region. Hydrological drought is a consequence of meteorological drought – it happens when surface and ground waters run low thanks to a prolonged rainfall shortage compared to historical conditions for a region. Human water consumption for irrigation, industry and household use intensifies hydrological drought [Wada 2013].
Yet, as seen in the examples above, proper land management can raise the water table in an area in spite of occasional episodes of meteorological drought. In other words, while we cannot directly control when and how much rain falls, we can manage what happens to water once it reaches the ground.
Rain falling on much of our modern built environment is managed with ditches, gutters, drains and sewers designed to whisk it away as quickly as possible, rather than absorbing it in place. Farmland too is sometimes fitted with underground “tiles” (pipes) to drain fields, or with dikes to keep water out. Moreover, by the very absence of design, nearly all conventional farmland is so lacking in organic matter that it can barely absorb rainfall, which instead runs off the soil surface carrying soil with it.
In short, water hitting the ground in today’s world moves quickly. Stormwater moves in torrents over land seeking an outlet. That is until the outlet is full, at which point the water stagnates, rising like a bathtub, soaking and rotting property.
Ironically, the water we seek to drain away when there’s plenty becomes desperately lacking after the rain has stopped. Two sides of the same coin, flooding and drought often go hand in hand. By the same token, because a surfeit of impervious surfaces is at the root of these twin challenges, the solution of turning more land into a spongy surface helps resolve both problems at once. Spongy surfaces slow down water, allowing it to percolate into groundwater reserves.
Two sides of the same coin, flooding and drought often go hand in hand. By the same token, because a surfeit of impervious surfaces is at the root of these twin challenges, the solution of turning more land into a spongy surface helps resolve both problems at once. Spongy surfaces slow down water, allowing it to percolate into groundwater reserves.
Ecosystem restoration creates a spongy land surface by protecting soil with vegetation, thus allowing the soil to repair itself with biodiversity and organic matter, key ingredients of a good soil sponge. And beyond fostering drought and flood resilience, healthy ecosystems serve a myriadof protective functions, including cooling their surroundings, cleaning polluted water, drawing down CO2, and harboring the biodiversity that is the magic making ecosystems perform so many vital functions.
However, because the services rendered by nature go widely unrecognized or taken for granted, nature’s power as an ally is often shackled.
Human societies tend to value the potential benefits that a landscape might provide in a limited way, adjusting management practices towards desired outputs by maximizing the benefits gained from one or some of the services (often the provision of goods) leading to the loss of multifunctionality and the degradation of natural capital at the expense of human welfare [Schindler 2014: 230].
An example of a landscape with undervalued ecosystem function is a river floodplain. Because floodplains are often favored for agricultural, industrial, commercial or residential uses, rivers are constrained to their channels and their banks leveed, despite that a river and its floodplains are members of a single interdependent ecosystem. Through seasonal pulses of floodwater over the banks, like a heart pumping blood through a body, a river replenishes groundwater and nutrients throughout its floodplains, making these areas some of the planet’s most productive and biodiverse ecosystems [Sparks 1995]. Consider, for example, that the Amazon Basin is a floodplain, where fish biodiversity actually depends on riparian forest cover [Arantes 2017]. Fish swim into flooded forests during flood pulses and directly consume terrestrial floodplain vegetation (seeds, fruits, detritus).
…a river and its floodplains are members of a single interdependent ecosystem. Through seasonal pulses of floodwater over the banks, like a heart pumping blood through a body, a river replenishes groundwater and nutrients throughout its floodplains, making these areas some of the planet’s most productive and biodiverse ecosystems.
In addition to supporting robust biodiversity, floodplain ecosystems give water somewhere to go during severe flooding events rather than damaging cropland, houses or other properties. By allowing water time to infiltrate into the ground, floodplains recharge groundwater, thus alleviating future droughts [Opperman 2009]. Riparian ecosystems also serve as a migration corridor for birds and fish especially, and also as a refuge from the heat of more exposed areas.
In other words, intact river-floodplain ecosystems perform multiple ecosystem services and thus help us manage some of our most pressing societal problems if only we acknowledge the value of floodplains in these terms.
Managing rainwater within a landscape so that neither heavy storms nor long dry spells devastate human endeavors and constructions is referred by Yu Kongjian as the “art of survival” [Yu 2012]. This Chinese landscape architect with an ecological mindset learned the art of survival by studying the ways of ancient peasant farmers. He contrasts the wisdom embodied in their simple structures, such as terraced crop fields on sloping land designed to capture and hold storm water for later usewith the modern “art of pleasure making and ornament.”
Modern urban design tends to favor non-functional decorative features – “monumental architecture” like a grand stadium, manicured lawns, or fruit trees grown for their blossoms rather than their fruit, for example. Such investments are pretty, but expensive and “easily superseded,” according to Yu. Equally beautiful urban designs such as green rooftops and boardwalk-accessible urban wetland parks, on the other hand, can be affordable, high-performing features designed to withstand environmental extremes.
World cities, and especially those in China, face deepening environmental problems: flood, drought, pollution, aquifer drop, loss of natural habitat and cultural heritage. A low-culture approach using what I term ‘adaptive design’ provides a technique for solving problems in an economical and ecological way [Yu 2012: 72].
For Yu, design is adaptive “when it responds elegantly and efficiently to its environmental setting so that new uses can endure” [Yu 2012: 72], meaning a design for urban resilience in the face of ever more severe weather.
It’s not only Chinese peasants who understood water cycles and how to manage them. Ancient peoples of the Middle East actually depended on seasonal flooding. Like we do today, these ancient farmers grew crops on river floodplains. Yet unlike today’s practices of diking off the river to keep croplands dry, they allowed annual flood pulses onto their fields.
Annual flood pulses are so predictable and long-lasting that plants, animals, and even human societies have adapted to take advantage of them. In ancient Egypt and Mesopotamia, the fertility of the soils was renewed each year by the annual overflow of the rivers, thereby sustaining large populations in one place for millennia and permitting the development of great civilizations [Sparks 1995: 168].
In India’s Rajasthan region, where the monsoon cycle brings torrents of rain all at once, after which begins a long dry season, rural farming communities built johads. These are small earthen dams on sloping land that create ponds or wetlands by harvesting stormwater during the rainy season. This water reserve then becomes a vital resource during the dry season. However, johads were abandoned in favor of more modern borehole wells, which are deep, not wide like a basin, and therefore not able to catch rainwater, which instead simply ran off the landscape. By the 1980s the johads were gone, wells were dry, people walked 9km in search of drinking water, and farmers left town in search of other employment. But then communities throughout the region started to rebuild johads, which raised the water table enough to refill wells, support agriculture and wildlife, and for streams to flow again [Singh 2015, SIWI 2015].
Similarly, severe drought in Burkina Faso in the Sahel region bordering the Sahara Desert presented farmers with a simple choice: find a way to restore the land and farm again, or migrate. As in Rajasthan, these farmers were prompted in desperation to rediscover their region’s own traditional techniques for water conservation. They dug rows of small pits in their fields to capture rainfall, which they filled with compost and manure, and into which they planted crops. Other farmers built stone terraces along the contours in their fields to capture rainfall and prevent runoff; some farmers did both. Over time, at least 140,000 farming households over 200,000 hectares or more were practicing these techniques, resulting in the revival of crop production, reestablishment of trees, shrubs and grasses, and the recharging of the area’s water table by a depth of five meters [Reij 2009].
While previous issues of the Compendium have addressed ecosystem strategies to reverse global warming, here we discuss ecosystem restoration to adapt to the consequences of climate change. From drought in Cape Town and wildfire in California and Greece to flooding in Beijing, Paris, Houston and North Carolina, each new report of catastrophe makes climate change more real and more frightening. And while taking the giant steps required within the next 12 years to avert climate catastrophe as the IPCC advises[3] may seem out of reach for everyday people, anyone can act locally and regionally to restore the ecosystems that protect our homes and communities. Happily, healthy ecosystems contribute both to mitigation and resilience.
Slowing down water and the art of survival
Managing rainwater within a landscape so that neither heavy storms nor long dry spells devastate human endeavors and constructions is referred by Yu Kongjian as the “art of survival” [Yu 2012]. This Chinese landscape architect with an ecological mindset learned the art of survival by studying the ways of ancient peasant farmers. He contrasts the wisdom embodied in their simple structures, such as terraced crop fields on sloping land designed to capture and hold storm water for later usewith the modern “art of pleasure making and ornament.”
Modern urban design tends to favor non-functional decorative features – “monumental architecture” like a grand stadium, manicured lawns, or fruit trees grown for their blossoms rather than their fruit, for example. Such investments are pretty, but expensive and “easily superseded,” according to Yu. Equally beautiful urban designs such as green rooftops and boardwalk-accessible urban wetland parks, on the other hand, can be affordable, high-performing features designed to withstand environmental extremes.
World cities, and especially those in China, face deepening environmental problems: flood, drought, pollution, aquifer drop, loss of natural habitat and cultural heritage. A low-culture approach using what I term ‘adaptive design’ provides a technique for solving problems in an economical and ecological way [Yu 2012: 72].
For Yu, design is adaptive “when it responds elegantly and efficiently to its environmental setting so that new uses can endure” [Yu 2012: 72], meaning a design for urban resilience in the face of ever more severe weather.
It’s not only Chinese peasants who understood water cycles and how to manage them. Ancient peoples of the Middle East actually depended on seasonal flooding. Like we do today, these ancient farmers grew crops on river floodplains. Yet unlike today’s practices of diking off the river to keep croplands dry, they allowed annual flood pulses onto their fields.
Annual flood pulses are so predictable and long-lasting that plants, animals, and even human societies have adapted to take advantage of them. In ancient Egypt and Mesopotamia, the fertility of the soils was renewed each year by the annual overflow of the rivers, thereby sustaining large populations in one place for millennia and permitting the development of great civilizations [Sparks 1995: 168].
In India’s Rajasthan region, where the monsoon cycle brings torrents of rain all at once, after which begins a long dry season, rural farming communities built johads. These are small earthen dams on sloping land that create ponds or wetlands by harvesting stormwater during the rainy season. This water reserve then becomes a vital resource during the dry season. However, johads were abandoned in favor of more modern borehole wells, which are deep, not wide like a basin, and therefore not able to catch rainwater, which instead simply ran off the landscape. By the 1980s the johads were gone, wells were dry, people walked 9km in search of drinking water, and farmers left town in search of other employment. But then communities throughout the region started to rebuild johads, which raised the water table enough to refill wells, support agriculture and wildlife, and for streams to flow again [Singh 2015, SIWI 2015].
Similarly, severe drought in Burkina Faso in the Sahel region bordering the Sahara Desert presented farmers with a simple choice: find a way to restore the land and farm again, or migrate. As in Rajasthan, these farmers were prompted in desperation to rediscover their region’s own traditional techniques for water conservation. They dug rows of small pits in their fields to capture rainfall, which they filled with compost and manure, and into which they planted crops. Other farmers built stone terraces along the contours in their fields to capture rainfall and prevent runoff; some farmers did both. Over time, at least 140,000 farming households over 200,000 hectares or more were practicing these techniques, resulting in the revival of crop production, reestablishment of trees, shrubs and grasses, and the recharging of the area’s water table by a depth of five meters [Reij 2009].
Land management and hydrology
The concept of hydrological drought (as distinct from meteorological drought) helps explain the success of these age-old techniques to enhance surface and groundwater supply. Meteorological drought is the occurence of abnormally low rainfall for a given region. Hydrological drought is a consequence of meteorological drought – it happens when surface and ground waters run low thanks to a prolonged rainfall shortage compared to historical conditions for a region. Human water consumption for irrigation, industry and household use intensifies hydrological drought [Wada 2013].
Yet, as seen in the examples above, proper land management can raise the water table in an area in spite of occasional episodes of meteorological drought. In other words, while we cannot directly control when and how much rain falls, we can manage what happens to water once it reaches the ground.
Rain falling on much of our modern built environment is managed with ditches, gutters, drains and sewers designed to whisk it away as quickly as possible, rather than absorbing it in place. Farmland too is sometimes fitted with underground “tiles” (pipes) to drain fields, or with dikes to keep water out. Moreover, by the very absence of design, nearly all conventional farmland is so lacking in organic matter that it can barely absorb rainfall, which instead runs off the soil surface carrying soil with it.
In short, water hitting the ground in today’s world moves quickly. Stormwater moves in torrents over land seeking an outlet. That is until the outlet is full, at which point the water stagnates, rising like a bathtub, soaking and rotting property.
Ironically, the water we seek to drain away when there’s plenty becomes desperately lacking after the rain has stopped. Two sides of the same coin, flooding and drought often go hand in hand. By the same token, because a surfeit of impervious surfaces is at the root of these twin challenges, the solution of turning more land into a spongy surface helps resolve both problems at once. Spongy surfaces slow down water, allowing it to percolate into groundwater reserves.
Two sides of the same coin, flooding and drought often go hand in hand. By the same token, because a surfeit of impervious surfaces is at the root of these twin challenges, the solution of turning more land into a spongy surface helps resolve both problems at once. Spongy surfaces slow down water, allowing it to percolate into groundwater reserves.
Ecosystem restoration creates a spongy land surface by protecting soil with vegetation, thus allowing the soil to repair itself with biodiversity and organic matter, key ingredients of a good soil sponge. And beyond fostering drought and flood resilience, healthy ecosystems serve a myriadof protective functions, including cooling their surroundings, cleaning polluted water, drawing down CO2, and harboring the biodiversity that is the magic making ecosystems perform so many vital functions.
However, because the services rendered by nature go widely unrecognized or taken for granted, nature’s power as an ally is often shackled.
Human societies tend to value the potential benefits that a landscape might provide in a limited way, adjusting management practices towards desired outputs by maximizing the benefits gained from one or some of the services (often the provision of goods) leading to the loss of multifunctionality and the degradation of natural capital at the expense of human welfare [Schindler 2014: 230].
An example of a landscape with undervalued ecosystem function is a river floodplain. Because floodplains are often favored for agricultural, industrial, commercial or residential uses, rivers are constrained to their channels and their banks leveed, despite that a river and its floodplains are members of a single interdependent ecosystem. Through seasonal pulses of floodwater over the banks, like a heart pumping blood through a body, a river replenishes groundwater and nutrients throughout its floodplains, making these areas some of the planet’s most productive and biodiverse ecosystems [Sparks 1995]. Consider, for example, that the Amazon Basin is a floodplain, where fish biodiversity actually depends on riparian forest cover [Arantes 2017]. Fish swim into flooded forests during flood pulses and directly consume terrestrial floodplain vegetation (seeds, fruits, detritus).
…a river and its floodplains are members of a single interdependent ecosystem. Through seasonal pulses of floodwater over the banks, like a heart pumping blood through a body, a river replenishes groundwater and nutrients throughout its floodplains, making these areas some of the planet’s most productive and biodiverse ecosystems.
In addition to supporting robust biodiversity, floodplain ecosystems give water somewhere to go during severe flooding events rather than damaging cropland, houses or other properties. By allowing water time to infiltrate into the ground, floodplains recharge groundwater, thus alleviating future droughts [Opperman 2009]. Riparian ecosystems also serve as a migration corridor for birds and fish especially, and also as a refuge from the heat of more exposed areas.
In other words, intact river-floodplain ecosystems perform multiple ecosystem services and thus help us manage some of our most pressing societal problems if only we acknowledge the value of floodplains in these terms.
Making space for water
Given competing interests for floodplain property, some have argued for strategic partial reconnection of floodplains to the river by allowing portions of floodplain to flood, so that pressure elsewhere along the river during a flood may be alleviated [Opperman 2009].
For example, California’s Yolo Bypass was created in the early 1900s after the Sacramento River flooded several times and levees proved inadequate to protect the city [Sommer 2001]. The site of the bypass was historically the vast wetland floodplain of the Sacramento River and other nearby rivers and streams that had since been converted to agriculture. Today, the bypass reconnects the river to its floodplains.
Since1997, Yolo Bypass also features more than 16,000 acres of wildlife area, including seasonal and perennial wetlands, riparian forest, pasture, and seasonal crop production where fields are allowed to flood during winter. This solution has not only kept Sacramento dry during numerous high water events, it has also restored diverse populations of fish, bird, snake, mammal and other species to the area, while providing recreational and educational opportunities to nearby communities.
While the Yolo Bypass Wilderness Area exemplifies the large-scale engineered reconnection of a major river to its floodplain, many smaller stream floodplains benefit from the work of non-human engineers. Long considered the nemesis of ranchers and farmers alike, beavers caught damming irrigation ditches or flooding fields are often summarily trapped and killed. Yet in Elko, Nevada, beavers and altered livestock grazing regimes have brought stream beds back to life [Goldfarb 2018, Evans & Griggs 2015].
One rancher’s management change began by excluding cattle from grazing along Elko’s Suzie Creek – just during the hot season when plants are vulnerable. This allowed rushes, sedges and other vegetation to grow back, slowing water down enough for sediment to fill in the gouged out gully and raise the streambed back up to the level of its floodplain. Once beavers discovered their favorite food – willow – growing again at Suzie Creek, they moved in and have since built 139 dams there. These dams, in turn, raised the water table by about two feet, becoming the natural irrigation system for the ranch’s now lush riparian pastureland. The beaver dams proved vital in 2012-2015 when Suzie Creek kept flowing despite several summers of drought that left the rest of the region parched.
The beaver dams proved vital in 2012-2015 when Suzie Creek kept flowing despite several summers of drought that left the rest of the region parched.
At around the same time as Suzie Creek’s revitalization, across the Atlantic in Devon, England, a pair of Eurasian beavers were introduced to a wooded stream at the headwaters of the Tamar River [Puttock 2016]. A few years and 13 dams later, the beavers’ activity was filtering pollutants out of water passing through the dam sites and slowing the flow so as to minimize downstream flooding during storms. As in North America, beavers were once abundant in Europe, but by the 16th Century were wiped out in the UK. In recognition of beavers’ beneficial effects on hydrological systems, multiple reintroduction programs have begun establishing colonies across Europe and North America.
“Because of their abilities to modify streams and floodplains, beavers have the potential to play a critical role in shaping how riparian and stream ecosystems respond to climate change,” explain the authors of a recent study of potentially suitable beaver reintroduction sites [Dittbrenner 2018: 2]. The authors continue:
By damming streams, beavers create pond and wetland complexes that increase … species and habitat diversity, and therefore ecosystem resilience to climate-induced environmental change. Beaver impoundments slow stream velocity allowing sediment suspended in the water column to settle, aggrading incised stream systems, and reconnecting streams with their floodplains. The increase in surface water promotes groundwater recharge, storage, and supplementation during base flows. The increased geomorphic complexity also promotes higher thermal variability and cold-water refugia in deeper waters and in areas of downstream upwelling [Dittbrenner 2018: 2].
Furthermore, by repairing hydrological functioning and increasing a landscape’s overall level of moisture, beaver populations could literally dampen conditions for wildfire, which is intensified by drought [Maughan 2013].
Such vital ecosystem services provided by a keystone species like beaver in the era of climate change are nothing to scoff at. And while beavers are still trapped and killed for sport and by hunters or land managers whoconsider them a nuisance [Goldfarb 2018], castor canadensis is also increasingly accepted, as evidenced by more pro-beaver attitudes within the fish and wildlife departments of western states that had previously considered them mainly as pests. Beavers are discussed on state wildlife agency websites in terms of “living with wildlife,”[4] where beaver life histories are described, along with explanations of the benefits of beaver dams for landowners and for the landscape overall.
Similarly, Holland is learning new ways to live with water. In the Netherlands, literally “low country” due to much of its land area being at or below sea level, there is an age-old struggle with water and flooding, notably through the use of dikes. However, alarmed by recent flooding and the prospects of sea-level rise from climate change, the nation is undergoing a paradigm shift wherein the guiding principle for water management has become “make room for the river” [Pahl-Wostl 2006]. Among other tactics, certain areas are being “depoldered,” meaning dikes removed from low-lying areas and the land returned to wetland; people living in those areas are assisted in relocating [Bentley 2016].
Similarly, several cities in China are striving to “make friends with water” through adoption of the concept of “sponge cities” that aim to “retain, adapt, slow down and reuse” stormwater by increasing the porosity of urban surfaces, including increasing the amount of ecologically functional urban green space [Guardian 2018].
In our panic over increasing numbers of extreme weather events, we may grasp at familiar solutions – extra air-conditioning to shelter from the heat, higher levees to hold back floodwaters, more irrigation to combat drought, or logging forests to reduce wildfire fuel. While these measures may (or may not) temporarily bandage the situation, they increase the fragility of our built environment and usher us further down the path of climate chaos though unrelenting energy consumption and increasingly hobbled ecosystems. For our own sake, it’s time to make friends with nature and to acknowledge her superior power by partnering with instead of continuously fighting her.
Danger in the Arctic:The Urgency of the Climate Situation
Atmospheric carbon dioxide levels have increased from 315 ppm in 1958 to 410 ppm in 2018. This is the first time in at least 2 million years that these levels have been reached on planet Earth. In the last five years, the yearly rise is accelerating and is now about a 2.5 ppm increase per year. At that rate, CO2 will exceed 500 ppm before 2060.
Evidence of the impact of rising greenhouse gases includes the rapid loss of mountain glacier ice and the accelerating loss of the Arctic ice cap. As the Arctic Ocean loses its ice cover in the summers, solar insolation is warming the seawater and releasing methane gas to the atmosphere, particularly from the shallow East Siberian Shelf.
Even remaining at 400 ppm will continue to warm and further acidify all the oceans. The possibility of a runaway greenhouse spike from the release of trapped methane in methane hydrates and permafrost might appear to be slight at the present time, but it will increase as the oceans warm. A “Business as Usual” increase to 500 ppm will increase that possibility significantly to a very dangerous level. As well, oceans have absorbed 93% of of the planet’s heat increase due to the absorption of CO2.
Figure 1, below, summarizes the present situation. Fossil fuel burning is adding 10 gigatons (billion tons) of carbon to the atmosphere each year. About half of that is being absorbed by processes on the land and its soils, or in the oceans. That increase in CO2 is acidifying the ocean and ocean warming is slowing the ocean’s ability to absorb more. (It’s like warming your soda drink, which releases the carbon dioxide because gases are less soluble in warmer water.)
Reducing the stress of acidification on the oceans by burping out excess CO2 is important.. Eliminating fossil fuel emissions will help, but not nearly enough to return to 300 ppm within a safe time frame. Managing the land in better ways appears to be the only option to draw down enough carbon within a few decades. Is this a reasonable possibility?
Figure 1. Scenario 300: Land ecosystems must eventually draw down 20 gigatons (Gt) of carbon from the atmosphere on a yearly basis to reach 300 ppm by 2061. In 2017 10 Gt of CO2 were added to the atmosphere from burning fossil fuels, 5 Gt were removed from the atmosphere by the oceans, with an uncertain amount sequestered in soils. This leads to a net 5 Gt increase in atmospheric burdens. Numbers are approximate. Note that 2 Gt is approximately equal to one ppm (parts per million).
Scenario 300 proposes that restoration of degraded ecosystems of several types could capture enough carbon in soils to reduce the atmospheric CO2 concentration from about 410 ppm in 2018 to 300 ppm before Halley’s Comet returns in 2061. Youth born at the beginning of the 21st Century will be reaching their retirement years and it is hoped that they will have a future to look forward to. Seeing this famous Comet return for their first viewing could be a time of celebration for the many tough choices they had to face and a time of reflection at their successes and failures. How can older generations, from baby boomers on, help prepare them for what is coming?
To reach this monumental goal of 300 ppm of atmospheric CO2within a timeframe of about four decades would require good management on about half of the available lands. If the net soil carbon increase reached 20 billion tons, it would be possible to rapidly draw down atmospheric CO2 levels by several ppm every year. Is there enough land to do this? Does nature have the capacity when there is good human management?
Figure 2 below proposes that we could achieve a 4 ppm yearly drawdown if the land ecosystems captured 20 Gt of carbon each year. As the CO2 levels begin to drop, the reduced partial pressure of CO2 in the atmosphere will eventually allow the oceans to “burp” out some CO2 to the atmosphere. How much of the CO2 reduction in the atmosphere will be neutralized by CO2 coming out of the oceans? For our purposes I have estimated one third of the net difference between carbon captured by the soils and the fossil fuels being burned.
Figure 2. Calculation of net atmospheric drawdown of 4 ppm CO2 (8 Gt carbon) annually, accounting for 8 Gt of fossil fuel emissions and 4 Gt of carbon released .oceans, offset by 20 Gt sequestered in soils.
This number will vary depending on factors like water temperatures that may slow down the return to 300 ppm. The good news is that ocean acidification will gradually be reversed. Figure 2 shows a 20% reduction in fossil fuel burning, down from 10 Gt to 8 Gt. One third of the difference between land and fossil fuels is 4 Gt coming out of the oceans. If this estimate is too low, then the return to 300 ppm will take longer and ocean acidification will reduce faster. If the estimate is too high, the reverse is true. Figure 2 also shows that CO2 can come down by 40 ppm in a decade if we can learn how to capture 20 Gt of carbon in the land ecosystems and soils yearly.
Do Earth’s ecosystems have the capacity to capture 20 Gt of carbon yearly?
Figure 3, below, the “Road Map to 300,” gives several pathways to achieve this goal. While there are potentially 29 billion acres to work with (see table), this scenario uses only about half of this potential. If half of the available land is managed well in a holistic way, the Earth has plenty of capacity to store gigatons of carbon. In the Road Map, there are 13.5 billion acres in the “Half-Earth” column.
Figure 3. The Half-Earth Plan.
The final column shows that these 13.5 billion acres could capture 26 Gt of carbon annually. To do this requires 6 billion acres of grasslands to capture 6 Gt of carbon per year, averaging 1 ton / acre. 3 billion acres of farmlands and croplands can capture another 6 Gt by averaging 2 tons per acre per year. Forests are similar with 3 billion acres capturing 6 Gt of carbon and also averaging 2 tons C per acre per year.
Wetlands are extremely valuable and should be included to rehydrate all the other ecosystems. If 5% to 10% of the restored land were wetlands, the task of capturing carbon would be so much easier because they capture so much sediment and nutrients. A billion acres of wetlands can capture 6 GT of carbon annually.
The fifth row is called “Living Shorelines.” As the oceans continue to rise until after the continents begin to cool, we have the choice to “build a wall” and fight the rising tides …or… we can restore much of the coast to what it once was, a biodiverse area with salt marshes, seagrasses, kelp forests, mangroves and coral beds. I estimate that 0.5 billion acres of living shorelines would sequester 4 tons C per acre. Human civilization will have to move “uphill” for a while, but restoring living coastlines and populations of forage fish can bring back a fishing industry that has been largely depleted.
Humans will have to make huge changes in the ways we interact with nature and with each other. Our actions must become less competitive as we work together to enable symbiosis in the recovery of massively degraded ecosystems. It will be the greatest challenge in human history and will require several decades to accomplish.
It can be inspiring too, as vast areas of land come back to life. As these ecosystems restore the water cycle and build deep carbon rich soils, there can be sufficient food and water for the human population as well as the many other organisms we depend on. As CO2 levels fall and small water cycle processes like infiltration and transpiration increase, the continents will begin to cool. Eventually continental cooling and ice formation at higher altitudes will begin to cool the oceans. Soil formation on the land will reduce erosion and ocean “dead zones” will diminish. As ocean acidity begins to drop, shellfish will also be less stressed and more productive.
Figure 4 below shows some possibilities in the next four decades. Also in Figure 4, we see that it takes two decades to “ramp up” restoration processes. In the 2020’s CO2 levels only drop by 10 ppm (1 ppm / year). As more people get involved and land restoration accelerates, we see a 25 ppm drop in the 2030’s. By 2030 the amount of carbon being captured by soil processes reaches 20 Gt / year as shown in Figure 2. In the 2040’s and 2050’s we have reached the “Half- Earth” plan of restoring 13 billion acres on the planet. CO2 levels in the atmosphere are now dropping by 4 ppm per year or 40 ppm per decade. We can reach 340 ppm by 2049 and 300 ppm by 2059 according to this scenario.
Figure 4. Steps in the progress of Scenario 300.
Is this a practical plan? Will humanity be able to change this rapidly? Many cannot imagine such a complete turnaround of our management of the planet’s lands and biodiversity. Do we have a choice? As the news and evidence from the Arctic and Antarctic continues to shock us with the possibility of a runaway greenhouse gas scenario and rapidly increasing methane levels, we will continue to maintain “Business as Usual” at our own peril. This plan may not be “practical” in the present cultural context, but there are serious consequences already evident in abundance if we do not embark on a plan like this.
Human extinction is possible as we proceed to 500 ppm. Human extinction may even be probable, but it is not necessary. We can turn this around by using what we already know about restoring the land and becoming a symbiotic catalyst to rehydrating and cooling the planet. We can do this. …so let’s get started.
Figure 5 below shows a Strategy Map for the “Lower 48 States” in the USA. This map should be considered a brainstorming tool and not a proposal. It does give a general plan for the 2 billion acres in this area. This area now burns enough fossil fuel to put about 2 billion tons of carbon into the atmosphere every year.
Figure 5.Strategy map in the U.S. Note: Brittle landscapes are drier lands that experience periodic dry seasons annually.
How could these lands capture 3 or even 4 billion tons of carbon and make the USA a net carbon sink? As we learn to capture carbon on the national level, we can also learn how to make North America and other continents net carbon sinks. Remember, the goal is to restore ecosystems capable of capturing 20 billion tons of carbon per year. When that level is reached, carbon dioxide will be falling out of the sky, dropping by several ppm every year.
Resources – Building a Case for Scenario 300
Pilot Analysis of Global Ecosystems: Grassland Ecosystems, by Robin White, Siobhan Murray, Mark Rohweder, World Resources Institute, 2000. This PAGE report is the source of my “Potential Billions of Acres” column in Figure 3. The “Half-Earth” column is the area needed to achieve the scenario’s goal of 300 ppm by 2061. Thus, with our estimates based on using only 50% of available land in various ecosystems, there is a good deal of latitude in each instance.
Wetlands are a special case. The goal of restoring 1 billion acres of wetlands can be done if wetlands are incorporated into the plans for restoration of other systems. Wetlands can raise the water tables several feet in dry areas as we have seen in Zimbabwe and in Nevada, improving grazing opportunities there. Wetlands established in areas with higher rainfall can also improve resiliency of forests and farms. If 5% to 10% of these lands were managed as wetlands, the benefits would be enormous and a lot of carbon would be captured, too. This percentage represents about 1 billion acres.
Bio4climate Videos: Biodiversity for a Livable Climate (bio4climate.org) is a non-profit in Massachusetts dedicated to restoring the lands, waters, and biodiversity of the planet. We have produced eleven conferences in our four years exploring how to do this and continue to learn at a rapid rate. Our website has documented these conferences and their diverse range of speakers and participants. All of our speaker presentations have been video recorded and are available free online (see an introductory playlist at https://www.youtube.com/playlist?list=PLsWWRqCX9eSYwxFIgBDGMMQXFvK2mwR06). There are now almost 200 videos on the main page or program page of each conference with over 108,000 views on YouTube (see https://bio4climate.org/conferences). Many of these will be highlighted as resources below.
Walter Jehne is an Australian microbiologist and climate scientist. He has outlined a global strategy for ecosystem restoration in response to the Virgin Earth Challenge. His plan also shows that 20 billion tons of annual carbon sequestration by ecological restoration is possible if humans have the will to make it happen.
Rangelands, Grasslands, and Deserts (see discussion of Allan Savory’s work in Compendium, Vol. 1 No. 1, pp. 59 ff.): The goal is 6 billion tons carbon capture per year (13 billion acres available – Half-Earth Plan > 6 billion acres x 1 ton C / acre per year).
Much of world’s rangelands are degraded and some are severely degraded. Allan Savory describes areas of seasonal rainfall as “brittle” environments. Areas that receive less than 20 inches of rainfall a year tend to be more brittle, but seasonality is important. In the “lower 48” United States lands west of the 100th meridian generally receive less than 20 inches of rain and are more brittle than lands east of this line. Most of these areas globally were once thriving biodiverse grasslands, but many are now growing “deserts” with vast areas of bare ground. Grasslands require grazing, hoof disturbance, and nutrients in the form of dung and urine from dense animal herds. The herds are not allowed to return to the impacted area until it has adequately recovered.Depending on brittleness, this might range from a few weeks to two years. While many rangelands have not been very productive, Allan Savory has demonstrated that these lands can come back within three or four years if managed holistically.
Many of these lands are now bare ground most of the year. Either they are devoid of animals, subjected to continuous grazing, or used as monoculture croplands for grains, cotton, etc. These croplands are usually high input operations using chemical fertilizers (NPK), pesticides, and irrigation water. Another problem in these brittle landscapes can be fire: both wildfires and intentional burns add much CO2 to the atmosphere.
The Africa Centre for Holistic Management in Zimbabwe.Allan Savory’s contributions continue in Zimbabwe workingwith the Africa Centre. He is most impressed by the rising water tables and the return of wildlife to the areas being grazed by communities using movable kraals (corrals). Water cycle improvements in infiltration ensure that a considerable amount of carbon is going into the ground: the raised water tables enable more vegetation and soil biota to grow.
The “Road Map to 300” table shows Grasslands and Deserts covering 13 billion acres (Fig. 3). If only half of this land were managed in a way that captured 1 ton of carbon per acre each year, 6 billion tons of carbon per year would be added to the soil. Is 1 ton of carbon captured per acre possible with good management? We are learning that it is not only possible but can also be very profitable.
Richard Teague from Texas A&M found many examples using multi-paddock grazing where carbon capture was between 1 and 2 tons per acre per year. Dr. Teague spoke at the first Bio4climate conference at Tufts University in 2014. Link is below:
Teague also gave a more detailed presentation at the Quivira conference in 2015.
Seehttps://www.youtube.com/watch?v=crG4L4J-OEg His summary at Quivira [37:30 to 39:53 minutes] emphasized that academics must work with leading ranchers and farmers to document how effective holistic management can be. He estimates that these ranchers and farmers are 20 to 30 years ahead of the academics in their ability to restore the land.
Christine Jones’ work on the Colin Seis ranch in Australia has involved the study of pasture cropping (grazing mixed with cover crops and crops sown into pastures). She is also measuring accumulated carbon levels down to two feet. Most studies don’t look past six inches of soil. The Seis Ranch has been getting about 1.8 tons carbon per acre per year over ten years, but in the last two years it has risen to 3.6 tons C per acre per year. It is important to note that no fertilizers are being used and mineral density in the crops is improving as carbon levels rise. Note that fungi “mine” rocks in the soil for minerals.[16] See http://www.amazingcarbon.com/PDF/JONES%27CarbonThatCounts%27.pdf
Gabe Brown in North Dakota has had even better results when he finally began mixing grazing with cover crops and crops sown into pasture. On some pastures Gabe is now capturing carbon at about 7 tons per acre per year. This area only receives 16 inches of annual rain and has a short growing season. The importance of mycorrhizal fungi is to be emphasized in both the Seis and Brown studies. By refusing to use chemical fertilizers, pesticides, and antibiotics, Gabe Brown’s lands are rich in mycorrhizal fungi and soil insectsand costs of production are far lower.
Gabe’s TED talk link: https://www.youtube.com/watch?v=QfTZ0rnowcc [11:15 – 15:30 minutes]. By integrating cover crops and multi-species grazing, he has seen organic matter rise from 4.2% to 11.1% from 2006 to 2013. This change represents a 6.9% increase in organic matter in seven years. I calculate that to be about 7 tons of carbon captured per acre per year. Gabe’s farm is in an area with 16 inches of rainfall and a 5-month growing season.
David C. Johnson from New Mexico State,speaking at the “Climate Reckoning” conference produced by Bio4climate in November 2017,amplified the message about fungi. In his research, his results show a capture rate of 10.7 tons C per hectare per year (4.33 tons C per acre per year). His method was no-till, no-chemicals, but he does not use grazing animals. In his presentation, he referred to Gabe Brown’s work. David’s calculation for carbon capture (adding his BEAM fungal/bacterial balancing technique) at the Brown Ranch is 20.63 tons C per hectare per year or 8.25 tons C per acre per year! Seehttps://www.youtube.com/watch?v=neIIPRRnXQQ&feature=youtu.be [28:10 – 36:15 min.]
“Make Soil – End Global Warming,” Figure 6, shows how important biodiversity is to stabilizing the climate. Look for the key players: grazing herds, perennial grasses, dung beetles, mycorrhizal fungi, carbon farmers … and more.
Figure 6. The importance of biodiversity.
Grasslands Summary:We are finding many examples of grasslands exceeding the 1 ton of carbon per acre yearly capture rate. If we can bring one half of the land in these brittle environments under good management within two decades, we should exceed the 6 billion tons annual carbon capture that is our goal for these lands. Integrating wetlands and wet meadows with these grasslands is another way to increase the carbon capture rates, which we will explore below. We are discovering that good management can work even in very brittle lands and has been demonstrated on tens of millions of acres on five continents. We know how to do this.
The overarching issue is that billions of acres of degraded land are still being overwhelmed by chemicals, pesticides, tillage, and/or continuous grazing, often at a financial loss to the landowner. The challenge is to bring these restorative possibilities to range managers everywhere.
Regenerative Farms and Permaculture.Goal is 6 billion tons carbon capture per year.
(6.3 billion acres available – Half-Earth Plan > 3 billion acres x 2 tons C / acre per year)
Agricultural lands are generally less brittle and receive more rainfall than rangelands. Because of this there is greater opportunity to capture carbon. Using regenerative techniques with the goal of building soil with healthy mycorrhizal networks should yield 2 tons of carbon sequestered per acre per year. If we are able to reach this goal on 3 billion acres (half of these lands) that would capture another 6 billion tons of carbon annually.
Most agricultural lands have been the victim of severe erosion due to tillage and synthetic chemical use. They have lost over half of their organic matter, which makes it difficult to hold water in these soils. There is a huge opportunity to reverse this process. It would require far less dependence on chemical fertilizers and pesticides.
Fungal networks are essential to bring nutrients to the plants. Feed lots must be ended: the animals are needed out on the land (both rangelands and croplands) as a nutrient pump distributing manure, urine, and saliva, as well as a food source for humans. Raising 100% grass-fed beef and ending ethanol subsidies would reduce incentives for grain production and lead to a more nutrient-dense diet.
Providing funding to ranchers and farmers on the land for ecosystem services would accelerate this conversion. Providing $20 per acre for good management on 12 billion acres described in this scenario would require $240 billion dollars. This is about a quarter of the world’s military budget. Because climate change is the greatest threat to global security, this is a relatively small price to pay.
Greg Judy is a grazier in central Missouri.He has seen an increase of as much as three inches of topsoil created in four years. An inch of this rich soil contains 8 tons or more of carbon per acre. Three inches over 4 years yields 6 tons C per acre per year.
Judy counts earthworms on the land he manages and finds as many as 17 piles of earthworm castings per square foot, or 700,000 casting piles per acre. His grazing plan includes a very dense herd in an area for a short duration (1 day) and 2 to 3 months recovery time. The manure deposited is significant and the trampling of the grass brings a great deal of biomass in contact with the soil to feed the earthworms. Dung beetles are also at work, drying out the dung and burying most of it in balls several feet deep. While the beetles are deepening carbon rich soils, the earthworms are mixing the same soil through many levels.
Greg Judy has written two books explaining his methods and includes his financial planning. These books are great sources for land managers, farmers, and ranchers:
No Risk Ranching: Custom Grazing on Leased Land
Comeback Farms: Regenerating Soils, Pastures and Profits with Livestock Grazing
Greg discusses biodiversity restoration in his lecture at the Virginia Association for Biological Farming Conference. Seehttps://www.youtube.com/watch?v=W6HGKSvjk5Q. Some good clips from this talk include [10:00 to 20:00 minutes] and [49:50 to 62:30].
Joel Salatin is a farmer in the Shenandoah Valley of Virginia.He has written many books on farming. He describes how 8 inches of topsoil have accumulated over bedrock in a decade (2000 to 2010). At 8 tons per acre per inch of topsoil, that’s 64 tons C in ten years. Five tons C per acre per year is a conservative estimate. (Remember, Scenario 300 only requires 2 tons per acre on half of the agricultural lands worldwide to see a major drawdown of atmospheric CO2.) Salatin’s description can be found in his book titled, The Sheer Ecstasy of a Lunatic Farmer[Ch. 1, pp. 2-15].
Mark Shepard is a farmer and grazier in Wisconsin. He is planting a diverse blend of nut and fruit trees mixed with perennial crops, mushrooms, and grazing. He is rebuilding soils using ideas like the permaculture techniques described by Bill Mollison[17] years ago. Shepard’s book, Restoration Agriculture: Real World Permaculture for Farmers, is very detailed and shows farmers how to grow nutrient dense food. This approach builds extensive mycorrhizal fungi in the soil which is a good sign of carbon rich soils. The perennial plants feed much carbon in the form of sugars and enzymes to the soil microbes and fungi.
Shepard also writes about his financial plan and shows how it is more profitable than the corn system most farmers now use in his area. If we subsidized “carbon farming” methods like Shepard, Salatin, Judy, and others are using, the transition to these methods could happen within the two decades required in Scenario 300. This is especially true if we stopped subsidizing soil-killing chemical agriculture.
Lands managed using permaculture, perennials, and holistic planned grazing would be amazing sites for long-term academic study over the time span of a decade or two. Farmers can rarely afford to do these studies; furthermore, they should be rewarded for ecosystem services, not charged for expensive lab work. Understanding the value of these systems should be a national priority. We are now subsidizing the “death of soils” by encouraging more chemical use. Using some of these funds for restoration studies would certainly be a step in the right direction.
Forests:Goal is 6 billion tons carbon capture per year. (7.1 billion acres of forest available – Half-Earth Plan > 3 billion acre x 2 tons C / acre per year.)
The “Half-Earth Plan” for forests has numbers similar to regenerative agriculture. If three billion acres were managed “holistically” to improve water and nutrient cycles, then 2 tons carbon capture per acre each year would achieve the goal.
Our understanding of how a healthy forest works is still very sketchy, but we are discovering that old-growth forests can capture more carbon and nitrogen than younger forests. Clearcutting every 40 years and burning the debris before replanting has been devastating to forest soils, but it was considered the best way to manage forests and remained largely unchallenged until the 1980s. What we have learned since gives hope that restoring forests can be a big part of Scenario 300.
Replanting clearcuts and selectively cutting in standing forests would be a good start. Allowing many mature forests to advance into “old-growth” stage is another possibility. Trees can feed mycorrhizal fungi and microbial networks. Grazing insects and nematodes release nitrogen for the trees slowly and their “bug poop” contains progressively more stable carbon compounds like humus and lignin. As soils in older forests mature, the amount of stable carbon in the ground continues to increase. Selectively cut wood products can hold their carbon for decades. Holistically managed forests that demonstrate the value of ecosystem services should qualify for subsidies mentioned above for well-managed ranches and farms.
Jon Luomawrote The Hidden Forest: Biography of an Ecosystem (1999). This excellent work describes research at the Andrews Forest in Oregon,where many discoveries have been made since its protection in 1948. Soil processes are described in great detail in Chapters 5 and 6.
Joan Maloof, writing in her book, Nature’s Temples: The Complex World of Old-Growth Forests (2016), has focused on old-growth forests and finds that “increasing the time between logging events for managed forests and preserving old-growth forests, the carbon stored in forests and kept out of the atmosphere could theoretically double (pp. 36- 37).
Paul Stametshas done amazing research on fungi for decades and gives us many insights about forest restoration from his knowledge of mycorrhizal fungi in his book, Mycelium Running: How Mushrooms Can Help Save the World(2005).
If forests contained 10% beaver meadows and other wetlands, the average carbon capture rates and biodiversity would rise. Rising water tables would also reduce fires, a major source of greenhouse gases. This theme of integrating up to 10% wetlands into our restoration plans for improving carbon sinks keeps coming up in this document. Let’s look at wetlands more closely.
Wetlands.1 billion acres restored globally is essential: (Goal is 6 billion tons carbon capture per year – 1 billion acres x 6 tons C / acre per year.)
The loss of wetlands is a big part of the climate crisis. If we are serious about reversing climate change we must learn how restore these wetlands. It is estimated that the continental United States was once 10% wetlands due largely to beaver activity (see Alice Outwater, Water: A Natural History 1996). Extending the 10% figure to the rest of the globe, 2 B acres of wetlands are a real possibility. If we make our goal half of that over the next 40 years, it would require a billion acres of new wetlands. The scenario uses a carbon capture rate of 6 tons per acre per year, which is much higher than other ecosystemsbut justified by the following examples.
Steven Apfelbaum has created wetlands which sequester 7 to 12 tons per acre in North Carolina and Illinois. He also stresses the importance of covering peatlands that have been exposed to the air by dropping water tables. Drying out these carbon rich soils can lead to huge fires. He agrees with Walter Jehne about the importance of reducing landscape scale fires, a huge source of atmospheric CO2. Apfelbaum spoke at the first Bio4climate conference at Tufts in 2014. See https://www.youtube.com/watch?v=gYq8WsLyPlg [9:58 – 20:04 minutes].
Carol Evans & Jon Griggs have encouraged beaver activity on Maggie and Susie Creeks in Nevada. The wet meadows now have 2 to 3 feet of mucky black soil and the water table in the area is 3 feet higher than 15 years ago. This muck contains significant carbon, at least 100 to 150 tons C per acre increase in 15 years. (This is what our climate scientists should be focused on.)
My estimate for Susie Creek is a yearly increase of 6 to 10 tons C per acre. Carol Evans is a fisheries biologist who has been studying these streams for several decades. Jon Griggs is the manager of Maggie Creek Ranch and is thrilled to see how the the vegetation can come back even in an area averaging less than 10 inches of precipitation per year. Link to their talk at our 2015 Tufts Conference: https://www.youtube.com/watch?v=lR7w9Tritj8&t=67s
While creating wetlands in populated areas – beavers often have different priorities than humans – we are learning how to work with them. The larger opportunity is in the desiccating and abandoned brittle landscapes. These lands need animal activity. Wild herds would help but rebuilding them will take awhile. Ranchers can bring in cattle, but they are often worried about water availability.
The Nevada case mentioned above is a great example of a regeneration strategy. Ranchers had a good management plan and kept the animals moving. As the creeks began to hold water longer, the beaver appeared. Without wood in those first years, they used rocks and mud and built deep channels to create permanent pools so they could survive the hot summers. The land holds much more water and the small, local water cycle is being enhanced (see reference below re Small Water Cycle). Trout and freshwater mussels are making a comeback in many of these streams.
Chernobyl – Restoration after Human Evacuation. The area around Chernobyl was a huge grain operation until the nuclear accident in 1986. Over 1000 square miles were evacuated and the area has become a curiosity for scientists who are doing research with strict exposure limits. This Exclusion Zone is now a biodiverse ecosystem with abundant wetlands and wet meadows due to beaver activity. Many threatened species have returned and are doing well including otters, moose, wild horses, and a diverse mix of water bird species. The wolf populations are doing well here approaching a hundred animals.
Because of radiation, most humans will avoid this area for a long time. Consequently, there is a great opportunity to observe how nature rehydrates the land and captures a great deal of carbon. Beaver once roamed most of central Asia all the way from France to China and Mongolia before they were trapped out in the 1300’s. Imagine rehydrating most of Asia’s degraded grasslands by nurturing beaver activity to raise water tables while restoring wild herds along with local grazing herds. Asia would become a net carbon sink. Check out this amazing Nature Documentary, Radioactive Wolves Of Chernobyl: https://www.youtube.com/watch?v=yzza7Aouzn8[Historical wetlands summary at 13:25 to 17:30 minutes].
Michal Kravčík, an engineer from Slovakia, has been restoring landscapes and educating communities on several continents. He wants to restart what he calls the “small water cycle” in as many areas as possible. Kravcik asserts that water is now flowing off the continents faster than it is returning from the oceans as rain. This would explain why the continents have been slowly desiccating (drying out) for centuries. By his calculation, the runoff exceeds the rainfall coming from the ocean, and the difference exceeds the volume of Lake Erie, every year.Michal and others have been helping to slow the water down, increasing infiltration and transpiration, and cooling the planet.
WetlandsSummary: Wetlands are also a must in other landscapes.If California returned 10% of its agricultural lands to wetlands and beaver meadows, it would have far fewer drought problems. If forest management required slowing of water and wetlands to raise water tables, the forests would have better immunity to pests, and wildfires would be reduced. Fire is not a very good tool if our aim to to reduce greenhouse gases. Keeping water at higher altitudes longer and having it go through many transpiration cycles (Kravčík’s Small Water Cycles) before it reaches the sea will cool the system whilebuilding many water storage “bank accounts” for the inevitable dry periods.
Living Shorelines. Potential is 2 billion tons carbon capture per year (0.5 billion acres needed x 4 tons per acre per year)
The coastlines are very developed in many areas with large cities and little biodiversity. The sub-surface continental shelves have been plagued with dead zones from soil runoff and frequent trawling of the bottom by large fishing fleets. We are also already experiencing the impact of rising oceans and can expect sea level rise to accelerate for the next few decades.
Our goal of restoring ecosystems, building soils, and rehydrating continents will eventually reduce greenhouse gas levels and cool the continents, but oceans still contain most of the heat energy that the ‘greenhouse blanket’ has captured in the last century. Until much of this energy can escape through an atmosphere with reduced levels of carbon dioxide, methane, and other GHGs, we will be faced with continuing melting ice in Antarctica, Greenland, and high altitude glaciers. In Scenario 300, it will still take about four decades before we attain an atmospheric CO2 level of 300 ppm. Hopefully at that 300 ppm level, we will begin to see some glaciers reforming in the higher altitudes and latitudes. It will be a big day when the Arctic Ocean begins to build up ice again, but the lag time might take many decades longer.
We expect at least a 5 or 6 foot sea level rise before the year 2100 even with the swift implementation of Scenario 300. However, without Scenario 300 the situation would be far worse. We could build walls to keep the ocean out, but the expense of this will exhaust resources needed for other challenges. Could we allow the ocean to rise and create large areas of saltwater ecosystems to supplement our carbon capture? Restoring these ecosystems and bringing back the plankton-eating forage fish could reinvigorate the coastal food webs. Strategically limiting coastal development and learning how to help cities thrive in a more aquatic environment is needed, now.
There are already many groups working on this possibility. Restoring salt marshes, sea grasses, mangroves, and corals is possible. Strategic thinking many decades into a rapidly changing future is already being done. In November 2016, Bio4climate organized an oceans conference at Harvard with the title: “Restoring Oceans, Restoring Climate,” see
This conference had over 20 presentations. The videos are available at the above link. A small sample of speakers includes Tom Goreau from the Global Coral Reef Alliance giving examples of his work to protect and restore coral reefs. Alfredo Quarto updated us on Mangrove restoration projects on several continents. Anamarija Frankic of UMass Boston and the Green Harbors Project has been reestablishing oyster and mussel reefs along the urban coastlines.
George Buckley from Harvard gave the natural history and importance of horseshoe crabs. Dwayne Shaw, from Downeast Salmon Federation, spoke about bringing back river herring in New England streams. John Todd and Brian Von Herzen gave several examples of ecological design to bring back the ocean food webs. Katherine Deuel of the Pew Charitable Trust spoke about ‘forage fish’ like sardines and menhaden which are the key to re-establishing the big fish to places like New England. The Pew group is a leader in getting coastal communities to think about creating “Living Shorelines.”
As the oceans warm and the seas rise, might we see the possibility of mangroves and corals moving into new areas farther north? Could these Living Shorelines with many diverse ecosystems become significant carbon sinks? Let’s find out. We have learned a great deal in the last decade about these possibilities.
Another example of strategic thinking regarding climate change is the Rice University Severe Storm Center (SSPEED). It is a collaboration between the Rice civil and environmental engineering departments and many environmental and community organizations in the Houston area. The SSPEED Center is working to protect as much coastal land as possible south of Houston along the Gulf Coast as a buffer to major hurricanes. Development and impermeable surfaces in the Houston area has made it very vulnerable to flooding: Hurricane Ike and Hurricane Harvey have been devastating. As sea level rises, there is now a lot of interest in rethinking land use on the coast. See the proposal for a Lone Star Coastal National Recreation Area: http://www.sspeed.rice.edu/lscnra. Jim Blackburn, the director at SSPEED, is a lawyer, environmental engineer, and nature enthusiast, and is featured in the video at this link: https://www.youtube.com/watch?v=P0YewbjwXq8
Can living shorelines be a significant carbon sink as the sea level rises? We are just beginning to study the possibilities, so the jury is still out. It is hopeful, though, considering how much interest has increased in a very few years.
Summary:
This period is the most challenging time in human history. The consequences for not facing this challenge are huge. Sea-level rise will be significant and will take decades to reverse under the best scenarios. As the Arctic Ocean becomes ice free and its waters become warmer, we face an increasing probability of a runaway methane spike. Now is the time to get started!!!
Nature’s Ecosystems have the capacity to reduce atmospheric CO2 levels to 300 ppm within a few decades.Enhancing biodiversity and Holistic Management are essential for this to happen. Humans must adopt a very different approach to ecosystem management on half or more of the available lands.
A determined human effort with millions of people involved and eager to restore these symbiotic processes, just might lead to 300 ppm again by the return of Halley’s Comet in 2061. It can certainly be a rewarding project for those involved.
In this study, long-term phosphorus fertilization limited the extent to which the genes and proteins of microbial communities were allocated to degrading recalcitrant soil phytate to acquire phosphorus. In phosphorus-deficient soil, on the other hand, the genes responsible for breaking down recalcitrant substrate to acquire phosphorus were more prevalent in microbial communities. In other words, microbial communities can adapt genetically to different levels of nutrients in the soil in order to continue meeting their nutritional requirements. This adds to the body of evidence that fertilizer use impairs the inherent qualities of a living soil to nourish the plants growing there.
A greater than fourfold increase in the gene abundance of 3-phytase was the strongest response of soil communities to phosphorus deficiency. Phytase catalyses the release of phosphate from phytate, the most recalcitrant phosphorus-containing compound in soil organic matter. Genes and proteins for the degradation of phosphorus-containing nucleic acids and phospholipids, as well as the decomposition of labile carbon and nitrogen, were also enhanced in the phosphorus-deficient soils. In contrast, microbial communities in the phosphorus-rich soils showed increased gene abundances for the degradation of recalcitrant aromatic compounds, transformation of nitrogenous compounds and assimilation of sulfur. Overall, these results demonstrate the adaptive allocation of genes and proteins in soil microbial communities in response to shifting nutrient constraints [Yao 2018: 499].
In conclusion, our proteogenomics results provide systems biology insights into the adaptation of soil microbial communities to different levels of phosphorus availability in a humid tropical forest environment. Phosphorus deficiency significantly enhanced the genetic capabilities of microbial communities to extract phosphorus from phytate and, to a lesser extent, from nucleic acids and phospholipids. Long-term phosphorus fertilization altered the allocation of genes and proteins by microbial communities to acquire carbon, nitrogen and sulfur from a variety of substrates. The results suggest that the selective degradation of recalcitrant substrates, including phytate in phosphorus-deficient soils and aromatic compounds in phosphorus-rich soils, is an important means for microbial communities to balance their elemental requirements. The adaptive allocation of genes and proteins for acquisition of these nutrients in different soils can be explained as an optimal foraging strategy by which microbial communities maintain efficient growth under resource limitation [Yao 2018: 505].
Despite increasing residue input in annual crop production systems, N fertilization does not increase soil organic carbon (SOC) over time because N fertilization also increases organic carbon (OC) decay. This study also shows that belowground OC inputs contribute to soil carbon sequestration more than aboveground OC inputs to the soil.
When all phases of the crop rotations were evaluated over the long term, OC decay rates increased concomitantly with OC input rates in several systems. Increases in decay rates with N fertilization apparently offset gains in carbon inputs to the soil in such a way that soil C sequestration was virtually nil in 78% of the systems studied, despite up to 48 years of N additions [Russell 2009: 1102].
Across all systems, SOC storage was significantly correlated with the quantity of belowground OM [organic matter] inputs (P < 0.01, both sites). In contrast, SOC was not correlated with the quantity of aboveground inputs (P = 0.45, Nashua; P = 0.55, Kanawha) [Russell 2009: 1111].
This study highlights the importance of incorporating both production and decomposition processes, as well as the location (above- or below-ground) of detrital inputs into models of N-fertilization effects on soil C dynamics in agroecosystems. These results are highly relevant for evaluating the potential of N fertilization to mitigate the effects of removal of organic-matter ‘‘residue’’ from the system for bioenergy production. Our data suggest that the stimulation of OC decomposition by the addition of fertilizer N would likely counteract the positive effects of N fertilization on inputs of OC to the soil, at least for annual crops. Given the current quantity of N that is applied over such a large area, management strategies that can maintain high yields and also reduce N-fertilizer use would also have beneficial environmental consequences. Our study indicates that selection of crops for higher belowground NPP [net primary production], in rotation with crops that fix N, could maximize both yields and soil C sequestration without excessive N-fertilizer additions [Russell 2009: 1111].
In this study, nitrogen fertilization altered the relative abundance of fungal taxa in the rhizosphere, increasing fungal genera with known pathogenic traits, and decreasing a fungal phyla (Basidiomycetes) known to break down lignin, thus important for carbon cycling in the soil.
Fungi play important roles as decomposers, plant symbionts and pathogens in soils. The structure of fungal communities in the rhizosphere is the result of complex interactions among selection factors that may favour beneficial or detrimental relationships. Using culture-independent fungal community profiling, we have investigated the effects of nitrogen fertilizer dosage on fungal communities in soil and rhizosphere of field-grown sugarcane.The results show that the concentration of nitrogen fertilizer strongly modifies the composition but not the taxon richness of fungal communities in soil and rhizosphere. Increased nitrogen fertilizer dosage has a potential negative impact on carbon cycling in soil and promotes fungal genera with known pathogenic traits, uncovering a negative effect of intensive fertilization [Paungfoo-Lonhienne 2015:just 1].
There is a prevailing view that global food and fiber production will continue to expand because of modern agricultural management systems with improved cultivars and intensive chemical inputs dominated by synthetic ammoniacal fertilizers. The use of these fertilizers has led to concerns regarding water and air pollution but is generally perceived to play an essential role for sustaining agricultural productivity, not only by supplying the most important nutrient for cereal production but also by increasing the input of crop residues for building soil organic matter. The scientific soundness of the buildup concept has yet to be substantiated empirically using baseline data sets from long-term cropping experiments. The present paper and a companion study by Khan et al. (2007) provide many such data sets that encompass a variety of cereal cropping and management systems in different parts of the world. Overwhelmingly, the evidence is diametrically opposed to the buildup concept and instead corroborates a view elaborated long ago by White (1927) and Albrecht (1938) that fertilizer N depletes soil organic matter by promoting microbial C utilization and N mineralization. An inexorable conclusion can be drawn: The scientific basis for input-intensive cereal production is seriously flawed. The long-term consequences of continued reliance on current production practices will be a decline in soil productivity that increases the need for synthetic N fertilization, threatens food security, and exacerbates environmental degradation [Mulvaney 2009: 2308].
The symbiosis between mycorrhizal fungi and plants drive carbon and nitrogen cycles. Fungi demand carbon exudate from plants in exchange for nitrogen and other nutrients retrieved and transported from the soil. The “liquid carbon” exuded from plant roots feeds mycorrhizal fungi and many other soil microbes, while also becoming stabilized in soil aggregates and humus. Jones explains that when this mycorrhizal exchange is inhibited by N fertilizer, which allows plants to absorb nitrogen “for free” (without providing liquid carbon in exchange), this reduces the flow of carbon into the soils, which in turn diminishes fungal networks and their delivery of micronutrients to plant hosts, and results in carbon-depleted soils.
Despite its abundance in the atmosphere, nitrogen is frequently the most limiting element for plants. There is a reason for this. Carbon, essential to photosynthesis and soil function, occurs as a trace gas, carbon dioxide, currently comprising 0.04% of the atmosphere. The most efficient way to transform CO2 to stable organic soil complexes (containing both C and N) is via the liquid carbon pathway. The requirement for biologically-fixed nitrogen drives this process.
If plants were able to access nitrogen directly from the atmosphere, their growth would be impeded by the absence of carbon-rich topsoil. We are witnessing an analogous situation in agriculture today. When inorganic nitrogen is provided, the supply of carbon to associative nitrogen fixing microbes is inhibited, resulting in carbon-depleted soils.
Reduced carbon flows impact a vast network of microbial communities, restricting the availability of essential minerals, trace elements, vitamins and hormones required for plant tolerance to environmental stresses such as frost and drought and resistance to insects and disease. Lowered micronutrient densities in plants also translate to reduced nutritional value of food [Jones 2014: 2-3].
Jones further explains how to modify agricultural practices to protect and build the soil: maintain year-round living ground cover, limit nitrogen and phosphorus fertilizer input, promote plant and microbial diversity, and integrate livestock into crop production systems.
There are a number of agricultural practices that will enhance fungi colonisation. Wherever possible, of course, the full range of critical soil health processes that govern productivity should be allowed to regenerate agricultural ecologies naturally. It may, however, be necessary or more practical to inoculate seed with fungi spores in order to recover degraded soils. A number of farmers in the Great Southern agricultural region of Western Australia are undertaking this course of action. Finding themselves confronted with an unsustainable spiral of ever-increasing commercial fertiliser costs and uneconomic or diminishing crop yields, it was realised that a different approach needed to be taken. In recent growing seasons, seed has been inoculated with commercial fungi spores just prior to planting. While it is still too early to provide statistically robust outcomes and, bearing in mind that there are no “silver bullets” in agricultural production, the indications are that mycorrhizal fungi are promoting improvements in crop vitality, yield and soil condition [Johns 2014: 4].
Given the global decline of reserves of both rock phosphate and fossil fuel, this study poses the question – to what extent can microbial inoculants replace/reduce the use of synthetic fertilizer? The authors find that “dryland agriculture can benefit most from biofertilizers [microbial inoculants used as fertilizers]. Due to climate change, in the future there will be even more dryland areas globally. Biofertilizers are thus a promising option for sustainable agriculture” [Schutz 2018: 11]. More specifically:
Our comprehensive meta-analysis with studies from all over the world revealed that biofertilizers were found to be most effective in dry climates. Biofertilizer also improved PUE [phosphorus use efficiency] and NUE [nitrogen use efficiency] greatly. Furthermore, we found that biofertilizers possessing both N fixing and P solubilizing traits have the highest potential to improve the crop yields. Interestingly, AMFs, known for facilitating P nutrient uptake in plants, were on par with applications of biofertilizers with the combined traits of N fixation and P solubilization, indicating the big potential of AMFs as sole biofertilizer for most crops and climatic situations [Schutz 2018: 5].
The paper argues for the use of bacterial and fungal inoculants in combination with organic amendments and cover crops to regenerate degraded soils. In order to produce enough food for a growing global population on ubiquitously degraded soils, synthetic fertilizers will be in increasingly high demand. However, these fertilizers require copious amounts of non-renewable energy to manufacture, and become pollutants when used. Here, the authors explain how bacteria and fungi make nutrients available to plants and how facilitate soil aggregation.
Given that nitrogen and phosphorus are the most limiting nutrients for crop growth, that global phosphorus supplies are becoming exhausted while the human population rapidly expands, and that arbuscular mycorrhizal fungi (AMF) symbioses improve crop phosphorus acquisition, AMF symbioses have a major role to play in current and future crop production.
The potential of AMF to help increase global food security lies in the fact that all globally important food crops naturally form this symbiosis and the fungi help plants more efficiently obtain phosphate from the soil (Smith and Read, 2008). Stocks of phosphate fertilizer are rapidly being depleted (Gross, 2010). There is a simultaneous increase in demand for phosphate to help feed the growing population (Gilbert, 2009). These two combined factors represent a major threat to global food security; a threat that can potentially be reduced by better phosphate acquisition through the AM[F] symbiosis. The potential of AMF to contribute to improved crop yields has been known for decades [Rodriquez & Sanders 2015: 1054].
However, for the widespread adoption of AMF inoculation to be effective and safe, a better understanding is needed of ecological principles related to soil fungi. The authors note that few studies have linked crop yield increases with successful colonization by an introduced AMF, and they outline several challenges and questions that should be resolved to pursue this promising technique more broadly. For example, they ask whether introduced AMF establish well, and how they affect native AMF populations, and how genetic diversity in AMF populations variously affects different crops.
Stocks of phosphate fertilizer are rapidly being depleted (Gross, 2010). There is a simultaneous increase in demand for phosphate to help feed the growing population (Gilbert, 2009). These two combined factors represent a major threat to global food security; a threat that can potentially be reduced by better phosphate acquisition through the [arbuscular mycorrhizal fungi] symbiosis [Rodriquez & Sanders 2015: 1054].
Under a microscope, tiny tunnels can be seen in mineral particles from conifer forest soil. Scientists believe it is mycorrhizal fungi penetrating these particles by excreting organic acids in order to mine nutrients for their plant hosts. An estimated 150 meters of pores are bored by fungi per year per liter of E-horizon (layer that has been leached of mineral and/or organic content, leaving silicate) soil.
Photo credit: Jongmans 1997. “Scanning electron micrograph, showing 4–6-mm-thick hyphae entering a calcium feldspar at a granite surface near Lunsen, Sweden” [Jongman 1997].
Our analysis shows that inputs of phosphorus through arbuscular mycorrhizal symbioses substantially increase the ability of plants to grow and maintain nutritional quality, cascading through the biomass of consumers and predators in the ecosystem. Although they account for less than 1% of the total modelled biomass, the predicted nutritional benefit provided by arbuscular mycorrhizal fungi increased the biomass of macro-organisms in the Serengeti by 48%. When considering the management of biodiversity, future ecosystem models should account for the influence of arbuscular mycorrhizal fungi on all trophic levels [Stevens 2018: 536].
More than 70% of all angiosperm families form AM symbioses (Brundrett, 2009), and these symbioses are often essential for plant nutrition (Marschner & Dell, 1994). Mycorrhizal symbioses also improve plant tolerance to drought (Augé, 2001) and resistance to pathogens (Cameron, Neal, van Wees, & Ton, 2013) [Stevens 2018: 537].
Plant taxa vary in the degree to which they depend upon mycorrhizas; but in general, AM symbioses are essential for the nutrition of tropical plants, and warm season grasses are often highly dependent on mycorrhizas, acquiring up to 90% of their phosphorus requirements from AM fungi [Stevens 2018: 537].
Thirty years ago, McNaughton, Ruess, and Seagle (1988) concluded that large mammals have a major organising effect in the Serengeti ecosystem. From our analysis, we can conclude that AM fungi also play a critical role in the trophic structure of the Serengeti. Our model simulations suggest that although AM fungi account for less than 1% of the total biomass, phosphorus supplied by AM symbioses sustains half the vegetation biomass, and accordingly, half of the biomass of iconic migratory herbivores and one-third of the carnivore biomass [Stevens 2018: 542].
The distribution of soil phosphorus in the Serengeti, transported through AM symbioses and accelerated by migratory ungulates, may be a significant driver of plant diversity and ultimately mammalian carrying capacity (Anderson et al., 2007; McNaughton, Zuniga, McNaughton, & Banyikwa, 1997). Without AM fungal inputs of phosphorus, these nutrient diffusion gradients would undoubtedly decline [Stevens 2018: 543].
Two major groups of mycorrhizal fungi are arbuscular mycorrhiza (AM) and ectomycorrrhiza (EM). Both form a symbiosis with plants by colonizing their roots and creating an interface where carbon from the plant can be exchanged for phosphorus,nitrogen and other nutrients from the soil andtransferred to the plant by the fungi. The extraradical[15] mycorrhizal mycelium (ERMM), which are the vast portion of the fungal network that branches out into the soil, is difficult to study and has therefore been considered the “hidden half” of the symbiosis.
Progress in understanding the nature, extent, functioning, and identity of mycorrhizal fungal networks has been seriously hampered by the difficulties inherent in observing and studying mycelial systems without disturbing and destroying them.… As a consequence, the external mycelium, which is the fungal structure of mycorrhiza that is most intimately associated with the soil and furthest from the roots, and by implication the most critical for nutrient uptake, is normally overlooked and has been rarely recorded. Only in the past decade have studies started to focus specifically on the extent and functioning of ERMM in the field [Leake 2004: 1017].
This article highlights the significant yet overlooked role of mycorrhizal fungi in ecosystem functioning and reviews some advances in the techniques used to study these hidden powerhouses.
ERMM is the hidden power behind plant community composition and ecosystem functioning through the major processes it carries out, such as nutrient uptake, weathering of minerals, soil aggregate stability, and the way in which it alters competition between plants [Leake 2004: 1039].
The symbiosis with plants is the source of power for these fungi, given that the carbon received from plant hosts is practically inexhaustible and costs the plants little.
Despite the substantial biomass and associated C drain on their hosts, the actual “cost” of mycorrhiza to plants may be negligible because mycorrhizal colonization can increase the rate of photosynthesis (Wright et al. 1998), alleviate shoot N and P limitation, and cause a substantial increase in leaf area arising from improved nutrition (Read and Perez-Moreno 2003) [Leake 2004: 1021].
Thus,
The empowerment of mycorrhizal networks with substantial amounts of host-derived C allows them to play central roles in major biogeochemical cycles [Leake 2004: 1030].
The article concludes by emphasizing the importance for sustainable agriculture of a broader public understanding of the role of mycorrhiza for improving soil health and crop yields.
AM [arbuscular mycorrhizal] hyphal lengths in soil show strong positive correlations with soil-aggregate stability (Rillig et al. 2002; Kabir and Koide 2002), P uptake efficiency (Schweiger and Jakobsen 2000), and crop-yield improvements (Kabir and Koide 2002). Interest in the development of less intensive management systems is presenting new opportunities for adapting agricultural production systems to enhance these benefits that can be gained from AM networks. Substantial improvements in “soil health” and AM functioning in field crops are gained by the doubling of lengths of AM hyphae in soil when tillage is reduced (Kabir et al. 1998a, 1998b). Similar gains are achieved by growth of AM-compatible cover crops in place of winter fallow (Kabir and Koide 2002) [Leake 2004: 1038].
ERMM [extraradical mycorrhizal mycelium, or fungi] is the hidden power behind plant community composition and ecosystem functioning through the major processes it carries out, such as nutrient uptake, weathering of minerals, soil aggregate stability, and the way in which it alters competition between plants [Leake 2004: 1039].
Consider the fate of the approximately 17.5 million tonnes of phosphorus mined in 2005, analysed in the paper by Cordell et al. About 14 million tonnes of this were used in fertilizer (much of the rest went into cattle-feed supplements, food preservatives, and the production of detergents and industrial cleaning agents) but only about 3 million tonnes made it to the fork (or chopstick). The largest loss — around 8 million tonnes — was directly from farms through soil leaching and erosion” [Elser & Bennett 2011: 30].
To handle the twin problems of phosphorus pollution and scarcity, strategies for phosphorus conservation and recycling are urgently needed.
The solutions to these problems lie in recapturing and recycling phosphorus, moving it from where there is too much to where there is too little, and developing ways to use it more efficiently. Many strategies are simple and readily available, even for poor farmers and developing economies [Elser & Bennett 2011: 30].
The authors’ solutions include: widespread adoption of agricultural conservation practices, reduction of food waste (at least in part by producing food within or closer to cities), recycling human waste, reducing meat consumption, recovering nutrients from confined livestock facilities (such as through bioreactors), and genetically engineering plants and animals to require lower phosphorus inputs. No mention in this article of the role of plant-fungi symbiosis in accessing phosphorus in the soil.
Our review of estimates of P recycling in the human P cycle show considerable variability and uncertainty, but today it appears that only about one-quarter of the fertilizer P used in agriculture is recycled back to fields. The rest is lost to the cycle, and much of this loss ends up in waterways, causing expensive eutrophication problems. As with other nonrenewable natural resources, a sustainable P supply is not assured, and some projections show economically viable mineral reserves being depleted within decades. In addition to our review of human effects on the global P cycle, we present a number of sustainable solutions that involve closing the loop on the human P cycle. Some of these solutions are relatively straightforward but many involve overcoming considerable infrastructural or institutional inertia [Childers 2011: 123].
Economically viable mineral phosphorus reserves may become depleted within decades, threatening global crop production for a growing world population. The authors discuss this problem in relation to human P cycle, where the vast majority of mined phosphorus is not recycled back onto farm fields, but is released more or less irretrievably into the environment, polluting water bodies. “There are considerable social and environmental costs of P being lost from the currently ‘open’ human P cycle” [Childers 2011: 120].
The authors present several options for closing the human P cycle at the points of agricultural production, distribution and consumption, and human waste treatment. These options include reducing fertilizer application rates to better match plant needs, reducing erosion rates, reducing food waste, and recycling human urine, which is rich in phosphorus and nitrogen. The authors state that their list of solutions is not exhaustive, but rather is meant to stimulate others to think about the sustainability challenges of the human P cycle. Indeed, missing in this paper’s list of solutions is a discussion of the role of fungi, which can access otherwise inaccessible soil phosphorus through symbiosis with plants.
Nitrogen (N) is the most important essential element for crop production because it is required in large amounts and is nearly always the first nutrient that becomes limiting after an ecosystem is converted to cropland. Cereal grains provide about 50% of the world’s calories, and their production has become largely dependent on the use of synthetic N fertilizer. However, fertilizer N not used by plants can degrade the environment and negatively impact both people and ecosystems. In addition, efficient use of N fertilizer generally requires phosphorus (P) fertilizer which is made from rock phosphate derived from mines. Therefore, huge amounts of N and P from outside sources are being added to the environment each successive year leading to additional environmental concerns [Stewart & Lal 2017: 124A].
This article articulates a presumed “nitrogen dilemma,” as described above, that, on the one hand, agriculture requires increasing amounts of nitrogen and phosphorus fertilizer, especially as the population surges toward 10 billion. On the other hand, ongoing fertilizer application will lead to increasingly polluted and impaired fresh waters around the world, increased greenhouse gas emissions, and over-reliance on limited supplies of mined phosphorus.
The difficulty of reducing nitrogen inputs is twofold according to the article: First, farmers cannot know exactly how much nitrogen their crops will need because yield depends on water supply and respiration rates, and only indirectly on nitrogen availability. Therefore, farmers are reluctant to limit fertilizer input for fear it could in turn limit water utilization. “Because N is usually the first limiting factor other than water, most farmers want to make sure they have enough N available to fully utilize the water” [p.126A]. Second, nutrient-polluted waters is a local problem, and therefore most likely requires a local political solution, rather than being manageable through national regulations. Local policy solutions will happen only when enough people feel the direct effects of the problem locally and demand action.
Curiously, the article fails to mention the farming practices that reduce the need for fertilizers, maximize the soil’s water-holding capacity, and cool the soil through continuous vegetative cover. Practices designed to enhance soil organic matter, such as cover-cropping and replacing synthetic fertilizer input with compost, manure and crop residues, can achieve the same goals that nitrogen fertilizer is supposed to address.
…it is water that determines yield, and the amount of water available for a crop is beyond control of the farmer, even if the crop is irrigated. This is because it is only the amount of water transpired by the growing crop that determines the amount of biomass produced by photosynthesis, and this is affected not only by the amount of water available but on other climatic factors such as temperature, radiation, humidity, and wind [Steward & Lal 2017: 126A].
We argue that there is no nitrogen dilemma unless we cling to the idea that industrial agriculture is the only way forward despite its increasingly apparent fragility, while rejecting the potential of multifunctional, regenerative agriculture to broadly achieve our production and environmental goals.
The nitrogen dilemma: food or the environment, Stewart & Lal 2017
Nitrogen (N) is the most important essential element for crop production because it is required in large amounts and is nearly always the first nutrient that becomes limiting after an ecosystem is converted to cropland. Cereal grains provide about 50% of the world’s calories, and their production has become largely dependent on the use of synthetic N fertilizer. However, fertilizer N not used by plants can degrade the environment and negatively impact both people and ecosystems. In addition, efficient use of N fertilizer generally requires phosphorus (P) fertilizer which is made from rock phosphate derived from mines. Therefore, huge amounts of N and P from outside sources are being added to the environment each successive year leading to additional environmental concerns [Stewart & Lal 2017: 124A].
This article articulates a presumed “nitrogen dilemma,” as described above, that, on the one hand, agriculture requires increasing amounts of nitrogen and phosphorus fertilizer, especially as the population surges toward 10 billion. On the other hand, ongoing fertilizer application will lead to increasingly polluted and impaired fresh waters around the world, increased greenhouse gas emissions, and over-reliance on limited supplies of mined phosphorus.
The difficulty of reducing nitrogen inputs is twofold according to the article: First, farmers cannot know exactly how much nitrogen their crops will need because yield depends on water supply and respiration rates, and only indirectly on nitrogen availability. Therefore, farmers are reluctant to limit fertilizer input for fear it could in turn limit water utilization. “Because N is usually the first limiting factor other than water, most farmers want to make sure they have enough N available to fully utilize the water” [p.126A]. Second, nutrient-polluted waters is a local problem, and therefore most likely requires a local political solution, rather than being manageable through national regulations. Local policy solutions will happen only when enough people feel the direct effects of the problem locally and demand action.
Curiously, the article fails to mention the farming practices that reduce the need for fertilizers, maximize the soil’s water-holding capacity, and cool the soil through continuous vegetative cover. Practices designed to enhance soil organic matter, such as cover-cropping and replacing synthetic fertilizer input with compost, manure and crop residues, can achieve the same goals that nitrogen fertilizer is supposed to address.
…it is water that determines yield, and the amount of water available for a crop is beyond control of the farmer, even if the crop is irrigated. This is because it is only the amount of water transpired by the growing crop that determines the amount of biomass produced by photosynthesis, and this is affected not only by the amount of water available but on other climatic factors such as temperature, radiation, humidity, and wind [Steward & Lal 2017: 126A].
We argue that there is no nitrogen dilemma unless we cling to the idea that industrial agriculture is the only way forward despite its increasingly apparent fragility, while rejecting the potential of multifunctional, regenerative agriculture to broadly achieve our production and environmental goals.
Sustainability challenges of phosphorus and food: solutions from closing the human phosphorus cycle, Childers 2011
Our review of estimates of P recycling in the human P cycle show considerable variability and uncertainty, but today it appears that only about one-quarter of the fertilizer P used in agriculture is recycled back to fields. The rest is lost to the cycle, and much of this loss ends up in waterways, causing expensive eutrophication problems. As with other nonrenewable natural resources, a sustainable P supply is not assured, and some projections show economically viable mineral reserves being depleted within decades. In addition to our review of human effects on the global P cycle, we present a number of sustainable solutions that involve closing the loop on the human P cycle. Some of these solutions are relatively straightforward but many involve overcoming considerable infrastructural or institutional inertia [Childers 2011: 123].
Economically viable mineral phosphorus reserves may become depleted within decades, threatening global crop production for a growing world population. The authors discuss this problem in relation to human P cycle, where the vast majority of mined phosphorus is not recycled back onto farm fields, but is released more or less irretrievably into the environment, polluting water bodies. “There are considerable social and environmental costs of P being lost from the currently ‘open’ human P cycle” [Childers 2011: 120].
The authors present several options for closing the human P cycle at the points of agricultural production, distribution and consumption, and human waste treatment. These options include reducing fertilizer application rates to better match plant needs, reducing erosion rates, reducing food waste, and recycling human urine, which is rich in phosphorus and nitrogen. The authors state that their list of solutions is not exhaustive, but rather is meant to stimulate others to think about the sustainability challenges of the human P cycle. Indeed, missing in this paper’s list of solutions is a discussion of the role of fungi, which can access otherwise inaccessible soil phosphorus through symbiosis with plants.
A broken biogeochemical cycle, Elser & Bennett 2011
Consider the fate of the approximately 17.5 million tonnes of phosphorus mined in 2005, analysed in the paper by Cordell et al. About 14 million tonnes of this were used in fertilizer (much of the rest went into cattle-feed supplements, food preservatives, and the production of detergents and industrial cleaning agents) but only about 3 million tonnes made it to the fork (or chopstick). The largest loss — around 8 million tonnes — was directly from farms through soil leaching and erosion” [Elser & Bennett 2011: 30].
To handle the twin problems of phosphorus pollution and scarcity, strategies for phosphorus conservation and recycling are urgently needed.
The solutions to these problems lie in recapturing and recycling phosphorus, moving it from where there is too much to where there is too little, and developing ways to use it more efficiently. Many strategies are simple and readily available, even for poor farmers and developing economies [Elser & Bennett 2011: 30].
The authors’ solutions include: widespread adoption of agricultural conservation practices, reduction of food waste (at least in part by producing food within or closer to cities), recycling human waste, reducing meat consumption, recovering nutrients from confined livestock facilities (such as through bioreactors), and genetically engineering plants and animals to require lower phosphorus inputs. No mention in this article of the role of plant-fungi symbiosis in accessing phosphorus in the soil.
Networks of power and influence: the role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning, Leake 2004
Two major groups of mycorrhizal fungi are arbuscular mycorrhiza (AM) and ectomycorrrhiza (EM). Both form a symbiosis with plants by colonizing their roots and creating an interface where carbon from the plant can be exchanged for phosphorus,nitrogen and other nutrients from the soil andtransferred to the plant by the fungi. The extraradical[15] mycorrhizal mycelium (ERMM), which are the vast portion of the fungal network that branches out into the soil, is difficult to study and has therefore been considered the “hidden half” of the symbiosis.
Progress in understanding the nature, extent, functioning, and identity of mycorrhizal fungal networks has been seriously hampered by the difficulties inherent in observing and studying mycelial systems without disturbing and destroying them.… As a consequence, the external mycelium, which is the fungal structure of mycorrhiza that is most intimately associated with the soil and furthest from the roots, and by implication the most critical for nutrient uptake, is normally overlooked and has been rarely recorded. Only in the past decade have studies started to focus specifically on the extent and functioning of ERMM in the field [Leake 2004: 1017].
This article highlights the significant yet overlooked role of mycorrhizal fungi in ecosystem functioning and reviews some advances in the techniques used to study these hidden powerhouses.
ERMM is the hidden power behind plant community composition and ecosystem functioning through the major processes it carries out, such as nutrient uptake, weathering of minerals, soil aggregate stability, and the way in which it alters competition between plants [Leake 2004: 1039].
The symbiosis with plants is the source of power for these fungi, given that the carbon received from plant hosts is practically inexhaustible and costs the plants little.
Despite the substantial biomass and associated C drain on their hosts, the actual “cost” of mycorrhiza to plants may be negligible because mycorrhizal colonization can increase the rate of photosynthesis (Wright et al. 1998), alleviate shoot N and P limitation, and cause a substantial increase in leaf area arising from improved nutrition (Read and Perez-Moreno 2003) [Leake 2004: 1021].
Thus,
The empowerment of mycorrhizal networks with substantial amounts of host-derived C allows them to play central roles in major biogeochemical cycles [Leake 2004: 1030].
The article concludes by emphasizing the importance for sustainable agriculture of a broader public understanding of the role of mycorrhiza for improving soil health and crop yields.
AM [arbuscular mycorrhizal] hyphal lengths in soil show strong positive correlations with soil-aggregate stability (Rillig et al. 2002; Kabir and Koide 2002), P uptake efficiency (Schweiger and Jakobsen 2000), and crop-yield improvements (Kabir and Koide 2002). Interest in the development of less intensive management systems is presenting new opportunities for adapting agricultural production systems to enhance these benefits that can be gained from AM networks. Substantial improvements in “soil health” and AM functioning in field crops are gained by the doubling of lengths of AM hyphae in soil when tillage is reduced (Kabir et al. 1998a, 1998b). Similar gains are achieved by growth of AM-compatible cover crops in place of winter fallow (Kabir and Koide 2002) [Leake 2004: 1038].
ERMM [extraradical mycorrhizal mycelium, or fungi] is the hidden power behind plant community composition and ecosystem functioning through the major processes it carries out, such as nutrient uptake, weathering of minerals, soil aggregate stability, and the way in which it alters competition between plants [Leake 2004: 1039].
Mycorrhizal symbioses influence the trophic structure of the Serengeti, Stevens 2018
Our analysis shows that inputs of phosphorus through arbuscular mycorrhizal symbioses substantially increase the ability of plants to grow and maintain nutritional quality, cascading through the biomass of consumers and predators in the ecosystem. Although they account for less than 1% of the total modelled biomass, the predicted nutritional benefit provided by arbuscular mycorrhizal fungi increased the biomass of macro-organisms in the Serengeti by 48%. When considering the management of biodiversity, future ecosystem models should account for the influence of arbuscular mycorrhizal fungi on all trophic levels [Stevens 2018: 536].
More than 70% of all angiosperm families form AM symbioses (Brundrett, 2009), and these symbioses are often essential for plant nutrition (Marschner & Dell, 1994). Mycorrhizal symbioses also improve plant tolerance to drought (Augé, 2001) and resistance to pathogens (Cameron, Neal, van Wees, & Ton, 2013) [Stevens 2018: 537].
Plant taxa vary in the degree to which they depend upon mycorrhizas; but in general, AM symbioses are essential for the nutrition of tropical plants, and warm season grasses are often highly dependent on mycorrhizas, acquiring up to 90% of their phosphorus requirements from AM fungi [Stevens 2018: 537].
Thirty years ago, McNaughton, Ruess, and Seagle (1988) concluded that large mammals have a major organising effect in the Serengeti ecosystem. From our analysis, we can conclude that AM fungi also play a critical role in the trophic structure of the Serengeti. Our model simulations suggest that although AM fungi account for less than 1% of the total biomass, phosphorus supplied by AM symbioses sustains half the vegetation biomass, and accordingly, half of the biomass of iconic migratory herbivores and one-third of the carnivore biomass [Stevens 2018: 542].
The distribution of soil phosphorus in the Serengeti, transported through AM symbioses and accelerated by migratory ungulates, may be a significant driver of plant diversity and ultimately mammalian carrying capacity (Anderson et al., 2007; McNaughton, Zuniga, McNaughton, & Banyikwa, 1997). Without AM fungal inputs of phosphorus, these nutrient diffusion gradients would undoubtedly decline [Stevens 2018: 543].
Rock-eating fungi, Jongmans 1997
Under a microscope, tiny tunnels can be seen in mineral particles from conifer forest soil. Scientists believe it is mycorrhizal fungi penetrating these particles by excreting organic acids in order to mine nutrients for their plant hosts. An estimated 150 meters of pores are bored by fungi per year per liter of E-horizon (layer that has been leached of mineral and/or organic content, leaving silicate) soil.
Photo credit: Jongmans 1997. “Scanning electron micrograph, showing 4–6-mm-thick hyphae entering a calcium feldspar at a granite surface near Lunsen, Sweden” [Jongman 1997].
The role of community and population ecology in applying mycorrhizal fungi for improved food security, Rodriguez & Sanders 2015
Given that nitrogen and phosphorus are the most limiting nutrients for crop growth, that global phosphorus supplies are becoming exhausted while the human population rapidly expands, and that arbuscular mycorrhizal fungi (AMF) symbioses improve crop phosphorus acquisition, AMF symbioses have a major role to play in current and future crop production.
The potential of AMF to help increase global food security lies in the fact that all globally important food crops naturally form this symbiosis and the fungi help plants more efficiently obtain phosphate from the soil (Smith and Read, 2008). Stocks of phosphate fertilizer are rapidly being depleted (Gross, 2010). There is a simultaneous increase in demand for phosphate to help feed the growing population (Gilbert, 2009). These two combined factors represent a major threat to global food security; a threat that can potentially be reduced by better phosphate acquisition through the AM[F] symbiosis. The potential of AMF to contribute to improved crop yields has been known for decades [Rodriquez & Sanders 2015: 1054].
However, for the widespread adoption of AMF inoculation to be effective and safe, a better understanding is needed of ecological principles related to soil fungi. The authors note that few studies have linked crop yield increases with successful colonization by an introduced AMF, and they outline several challenges and questions that should be resolved to pursue this promising technique more broadly. For example, they ask whether introduced AMF establish well, and how they affect native AMF populations, and how genetic diversity in AMF populations variously affects different crops.
Stocks of phosphate fertilizer are rapidly being depleted (Gross, 2010). There is a simultaneous increase in demand for phosphate to help feed the growing population (Gilbert, 2009). These two combined factors represent a major threat to global food security; a threat that can potentially be reduced by better phosphate acquisition through the [arbuscular mycorrhizal fungi] symbiosis [Rodriquez & Sanders 2015: 1054].
Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils, Rashid 2016
The paper argues for the use of bacterial and fungal inoculants in combination with organic amendments and cover crops to regenerate degraded soils. In order to produce enough food for a growing global population on ubiquitously degraded soils, synthetic fertilizers will be in increasingly high demand. However, these fertilizers require copious amounts of non-renewable energy to manufacture, and become pollutants when used. Here, the authors explain how bacteria and fungi make nutrients available to plants and how facilitate soil aggregation.
Meta-analysis of biofertilizer application in agriculture, Schutz 2018
Given the global decline of reserves of both rock phosphate and fossil fuel, this study poses the question – to what extent can microbial inoculants replace/reduce the use of synthetic fertilizer? The authors find that “dryland agriculture can benefit most from biofertilizers [microbial inoculants used as fertilizers]. Due to climate change, in the future there will be even more dryland areas globally. Biofertilizers are thus a promising option for sustainable agriculture” [Schutz 2018: 11]. More specifically:
Our comprehensive meta-analysis with studies from all over the world revealed that biofertilizers were found to be most effective in dry climates. Biofertilizer also improved PUE [phosphorus use efficiency] and NUE [nitrogen use efficiency] greatly. Furthermore, we found that biofertilizers possessing both N fixing and P solubilizing traits have the highest potential to improve the crop yields. Interestingly, AMFs, known for facilitating P nutrient uptake in plants, were on par with applications of biofertilizers with the combined traits of N fixation and P solubilization, indicating the big potential of AMFs as sole biofertilizer for most crops and climatic situations [Schutz 2018: 5].
Future Directions International Strategic Directions Paper: Agricultural Application of Mycorrhizal Fungi to Increase Crop Yields, Promote Soil Health and Combat Climate Change, Johns 2014
There are a number of agricultural practices that will enhance fungi colonisation. Wherever possible, of course, the full range of critical soil health processes that govern productivity should be allowed to regenerate agricultural ecologies naturally. It may, however, be necessary or more practical to inoculate seed with fungi spores in order to recover degraded soils. A number of farmers in the Great Southern agricultural region of Western Australia are undertaking this course of action. Finding themselves confronted with an unsustainable spiral of ever-increasing commercial fertiliser costs and uneconomic or diminishing crop yields, it was realised that a different approach needed to be taken. In recent growing seasons, seed has been inoculated with commercial fungi spores just prior to planting. While it is still too early to provide statistically robust outcomes and, bearing in mind that there are no “silver bullets” in agricultural production, the indications are that mycorrhizal fungi are promoting improvements in crop vitality, yield and soil condition [Johns 2014: 4].
Nitrogen: the double-edged sword, Jones 2014
The symbiosis between mycorrhizal fungi and plants drive carbon and nitrogen cycles. Fungi demand carbon exudate from plants in exchange for nitrogen and other nutrients retrieved and transported from the soil. The “liquid carbon” exuded from plant roots feeds mycorrhizal fungi and many other soil microbes, while also becoming stabilized in soil aggregates and humus. Jones explains that when this mycorrhizal exchange is inhibited by N fertilizer, which allows plants to absorb nitrogen “for free” (without providing liquid carbon in exchange), this reduces the flow of carbon into the soils, which in turn diminishes fungal networks and their delivery of micronutrients to plant hosts, and results in carbon-depleted soils.
Despite its abundance in the atmosphere, nitrogen is frequently the most limiting element for plants. There is a reason for this. Carbon, essential to photosynthesis and soil function, occurs as a trace gas, carbon dioxide, currently comprising 0.04% of the atmosphere. The most efficient way to transform CO2 to stable organic soil complexes (containing both C and N) is via the liquid carbon pathway. The requirement for biologically-fixed nitrogen drives this process.
If plants were able to access nitrogen directly from the atmosphere, their growth would be impeded by the absence of carbon-rich topsoil. We are witnessing an analogous situation in agriculture today. When inorganic nitrogen is provided, the supply of carbon to associative nitrogen fixing microbes is inhibited, resulting in carbon-depleted soils.
Reduced carbon flows impact a vast network of microbial communities, restricting the availability of essential minerals, trace elements, vitamins and hormones required for plant tolerance to environmental stresses such as frost and drought and resistance to insects and disease. Lowered micronutrient densities in plants also translate to reduced nutritional value of food [Jones 2014: 2-3].
Jones further explains how to modify agricultural practices to protect and build the soil: maintain year-round living ground cover, limit nitrogen and phosphorus fertilizer input, promote plant and microbial diversity, and integrate livestock into crop production systems.
Synthetic nitrogen fertilizers deplete soil nitrogen: a global dilemma for sustainable cereal production, Mulvaney 2009
There is a prevailing view that global food and fiber production will continue to expand because of modern agricultural management systems with improved cultivars and intensive chemical inputs dominated by synthetic ammoniacal fertilizers. The use of these fertilizers has led to concerns regarding water and air pollution but is generally perceived to play an essential role for sustaining agricultural productivity, not only by supplying the most important nutrient for cereal production but also by increasing the input of crop residues for building soil organic matter. The scientific soundness of the buildup concept has yet to be substantiated empirically using baseline data sets from long-term cropping experiments. The present paper and a companion study by Khan et al. (2007) provide many such data sets that encompass a variety of cereal cropping and management systems in different parts of the world. Overwhelmingly, the evidence is diametrically opposed to the buildup concept and instead corroborates a view elaborated long ago by White (1927) and Albrecht (1938) that fertilizer N depletes soil organic matter by promoting microbial C utilization and N mineralization. An inexorable conclusion can be drawn: The scientific basis for input-intensive cereal production is seriously flawed. The long-term consequences of continued reliance on current production practices will be a decline in soil productivity that increases the need for synthetic N fertilization, threatens food security, and exacerbates environmental degradation [Mulvaney 2009: 2308].
Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere, Paungfoo-Lonhienne 2015
In this study, nitrogen fertilization altered the relative abundance of fungal taxa in the rhizosphere, increasing fungal genera with known pathogenic traits, and decreasing a fungal phyla (Basidiomycetes) known to break down lignin, thus important for carbon cycling in the soil.
Fungi play important roles as decomposers, plant symbionts and pathogens in soils. The structure of fungal communities in the rhizosphere is the result of complex interactions among selection factors that may favour beneficial or detrimental relationships. Using culture-independent fungal community profiling, we have investigated the effects of nitrogen fertilizer dosage on fungal communities in soil and rhizosphere of field-grown sugarcane.The results show that the concentration of nitrogen fertilizer strongly modifies the composition but not the taxon richness of fungal communities in soil and rhizosphere. Increased nitrogen fertilizer dosage has a potential negative impact on carbon cycling in soil and promotes fungal genera with known pathogenic traits, uncovering a negative effect of intensive fertilization [Paungfoo-Lonhienne 2015:just 1].
Nitrogen fertilizer effects on soil carbon balances in Midwestern U.S. agricultural systems, Russell 2009
Despite increasing residue input in annual crop production systems, N fertilization does not increase soil organic carbon (SOC) over time because N fertilization also increases organic carbon (OC) decay. This study also shows that belowground OC inputs contribute to soil carbon sequestration more than aboveground OC inputs to the soil.
When all phases of the crop rotations were evaluated over the long term, OC decay rates increased concomitantly with OC input rates in several systems. Increases in decay rates with N fertilization apparently offset gains in carbon inputs to the soil in such a way that soil C sequestration was virtually nil in 78% of the systems studied, despite up to 48 years of N additions [Russell 2009: 1102].
Across all systems, SOC storage was significantly correlated with the quantity of belowground OM [organic matter] inputs (P < 0.01, both sites). In contrast, SOC was not correlated with the quantity of aboveground inputs (P = 0.45, Nashua; P = 0.55, Kanawha) [Russell 2009: 1111].
This study highlights the importance of incorporating both production and decomposition processes, as well as the location (above- or below-ground) of detrital inputs into models of N-fertilization effects on soil C dynamics in agroecosystems. These results are highly relevant for evaluating the potential of N fertilization to mitigate the effects of removal of organic-matter ‘‘residue’’ from the system for bioenergy production. Our data suggest that the stimulation of OC decomposition by the addition of fertilizer N would likely counteract the positive effects of N fertilization on inputs of OC to the soil, at least for annual crops. Given the current quantity of N that is applied over such a large area, management strategies that can maintain high yields and also reduce N-fertilizer use would also have beneficial environmental consequences. Our study indicates that selection of crops for higher belowground NPP [net primary production], in rotation with crops that fix N, could maximize both yields and soil C sequestration without excessive N-fertilizer additions [Russell 2009: 1111].
Community proteogenomics reveals the systemic impact of phosphorus availability on microbial functions in tropical soil, Yao 2018
In this study, long-term phosphorus fertilization limited the extent to which the genes and proteins of microbial communities were allocated to degrading recalcitrant soil phytate to acquire phosphorus. In phosphorus-deficient soil, on the other hand, the genes responsible for breaking down recalcitrant substrate to acquire phosphorus were more prevalent in microbial communities. In other words, microbial communities can adapt genetically to different levels of nutrients in the soil in order to continue meeting their nutritional requirements. This adds to the body of evidence that fertilizer use impairs the inherent qualities of a living soil to nourish the plants growing there.
A greater than fourfold increase in the gene abundance of 3-phytase was the strongest response of soil communities to phosphorus deficiency. Phytase catalyses the release of phosphate from phytate, the most recalcitrant phosphorus-containing compound in soil organic matter. Genes and proteins for the degradation of phosphorus-containing nucleic acids and phospholipids, as well as the decomposition of labile carbon and nitrogen, were also enhanced in the phosphorus-deficient soils. In contrast, microbial communities in the phosphorus-rich soils showed increased gene abundances for the degradation of recalcitrant aromatic compounds, transformation of nitrogenous compounds and assimilation of sulfur. Overall, these results demonstrate the adaptive allocation of genes and proteins in soil microbial communities in response to shifting nutrient constraints [Yao 2018: 499].
In conclusion, our proteogenomics results provide systems biology insights into the adaptation of soil microbial communities to different levels of phosphorus availability in a humid tropical forest environment. Phosphorus deficiency significantly enhanced the genetic capabilities of microbial communities to extract phosphorus from phytate and, to a lesser extent, from nucleic acids and phospholipids. Long-term phosphorus fertilization altered the allocation of genes and proteins by microbial communities to acquire carbon, nitrogen and sulfur from a variety of substrates. The results suggest that the selective degradation of recalcitrant substrates, including phytate in phosphorus-deficient soils and aromatic compounds in phosphorus-rich soils, is an important means for microbial communities to balance their elemental requirements. The adaptive allocation of genes and proteins for acquisition of these nutrients in different soils can be explained as an optimal foraging strategy by which microbial communities maintain efficient growth under resource limitation [Yao 2018: 505].
Agrochemical companies argue that crops can’t be grown without their products. And in a sense, they are right, as long as we accept as inevitable a dysfunctional soil food web [LSP 2018: 16].
The importance of synthetic fertilizer for global crop production and the environmental consequences of its excessive use is often presented as a dilemma [Steward & Lal 2017, Mulvaney 2009].
Indeed, many problems arise from our dependence on fertilizers, including the energy-intensiveness of nitrogen fertilizer production, the increasing scarcity of global phosphorus reserves, and the leaching of both nutrients from farm fields, polluting surface waters.
To remedy this, some advocate for a more judicious use of fertilizers and better ways to recapture and recycle it. Only a quarter of mined phosphorus is recycled back on to cropland [Childers 2011] while the rest is lost, becoming a pollutant. Even phosphorus that has made it into our bodies as food eventually becomes human waste, which could be though is often not recaptured and recycled.
While the idea of recycling phosphorus is relevant and timely, it presumes the continuation of conventional high-input agriculture. It assumes ongoing dependence on synthetic fertilizers. It presumes this ‘dilemma.’
Yet there may be a simpler, more elegant solution. Soil microbes provide benefits comparable to chemical inputs in terms of crop yield, but without the negative side-effects. Indeed,healthy soil is teeming with diversity, where billions of mostly microscopic “willing workers” in microbiologist Elaine Ingham’s words [LSP 2018: 16] make nutrients available to plants in the process of breaking down organic material and mining soil particles.
In a sample of conifer forest soil, for example, tiny tunnels can be seen in mineral particles [Jongmans 1997]. Scientists believe that mycorrhizal fungi penetrate these particles by excreting organic acids in order to mine nutrients for their plant hosts. An estimated 150 meters of pores are bored by fungi into grains of feldspar sand per year per liter of soil.
Dr. David Johnson of New Mexico State University has found that the most productive plants are not those grown with fertilizer, nor even with the most organic matter per se [Johnson 2017]. Rather, plant productivity stems from the robustness of the soil microbial community. He discovered this in an experiment that compared the growth rate of chili peppers in different soils, including a fungi-rich compost on one hand and a bacterial-dominant soil typical of most croplands on the other.
Johnson [2017] found that only 3% of carbon flow went into plant biomass production when the soil’s fungi to bacteria (F:B) ratio was low (0.04). The remainder of the carbon produced by these plants was going into other functions, including nitrogen fixation, exudates to the soil, and respiration. At a higher F:B ratio (3.68), by contrast, plant growth was more efficient with 56% of carbon flow going to the development of the plants’ roots, stems, leaves and fruit, resulting in bigger plants. Similarly, in a cover crop experiment where a desert soil was inoculated with a robust compost-derived microbial community, Johnson produced biomass comparable to the most productive (tropical rainforest) ecosystems on Earth.
“Diversity is the currency of survival, and that’s what’s making this system work so well” [Johnson 2017: 28:08 min]. Johnson explains that the key to plant productivity is microbial diversity, where multiple populations of organisms are serving vital ecosystem functions, including fixing nitrogen, solubilizing phosphorus, and secreting plant growth hormones, for example. Mycorrhizal fungi, in particular, play a key role in connecting plants to the soil ecosystems that nourish them. These fungi colonize the surface of plant roots and branch out into the soil, effectively extending the roots further to collect nutrients otherwise out of reach. In short, fertilizer inputs are not needed when microorganisms in the soil are there to pull the requisite nitrogen from the air and minerals from the ground on behalf of plants.
In return for their services, bacteria and fungi are nourished by carbon from plant litter and root exudates. For the whole system to function, Johnson explains, a constant input of energy is needed in the form of carbon compounds manufactured by plants through photosynthesis. Therefore, bare fallow fields are deadly for the soil ecosystem, and in turn less hospitable to crops later grown there.
Taking an ecosystem-wide view, Stevens [2018] found that arbuscular mycorrhizal fungi, though “they account for less than 1% of the total modelled biomass … increased the biomass of macro-organisms in the Serengeti by 48%.” In other words, absent fungi, plants would be only half as productive, resulting in less food for herbivores, and half the biomass growth all the way up the food chain. While plants differ in their relative dependence on fungi, warm season grasses derive as much as 90% of their phosphorus from mycorrhizal symbioses [Stevens 2018: 537].
We don’t fertilize nature and yet it can achieve some triple the productivity of the world’s best crop plots [Johnson 2017]. Why, then, do we rely almost exclusively on fertilizers, and why have most of us never heard about the power of soil microorganisms for improving crop productivity? In part, our ignorance stems from the difficulty of studying the soil and its microscopic inhabitants.
Progress in understanding the nature, extent, functioning, and identity of mycorrhizal fungal networks has been seriously hampered by the difficulties inherent in observing and studying mycelial systems without disturbing and destroying them [Leake 2004: 1017].
Further blocking our collective awareness of soil microbes’ role in plant productivity is their erasure by tillage, fertilizer and pesticides. There is a physical erasure in terms of the damage these practices do to the soil ecosystem. And there’s a cognitive erasure in terms of our general acceptance that yield goals are attainable only with chemical inputs.
Ironically, at the same time that chemical inputs and tillage replace soil microbes’ work by supplying nutrients to plants and defending against pests, these practices also disrupt the soil microbial community. Tillage disturbs mycorrhizal fungi by breaking their hyphal networks. Fertilizer application disrupts the exchange between plants and microorganisms. When plants can absorb nitrogen “for free” (without providing carbon in exchange), explains soil ecologist Christine Jones [2014], this reduces the flow of carbon into the soil resulting in carbon-depleted soils and diminishing fungal networks and their delivery of micronutrients to plant hosts.
Reduced carbon flows impact a vast network of microbial communities, restricting the availability of essential minerals, trace elements, vitamins and hormones required for plant tolerance to environmental stresses such as frost and drought and resistance to insects and disease. Lowered micronutrient densities in plants also translate to reduced nutritional value of food [Jones 2014: 2-3].
Fertilizer application has also been shown to lower fungal diversity [Zhao 2016] and to favor fungal genera with known pathogenic traits [Paungfoo-Lonhienne 2015]. Furthermore, chronic fertilizer use diminishes soil fertility [Russell 2009, Khan 2007, Clemmenson 2013, Shahbaz 2016, Mulvaney 2009]. This finding contradicts the commonly held view that fertilizer use over time builds up soil fertility by increasing plant biomass, and thus plant residue input to the soil. In fact, fertilizer speeds up the breakdown and loss of soil organic carbon and soil nitrogen.
Overwhelmingly, the evidence is diametrically opposed to the buildup concept and instead corroborates a view elaborated long ago by White (1927) and Albrecht (1938) that fertilizer N depletes soil organic matter by promoting microbial C utilization and N mineralization. An inexorable conclusion can be drawn: The scientific basis for input-intensive cereal production is seriously flawed. The long-term consequences of continued reliance on current production practices will be a decline in soil productivity that increases the need for synthetic N fertilization, threatens food security, and exacerbates environmental degradation [Mulvaney 2009: 2308].
An inexorable conclusion can be drawn: The scientific basis for input-intensive cereal production is seriously flawed. The long-term consequences of continued reliance on current production practices will be a decline in soil productivity that increases the need for synthetic N fertilization, threatens food security, and exacerbates environmental degradation [Mulvaney 2009: 2308].
Indeed, due to carbon-diminishing management practices, agricultural soils contain 25% to 75% less SOC than soils in undisturbed, natural ecosystems [Lal 2010]. Consequently, scientists and farmers alike are beginning to look beyond fertilizer for a solution to poorly functioning soils. For instance, in hot, dry Western Australia farmers are experimenting with inoculating seeds with beneficial fungi.
Finding themselves confronted with an unsustainable spiral of ever-increasing commercial fertiliser costs and uneconomic or diminishing crop yields, it was realised that a different approach needed to be taken. In recent growing seasons, seed has been inoculated with commercial fungi spores just prior to planting. While it is still too early to provide statistically robust outcomes and, bearing in mind that there are no “silver bullets” in agricultural production, the indications are that mycorrhizal fungi is promoting improvements in crop vitality, yield and soil condition [Johns 2014].
A recent meta-analysis [Schutz 2018] suggests that these Australian farmers are on the right track. Researchers analyzed a couple hundred studies of various microbial inoculants used as “biofertilizers,” grouping them by their functional traits: nitrogen fixation, solubilizing phosphorus, or mycorrhizal fungi. Corroborating David Johnson’s findings, they concluded that microbial inoculants, especially mycorrhizal fungi, are a promising option for sustainable agriculture, especially in dry climates.
Christine Jones [2014] expands on this idea, saying that in addition to promoting plant and microbial diversity, farmers wanting to build soil health should maintain year-round living ground cover, limit nitrogen and phosphorus fertilizer input, and integrate livestock into crop production systems. And here we have the core practices of agroecology, variously referred to as regenerative, organic, “biologique” in French, or sustainable agriculture – each name emphasizing a different aspect of a shared philosophy.
In chemistry, the word “organic” refers to almost any molecule that contains carbon. Carbon is made available to the biosphere primarily by plants through photosynthesis. Carbon is the basis for all living tissue and is thus also present in the remnants of dead organisms, otherwise known as organic material, which is plentiful in healthy soil. “Biologique” suggests the favoring of biological processes and symbioses to support plant growth. “Agroecological” emphasizes the idea that all species (including crops) present in an ecosystem rely on one another for food, shelter and immune defense, and cannot be isolated without harm being done (such as pest outbreaks and nutrient deficiencies). “Sustainable,” of course, means that the crops we grow today will not diminish the land’s ability to grow as many or more crops again tomorrow.
All these labels make reference to the importance of a living soil. Anyone who marvels at the wonder of life writ large is already at least halfway to the point of accepting that the same magic buzzes underground even though we can’t see it. Billions of microscopic “willing workers” are getting the job done – helping plants grow and thus supporting everything else up the food chain. Up here at the top of the food chain, it’s our job is to complete the circle and support those guys at the bottom.
Fertilizer vs. Fungi Article Summaries
The nitrogen dilemma: food or the environment, Stewart & Lal 2017
Nitrogen (N) is the most important essential element for crop production because it is required in large amounts and is nearly always the first nutrient that becomes limiting after an ecosystem is converted to cropland. Cereal grains provide about 50% of the world’s calories, and their production has become largely dependent on the use of synthetic N fertilizer. However, fertilizer N not used by plants can degrade the environment and negatively impact both people and ecosystems. In addition, efficient use of N fertilizer generally requires phosphorus (P) fertilizer which is made from rock phosphate derived from mines. Therefore, huge amounts of N and P from outside sources are being added to the environment each successive year leading to additional environmental concerns [Stewart & Lal 2017: 124A].
This article articulates a presumed “nitrogen dilemma,” as described above, that, on the one hand, agriculture requires increasing amounts of nitrogen and phosphorus fertilizer, especially as the population surges toward 10 billion. On the other hand, ongoing fertilizer application will lead to increasingly polluted and impaired fresh waters around the world, increased greenhouse gas emissions, and over-reliance on limited supplies of mined phosphorus.
The difficulty of reducing nitrogen inputs is twofold according to the article: First, farmers cannot know exactly how much nitrogen their crops will need because yield depends on water supply and respiration rates, and only indirectly on nitrogen availability. Therefore, farmers are reluctant to limit fertilizer input for fear it could in turn limit water utilization. “Because N is usually the first limiting factor other than water, most farmers want to make sure they have enough N available to fully utilize the water” [p.126A]. Second, nutrient-polluted waters is a local problem, and therefore most likely requires a local political solution, rather than being manageable through national regulations. Local policy solutions will happen only when enough people feel the direct effects of the problem locally and demand action.
Curiously, the article fails to mention the farming practices that reduce the need for fertilizers, maximize the soil’s water-holding capacity, and cool the soil through continuous vegetative cover. Practices designed to enhance soil organic matter, such as cover-cropping and replacing synthetic fertilizer input with compost, manure and crop residues, can achieve the same goals that nitrogen fertilizer is supposed to address.
…it is water that determines yield, and the amount of water available for a crop is beyond control of the farmer, even if the crop is irrigated. This is because it is only the amount of water transpired by the growing crop that determines the amount of biomass produced by photosynthesis, and this is affected not only by the amount of water available but on other climatic factors such as temperature, radiation, humidity, and wind [Steward & Lal 2017: 126A].
We argue that there is no nitrogen dilemma unless we cling to the idea that industrial agriculture is the only way forward despite its increasingly apparent fragility, while rejecting the potential of multifunctional, regenerative agriculture to broadly achieve our production and environmental goals.
Sustainability challenges of phosphorus and food: solutions from closing the human phosphorus cycle, Childers 2011
Our review of estimates of P recycling in the human P cycle show considerable variability and uncertainty, but today it appears that only about one-quarter of the fertilizer P used in agriculture is recycled back to fields. The rest is lost to the cycle, and much of this loss ends up in waterways, causing expensive eutrophication problems. As with other nonrenewable natural resources, a sustainable P supply is not assured, and some projections show economically viable mineral reserves being depleted within decades. In addition to our review of human effects on the global P cycle, we present a number of sustainable solutions that involve closing the loop on the human P cycle. Some of these solutions are relatively straightforward but many involve overcoming considerable infrastructural or institutional inertia [Childers 2011: 123].
Economically viable mineral phosphorus reserves may become depleted within decades, threatening global crop production for a growing world population. The authors discuss this problem in relation to human P cycle, where the vast majority of mined phosphorus is not recycled back onto farm fields, but is released more or less irretrievably into the environment, polluting water bodies. “There are considerable social and environmental costs of P being lost from the currently ‘open’ human P cycle” [Childers 2011: 120].
The authors present several options for closing the human P cycle at the points of agricultural production, distribution and consumption, and human waste treatment. These options include reducing fertilizer application rates to better match plant needs, reducing erosion rates, reducing food waste, and recycling human urine, which is rich in phosphorus and nitrogen. The authors state that their list of solutions is not exhaustive, but rather is meant to stimulate others to think about the sustainability challenges of the human P cycle. Indeed, missing in this paper’s list of solutions is a discussion of the role of fungi, which can access otherwise inaccessible soil phosphorus through symbiosis with plants.
A broken biogeochemical cycle, Elser & Bennett 2011
Consider the fate of the approximately 17.5 million tonnes of phosphorus mined in 2005, analysed in the paper by Cordell et al. About 14 million tonnes of this were used in fertilizer (much of the rest went into cattle-feed supplements, food preservatives, and the production of detergents and industrial cleaning agents) but only about 3 million tonnes made it to the fork (or chopstick). The largest loss — around 8 million tonnes — was directly from farms through soil leaching and erosion” [Elser & Bennett 2011: 30].
To handle the twin problems of phosphorus pollution and scarcity, strategies for phosphorus conservation and recycling are urgently needed.
The solutions to these problems lie in recapturing and recycling phosphorus, moving it from where there is too much to where there is too little, and developing ways to use it more efficiently. Many strategies are simple and readily available, even for poor farmers and developing economies [Elser & Bennett 2011: 30].
The authors’ solutions include: widespread adoption of agricultural conservation practices, reduction of food waste (at least in part by producing food within or closer to cities), recycling human waste, reducing meat consumption, recovering nutrients from confined livestock facilities (such as through bioreactors), and genetically engineering plants and animals to require lower phosphorus inputs. No mention in this article of the role of plant-fungi symbiosis in accessing phosphorus in the soil.
Networks of power and influence: the role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning, Leake 2004
Two major groups of mycorrhizal fungi are arbuscular mycorrhiza (AM) and ectomycorrrhiza (EM). Both form a symbiosis with plants by colonizing their roots and creating an interface where carbon from the plant can be exchanged for phosphorus,nitrogen and other nutrients from the soil andtransferred to the plant by the fungi. The extraradical[15] mycorrhizal mycelium (ERMM), which are the vast portion of the fungal network that branches out into the soil, is difficult to study and has therefore been considered the “hidden half” of the symbiosis.
Progress in understanding the nature, extent, functioning, and identity of mycorrhizal fungal networks has been seriously hampered by the difficulties inherent in observing and studying mycelial systems without disturbing and destroying them.… As a consequence, the external mycelium, which is the fungal structure of mycorrhiza that is most intimately associated with the soil and furthest from the roots, and by implication the most critical for nutrient uptake, is normally overlooked and has been rarely recorded. Only in the past decade have studies started to focus specifically on the extent and functioning of ERMM in the field [Leake 2004: 1017].
This article highlights the significant yet overlooked role of mycorrhizal fungi in ecosystem functioning and reviews some advances in the techniques used to study these hidden powerhouses.
ERMM is the hidden power behind plant community composition and ecosystem functioning through the major processes it carries out, such as nutrient uptake, weathering of minerals, soil aggregate stability, and the way in which it alters competition between plants [Leake 2004: 1039].
The symbiosis with plants is the source of power for these fungi, given that the carbon received from plant hosts is practically inexhaustible and costs the plants little.
Despite the substantial biomass and associated C drain on their hosts, the actual “cost” of mycorrhiza to plants may be negligible because mycorrhizal colonization can increase the rate of photosynthesis (Wright et al. 1998), alleviate shoot N and P limitation, and cause a substantial increase in leaf area arising from improved nutrition (Read and Perez-Moreno 2003) [Leake 2004: 1021].
Thus,
The empowerment of mycorrhizal networks with substantial amounts of host-derived C allows them to play central roles in major biogeochemical cycles [Leake 2004: 1030].
The article concludes by emphasizing the importance for sustainable agriculture of a broader public understanding of the role of mycorrhiza for improving soil health and crop yields.
AM [arbuscular mycorrhizal] hyphal lengths in soil show strong positive correlations with soil-aggregate stability (Rillig et al. 2002; Kabir and Koide 2002), P uptake efficiency (Schweiger and Jakobsen 2000), and crop-yield improvements (Kabir and Koide 2002). Interest in the development of less intensive management systems is presenting new opportunities for adapting agricultural production systems to enhance these benefits that can be gained from AM networks. Substantial improvements in “soil health” and AM functioning in field crops are gained by the doubling of lengths of AM hyphae in soil when tillage is reduced (Kabir et al. 1998a, 1998b). Similar gains are achieved by growth of AM-compatible cover crops in place of winter fallow (Kabir and Koide 2002) [Leake 2004: 1038].
ERMM [extraradical mycorrhizal mycelium, or fungi] is the hidden power behind plant community composition and ecosystem functioning through the major processes it carries out, such as nutrient uptake, weathering of minerals, soil aggregate stability, and the way in which it alters competition between plants [Leake 2004: 1039].
Mycorrhizal symbioses influence the trophic structure of the Serengeti, Stevens 2018
Our analysis shows that inputs of phosphorus through arbuscular mycorrhizal symbioses substantially increase the ability of plants to grow and maintain nutritional quality, cascading through the biomass of consumers and predators in the ecosystem. Although they account for less than 1% of the total modelled biomass, the predicted nutritional benefit provided by arbuscular mycorrhizal fungi increased the biomass of macro-organisms in the Serengeti by 48%. When considering the management of biodiversity, future ecosystem models should account for the influence of arbuscular mycorrhizal fungi on all trophic levels [Stevens 2018: 536].
More than 70% of all angiosperm families form AM symbioses (Brundrett, 2009), and these symbioses are often essential for plant nutrition (Marschner & Dell, 1994). Mycorrhizal symbioses also improve plant tolerance to drought (Augé, 2001) and resistance to pathogens (Cameron, Neal, van Wees, & Ton, 2013) [Stevens 2018: 537].
Plant taxa vary in the degree to which they depend upon mycorrhizas; but in general, AM symbioses are essential for the nutrition of tropical plants, and warm season grasses are often highly dependent on mycorrhizas, acquiring up to 90% of their phosphorus requirements from AM fungi [Stevens 2018: 537].
Thirty years ago, McNaughton, Ruess, and Seagle (1988) concluded that large mammals have a major organising effect in the Serengeti ecosystem. From our analysis, we can conclude that AM fungi also play a critical role in the trophic structure of the Serengeti. Our model simulations suggest that although AM fungi account for less than 1% of the total biomass, phosphorus supplied by AM symbioses sustains half the vegetation biomass, and accordingly, half of the biomass of iconic migratory herbivores and one-third of the carnivore biomass [Stevens 2018: 542].
The distribution of soil phosphorus in the Serengeti, transported through AM symbioses and accelerated by migratory ungulates, may be a significant driver of plant diversity and ultimately mammalian carrying capacity (Anderson et al., 2007; McNaughton, Zuniga, McNaughton, & Banyikwa, 1997). Without AM fungal inputs of phosphorus, these nutrient diffusion gradients would undoubtedly decline [Stevens 2018: 543].
Rock-eating fungi, Jongmans 1997
Under a microscope, tiny tunnels can be seen in mineral particles from conifer forest soil. Scientists believe it is mycorrhizal fungi penetrating these particles by excreting organic acids in order to mine nutrients for their plant hosts. An estimated 150 meters of pores are bored by fungi per year per liter of E-horizon (layer that has been leached of mineral and/or organic content, leaving silicate) soil.
Photo credit: Jongmans 1997. “Scanning electron micrograph, showing 4–6-mm-thick hyphae entering a calcium feldspar at a granite surface near Lunsen, Sweden” [Jongman 1997].
The role of community and population ecology in applying mycorrhizal fungi for improved food security, Rodriguez & Sanders 2015
Given that nitrogen and phosphorus are the most limiting nutrients for crop growth, that global phosphorus supplies are becoming exhausted while the human population rapidly expands, and that arbuscular mycorrhizal fungi (AMF) symbioses improve crop phosphorus acquisition, AMF symbioses have a major role to play in current and future crop production.
The potential of AMF to help increase global food security lies in the fact that all globally important food crops naturally form this symbiosis and the fungi help plants more efficiently obtain phosphate from the soil (Smith and Read, 2008). Stocks of phosphate fertilizer are rapidly being depleted (Gross, 2010). There is a simultaneous increase in demand for phosphate to help feed the growing population (Gilbert, 2009). These two combined factors represent a major threat to global food security; a threat that can potentially be reduced by better phosphate acquisition through the AM[F] symbiosis. The potential of AMF to contribute to improved crop yields has been known for decades [Rodriquez & Sanders 2015: 1054].
However, for the widespread adoption of AMF inoculation to be effective and safe, a better understanding is needed of ecological principles related to soil fungi. The authors note that few studies have linked crop yield increases with successful colonization by an introduced AMF, and they outline several challenges and questions that should be resolved to pursue this promising technique more broadly. For example, they ask whether introduced AMF establish well, and how they affect native AMF populations, and how genetic diversity in AMF populations variously affects different crops.
Stocks of phosphate fertilizer are rapidly being depleted (Gross, 2010). There is a simultaneous increase in demand for phosphate to help feed the growing population (Gilbert, 2009). These two combined factors represent a major threat to global food security; a threat that can potentially be reduced by better phosphate acquisition through the [arbuscular mycorrhizal fungi] symbiosis [Rodriquez & Sanders 2015: 1054].
Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils, Rashid 2016
The paper argues for the use of bacterial and fungal inoculants in combination with organic amendments and cover crops to regenerate degraded soils. In order to produce enough food for a growing global population on ubiquitously degraded soils, synthetic fertilizers will be in increasingly high demand. However, these fertilizers require copious amounts of non-renewable energy to manufacture, and become pollutants when used. Here, the authors explain how bacteria and fungi make nutrients available to plants and how facilitate soil aggregation.
Meta-analysis of biofertilizer application in agriculture, Schutz 2018
Given the global decline of reserves of both rock phosphate and fossil fuel, this study poses the question – to what extent can microbial inoculants replace/reduce the use of synthetic fertilizer? The authors find that “dryland agriculture can benefit most from biofertilizers [microbial inoculants used as fertilizers]. Due to climate change, in the future there will be even more dryland areas globally. Biofertilizers are thus a promising option for sustainable agriculture” [Schutz 2018: 11]. More specifically:
Our comprehensive meta-analysis with studies from all over the world revealed that biofertilizers were found to be most effective in dry climates. Biofertilizer also improved PUE [phosphorus use efficiency] and NUE [nitrogen use efficiency] greatly. Furthermore, we found that biofertilizers possessing both N fixing and P solubilizing traits have the highest potential to improve the crop yields. Interestingly, AMFs, known for facilitating P nutrient uptake in plants, were on par with applications of biofertilizers with the combined traits of N fixation and P solubilization, indicating the big potential of AMFs as sole biofertilizer for most crops and climatic situations [Schutz 2018: 5].
Future Directions International Strategic Directions Paper: Agricultural Application of Mycorrhizal Fungi to Increase Crop Yields, Promote Soil Health and Combat Climate Change, Johns 2014
There are a number of agricultural practices that will enhance fungi colonisation. Wherever possible, of course, the full range of critical soil health processes that govern productivity should be allowed to regenerate agricultural ecologies naturally. It may, however, be necessary or more practical to inoculate seed with fungi spores in order to recover degraded soils. A number of farmers in the Great Southern agricultural region of Western Australia are undertaking this course of action. Finding themselves confronted with an unsustainable spiral of ever-increasing commercial fertiliser costs and uneconomic or diminishing crop yields, it was realised that a different approach needed to be taken. In recent growing seasons, seed has been inoculated with commercial fungi spores just prior to planting. While it is still too early to provide statistically robust outcomes and, bearing in mind that there are no “silver bullets” in agricultural production, the indications are that mycorrhizal fungi are promoting improvements in crop vitality, yield and soil condition [Johns 2014: 4].
Nitrogen: the double-edged sword, Jones 2014
The symbiosis between mycorrhizal fungi and plants drive carbon and nitrogen cycles. Fungi demand carbon exudate from plants in exchange for nitrogen and other nutrients retrieved and transported from the soil. The “liquid carbon” exuded from plant roots feeds mycorrhizal fungi and many other soil microbes, while also becoming stabilized in soil aggregates and humus. Jones explains that when this mycorrhizal exchange is inhibited by N fertilizer, which allows plants to absorb nitrogen “for free” (without providing liquid carbon in exchange), this reduces the flow of carbon into the soils, which in turn diminishes fungal networks and their delivery of micronutrients to plant hosts, and results in carbon-depleted soils.
Despite its abundance in the atmosphere, nitrogen is frequently the most limiting element for plants. There is a reason for this. Carbon, essential to photosynthesis and soil function, occurs as a trace gas, carbon dioxide, currently comprising 0.04% of the atmosphere. The most efficient way to transform CO2 to stable organic soil complexes (containing both C and N) is via the liquid carbon pathway. The requirement for biologically-fixed nitrogen drives this process.
If plants were able to access nitrogen directly from the atmosphere, their growth would be impeded by the absence of carbon-rich topsoil. We are witnessing an analogous situation in agriculture today. When inorganic nitrogen is provided, the supply of carbon to associative nitrogen fixing microbes is inhibited, resulting in carbon-depleted soils.
Reduced carbon flows impact a vast network of microbial communities, restricting the availability of essential minerals, trace elements, vitamins and hormones required for plant tolerance to environmental stresses such as frost and drought and resistance to insects and disease. Lowered micronutrient densities in plants also translate to reduced nutritional value of food [Jones 2014: 2-3].
Jones further explains how to modify agricultural practices to protect and build the soil: maintain year-round living ground cover, limit nitrogen and phosphorus fertilizer input, promote plant and microbial diversity, and integrate livestock into crop production systems.
Synthetic nitrogen fertilizers deplete soil nitrogen: a global dilemma for sustainable cereal production, Mulvaney 2009
There is a prevailing view that global food and fiber production will continue to expand because of modern agricultural management systems with improved cultivars and intensive chemical inputs dominated by synthetic ammoniacal fertilizers. The use of these fertilizers has led to concerns regarding water and air pollution but is generally perceived to play an essential role for sustaining agricultural productivity, not only by supplying the most important nutrient for cereal production but also by increasing the input of crop residues for building soil organic matter. The scientific soundness of the buildup concept has yet to be substantiated empirically using baseline data sets from long-term cropping experiments. The present paper and a companion study by Khan et al. (2007) provide many such data sets that encompass a variety of cereal cropping and management systems in different parts of the world. Overwhelmingly, the evidence is diametrically opposed to the buildup concept and instead corroborates a view elaborated long ago by White (1927) and Albrecht (1938) that fertilizer N depletes soil organic matter by promoting microbial C utilization and N mineralization. An inexorable conclusion can be drawn: The scientific basis for input-intensive cereal production is seriously flawed. The long-term consequences of continued reliance on current production practices will be a decline in soil productivity that increases the need for synthetic N fertilization, threatens food security, and exacerbates environmental degradation [Mulvaney 2009: 2308].
Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere, Paungfoo-Lonhienne 2015
In this study, nitrogen fertilization altered the relative abundance of fungal taxa in the rhizosphere, increasing fungal genera with known pathogenic traits, and decreasing a fungal phyla (Basidiomycetes) known to break down lignin, thus important for carbon cycling in the soil.
Fungi play important roles as decomposers, plant symbionts and pathogens in soils. The structure of fungal communities in the rhizosphere is the result of complex interactions among selection factors that may favour beneficial or detrimental relationships. Using culture-independent fungal community profiling, we have investigated the effects of nitrogen fertilizer dosage on fungal communities in soil and rhizosphere of field-grown sugarcane.The results show that the concentration of nitrogen fertilizer strongly modifies the composition but not the taxon richness of fungal communities in soil and rhizosphere. Increased nitrogen fertilizer dosage has a potential negative impact on carbon cycling in soil and promotes fungal genera with known pathogenic traits, uncovering a negative effect of intensive fertilization [Paungfoo-Lonhienne 2015:just 1].
Nitrogen fertilizer effects on soil carbon balances in Midwestern U.S. agricultural systems, Russell 2009
Despite increasing residue input in annual crop production systems, N fertilization does not increase soil organic carbon (SOC) over time because N fertilization also increases organic carbon (OC) decay. This study also shows that belowground OC inputs contribute to soil carbon sequestration more than aboveground OC inputs to the soil.
When all phases of the crop rotations were evaluated over the long term, OC decay rates increased concomitantly with OC input rates in several systems. Increases in decay rates with N fertilization apparently offset gains in carbon inputs to the soil in such a way that soil C sequestration was virtually nil in 78% of the systems studied, despite up to 48 years of N additions [Russell 2009: 1102].
Across all systems, SOC storage was significantly correlated with the quantity of belowground OM [organic matter] inputs (P < 0.01, both sites). In contrast, SOC was not correlated with the quantity of aboveground inputs (P = 0.45, Nashua; P = 0.55, Kanawha) [Russell 2009: 1111].
This study highlights the importance of incorporating both production and decomposition processes, as well as the location (above- or below-ground) of detrital inputs into models of N-fertilization effects on soil C dynamics in agroecosystems. These results are highly relevant for evaluating the potential of N fertilization to mitigate the effects of removal of organic-matter ‘‘residue’’ from the system for bioenergy production. Our data suggest that the stimulation of OC decomposition by the addition of fertilizer N would likely counteract the positive effects of N fertilization on inputs of OC to the soil, at least for annual crops. Given the current quantity of N that is applied over such a large area, management strategies that can maintain high yields and also reduce N-fertilizer use would also have beneficial environmental consequences. Our study indicates that selection of crops for higher belowground NPP [net primary production], in rotation with crops that fix N, could maximize both yields and soil C sequestration without excessive N-fertilizer additions [Russell 2009: 1111].
Community proteogenomics reveals the systemic impact of phosphorus availability on microbial functions in tropical soil, Yao 2018
In this study, long-term phosphorus fertilization limited the extent to which the genes and proteins of microbial communities were allocated to degrading recalcitrant soil phytate to acquire phosphorus. In phosphorus-deficient soil, on the other hand, the genes responsible for breaking down recalcitrant substrate to acquire phosphorus were more prevalent in microbial communities. In other words, microbial communities can adapt genetically to different levels of nutrients in the soil in order to continue meeting their nutritional requirements. This adds to the body of evidence that fertilizer use impairs the inherent qualities of a living soil to nourish the plants growing there.
A greater than fourfold increase in the gene abundance of 3-phytase was the strongest response of soil communities to phosphorus deficiency. Phytase catalyses the release of phosphate from phytate, the most recalcitrant phosphorus-containing compound in soil organic matter. Genes and proteins for the degradation of phosphorus-containing nucleic acids and phospholipids, as well as the decomposition of labile carbon and nitrogen, were also enhanced in the phosphorus-deficient soils. In contrast, microbial communities in the phosphorus-rich soils showed increased gene abundances for the degradation of recalcitrant aromatic compounds, transformation of nitrogenous compounds and assimilation of sulfur. Overall, these results demonstrate the adaptive allocation of genes and proteins in soil microbial communities in response to shifting nutrient constraints [Yao 2018: 499].
In conclusion, our proteogenomics results provide systems biology insights into the adaptation of soil microbial communities to different levels of phosphorus availability in a humid tropical forest environment. Phosphorus deficiency significantly enhanced the genetic capabilities of microbial communities to extract phosphorus from phytate and, to a lesser extent, from nucleic acids and phospholipids. Long-term phosphorus fertilization altered the allocation of genes and proteins by microbial communities to acquire carbon, nitrogen and sulfur from a variety of substrates. The results suggest that the selective degradation of recalcitrant substrates, including phytate in phosphorus-deficient soils and aromatic compounds in phosphorus-rich soils, is an important means for microbial communities to balance their elemental requirements. The adaptive allocation of genes and proteins for acquisition of these nutrients in different soils can be explained as an optimal foraging strategy by which microbial communities maintain efficient growth under resource limitation [Yao 2018: 505].
Riparian[14] ecosystems are naturally resilient, provide linear habitat connectivity, link aquatic and terrestrial ecosystems, and create thermal refugia for wildlife: all characteristics that can contribute to ecological adaptation to climate change [Seavy 2009: 330].
Arguing for the restoration of riparian areas because of their ecological significance and inherent resilience, these authors articulate the importance of both surface and groundwater – protected within a biodiverse ecosystem – for its cooling effect.
Because riparian areas have higher water content than surrounding upland areas, they absorb heat and buffer organisms against extreme temperatures (Naiman et al. 2000). During previous periods of climate change, riparian areas served as refugia because they provided microclimates that protected plant biodiversity (Bakker 1984, Meave and Kellman 1994). Riparian vegetation can maintain cooler water temperatures by shading water from sunlight (Sridhar et al. 2004, Cassie 2006) and the infusion of cold groundwater into warmer surface waters creates and maintains pockets of cool water (Chu et al. 2008). Thus, riparian areas provide thermal refugia for animals with thermoregulatory limitations [Seavy 2009: 332].
The importance of water retention (the rainfall absorbed or used within an ecosystem) for mitigating flood and drought conditions and contributing to clean drinking water, for example, has been increasingly recognized in Europe in the past decade. Along with wetland preservation, better agriculture practices and other measures, preserving and re-growing forests are seen as key to enhanced natural water retention. Forests cover a third of Europe, and:
can soak up excess rainwater, preventing run-offs and damage from flooding. By releasing water in the dry season, forests can help to provide clean water and mitigate the effects of droughts [EEA 2015: 6].
In recognition of the important water management role of forests and other natural ecosystems, new policy instruments have proposed Natural Water-Retention Measures (NWRMs).
Natural Water-Retention Measures (NWRMs) are defined as ‘measures to protect and manage water resources and to address water-related challenges by restoring or maintaining ecosystems, natural features and characteristics of water bodies using natural means and processes’ (European Commission and Directorate-General for the Environment 2014). … The main focus is to enhance and preserve the water retention capacity of aquifers, soil and ecosystems and improve their status [EEA 2015: 9].
This EEA study found that:
In water-basins where the forest cover is 30%, water retention is 25% higher than in basins where the forest cover is only 10%. In basins where the forest cover is 70%, water retention is 50% higher than in basins where the forest cover is only 10%. … Coniferous forests in general retain 10% more water than broadleaved forests or mixed forests [EEA 2015: 5].
When managed with both climate adaptation and mitigation in mind, tropical reforestation (TR) can serve multiple synergistic functions. TR mitigates regional and global climate change, not only by sequestering carbon but also through biophysical cooling (via evapotranspiration), by recycling rainfall regionally, and by reducing pressure on old growth forests.
Furthermore, TR helps local communities adapt to climate change by recharging stream flow in the dry season, reducing the severity of floods, protecting slopes against landslides and, through mangrove reforestation, reducing the impact of coastal storms and waves. Reforestation also creates livelihood opportunities through the sustainable harvest of forest products, and creates shelter and habitat for species vulnerable to climate change. However, to achieve this broad range of benefits, “reforestation practices should be designed to avoid the implementation of one strategy (mitigation or adaptation) to the detriment of the other.” Arguing for the application of what they term “climate-smart reforestation,” the authors recommend the following:
The challenge for climate-smart reforestation is to implement an effective combination of approaches to meet all three objectives: societal adaptation, climate mitigation, and ecological resilience [Locatelli 2015: 4].
However, as most policies consider the three objectives of climate-smart reforestation separately, they often overlook possible trade-offs and synergies. For example,reforestation projects managed with a carbon purpose could have detrimental consequences on water availability in the semi-arid tropics (Trabucco et al. 2008) or on biodiversity (O’Connor 2008). By contrast, reforestation that is explicitly climate-smart uses a multi-objective planning focus that enables different objectives to reinforce each other so that their interactions produce synergies rather than trade-offs. For example, tree regeneration in Tanzania under the Ngitili resource management system achieves carbon storage together with improved watershed conservation and greater provision of natural resources (water, food, and fodder) for livelihoods (Duguma et al. 2014). A proposed adaptation project in Colombia aims to reforest with flood-resistant native tree species to reduce flood impacts on downstream communities (UNDP 2012). A project in Costa Rica is testing different mixes of species and silvicultural practices to reduce vulnerability to storms and fires while also achieving carbon storage (Locatelli et al. 2011) [Locatelli 2015: 4-5].
This article underscores a key concept of this compendium – that functioning ecosystems (whether old growth or restored forests, for example) provide multiple, interwoven functions that support human and biodiverse life by regulating local, regional and global climate conditions.
The Indian summer monsoon rainfall has decreased since 1950, and several hypotheses have been proposed to explain why. Most of these hypotheses involving weakening temperature gradients over the continent. This study explores the potential link between a weakening monsoon and widespread land use land cover (LULC) change from woody savanna to cropland in recent decades. Citing earlier studies, the authors note that:
Precipitation resulting from local land surface Evapotranspiration (ET) is known as recycled precipitation. Pathak et al. found that evapotranspiration from land surface vegetation plays a major role during the end of a monsoon. They observed that, during the initial phase of a monsoon, oceanic sources play a major role, and the soil is recharged with moisture. However, during the latter half of a summer monsoon (August and September), land surface ET increases as recycled precipitation increases, a pattern that is more prominent in the Ganga Basin and Northeast India. This recycled precipitation accounts for approximately 20–25% of the rainfall in North India (Ganga Basin) and Northeast India during August and September. Hence, deforestation associated with changes in LULC may affect ET and may significantly affect monsoon rainfall [Paul 2016: 1-2].
Summarizing their own study, the authors conclude:
Here, we performed a sensitivity analysis to quantify the impacts of large-scale conversion from woody savannah to crop land in India on monsoon precipitation. We found such a change results in decreased ET and subsequently a decrease in recycled precipitation leading to a decline in monsoon precipitation. This decline is similar in extent to the observed recent decadal weakening of monsoon precipitation. However, other reasons may account for this observed weakening, such as the warming of Indian Ocean SST [sea surface temperature] [Paul 2016: 5-6].
To better clarify causality of the weakening monsoon, given other potential factors, the authors propose that:
The future scope of this present work is to perform detection and attribution studies for potential declines of Indian monsoons with model runs forced with SST warming only, aerosol forcing only, LULC changes only and all controlling factors together [Paul 2016: 6].
This study finds that the physiological response of plants to increased atmospheric CO2 affects the global hydrological cycle even more than does the greenhouse effect and changes in precipitation. The authors conclude:
This highlights the key role of vegetation in controlling future terrestrial hydrologic response and emphasizes that the carbon and water cycles are intimately coupled over land [LeMordant 2018: 1].
With increasing [CO2] at the leaf surface, the density of stomata at the leaf surface is decreased and their individual opening is reduced and therefore less water is transpired per unit leaf area. In other words, leaf-level water use efficiency increases, potentially increasing surface soil moisture and runoff. On the other hand, leaf biomass tends to also increase with increasing [CO2] … generating a larger evaporative surface that can partly offset the reduction in stomatal conductance and negate the soil water savings. Our objective is therefore to quantify how such plant [CO2] effects influence future hydrological variable responses compared with radiative effects ––the atmospheric impact of the “greenhouse effect.” Radiative effects impact precipitation, i.e., water supply, and evaporative demand, through increase in radiation, temperature, and atmospheric dryness as estimated by the vapor pressure deficit (VPD), i.e., saturation minus actual vapor pressure [LeMordant 2018: 1].
Our study illustrates how deeply the physiological effects [on vegetation] due to increasing atmospheric [CO2] impact the continental water cycle. Contrary to previous wisdom, changes in precipitation and radiation [greenhouse effect] do not play the primary role in future drying and moistening in most regions. Rather, biosphere physiological effects and related biosphere–atmosphere interactions are key for predicting future continental water stress as represented by ET [evapotranspiration], long-term runoff, EF, or leaf area index. In turn, vegetation water stress largely regulates land carbon uptake, further emphasizing how tightly the future carbon and water cycles are coupled so that they cannot be evaluated in isolation [LeMordant 2018: 5].
Responding to a common belief that trees lower groundwater infiltration due to transpiration, and a contrasting view that trees increase groundwater infiltration by increasing organic matter and soil porosity, these authors test an “optimum tree cover theory.”
They find that “intermediate” tree cover maximizes groundwater recharge in the tropics, resulting in a 2-14% increase in total annual water input from rainfall. However, the tree species used in this study consume more water compared to many other tree species in the semi-arid tropics. Therefore, the results here may be conservative in terms of the potential of trees to increase groundwater recharge. Furthermore, the study doesn’t consider the potential effects of greater transpiration from increased tree cover on local rainfall patterns.
Noting that “major increases in crop productivity and changes in regional climate are generally collocated in time and space over the central United States” [Alter 2018: 1587], the study tested the hypothesis that there is a causal relationship – that historical agricultural intensification has affected regional summer climate in this area.
… from 1950 to 2010, the amount of corn harvested annually in the Corn Belt increased by 400%, from 2 billion to 10 billion bushels (National Agricultural Statistics Service, 2016) [Alter 2018: 1586].
and
From 1910 to 1949 (pre-agricultural development, pre-DEV) to 1970–2009 (full agricultural development, full-DEV), the central United States experienced large-scale increases in rainfall of up to 35% and decreases in surface air temperature of up to 1°C during the boreal summer months of July and August, when crop water use in the Corn Belt is at its peak [Alter 2018: 1586].
The authors used a regional climate model to test their hypothesis by comparing a set of simulations where “enhanced photosynthesis over cropland [serves] as a proxy for agricultural intensification” [Alter 2018: 1589] to a control simulation with no agricultural intensification. They found that:
Over the region that has experienced significant increases in observed rainfall (region of significant change—ROSC), the mean rainfall increase is ~7% (0.20mm/d) for the simulations and ~15% (0.37mm/d) for the observations. Thus, it seems that agricultural intensification has been a major contributor to the observed increase in summer rainfall in the central United States [Alter 2018: 1589].
Strikingly, these increases in rainfall are also very consistent: Agricultural intensification enhances simulated rainfall across the aforementioned swath in the central United States during at least 62% of the 150 ensemble years (significant at the 5% level using the chi-square test). In the observational data, a similar consistency in precipitation enhancement is evident when comparing the pre-DEV and full-DEV time periods. This suggests that the changes in rainfall due to agricultural intensification are not the result of occasional increases but instead are indicative of a more systematic change in the summer rainfall regime of the central United States [Alter 2018: 1589].
This study usefully contributes evidence that vegetation cover affects local and regional climates, while drawing conclusions, however, that are not necessarily helpful to understanding how to mitigate and adapt to climate change. The study’s findings suggest that agricultural intensification can potentially mitigate local climate change effects in the future, but it is unlikely that the methods that drove agricultural intensification in the 20th Century will continue to work in a changing climate. The reason that these methods are now obsolete is that they strip the soils of the organic material and living organisms necessary for the resilience of plants, and their ability to cope with droughts, floods, heat and other challenging conditions.
The model here uses “enhanced photosynthesis” as a proxy for agricultural intensification. While the increase in yield between early and late 20th Century Corn Belt production represents an increase in photosynthesis, high-input agriculture is but one pathway to enhanced photosynthesis. Moreover, it is an extremely problematicone with respect to climate change, given the high energy costs of fertilizer, pesticides and fuel, and the damage to the soils from these practices.
Instead, a useful lesson to draw from this study is simply that enhanced photosynthesis itself can mitigate climate change regionally. In the context of agricultural production in the era of climate change, enhanced photosynthesis might best be accomplished through ecological intensification, a strategy for improving resilience within an agro-ecosystem, and thereby greater photosynthesis and more reliable crop production.
Deforestation contributes to climate change on a global scale through carbon emissions (biogeochemical effects), and on a local/regional scale through biogeophysical effects related to albedo, evapotranspiration and roughness, affecting surface energy budgets.
Here, we show that historical deforestation has led to a substantial local warming of hot days over the northern mid-latitudes – a finding that contrasts with most previous model results. Based on observation-constrained state-of-the-art climate-model experiments, we estimate that moderate reductions in tree cover in these regions have contributed at least one-third of the local present-day warming of the hottest day of the year since pre-industrial time, and were responsible for most of this warming before 1980 [LeJeune 2018: 1].
The study uses observational data to constrain the outcome of a climate model simulating the effects of deforestation on regional temperatures. The authors found that during most of the 20th century, the biogeophysical effects of deforestation were the main cause of regional temperature increases, and that by 1980 deforestation in northern mid-latitudes had declined. By that time other forcings began to take on a proportionally greater role in regional temperature increases.
This climate model simulation illustrates how the biosphere affects the climate system. With “maximum vegetation,” more water is absorbed in the ground, allowing for evaporation to cool the land surface while also recycling more rain. This simulation resulted in an average temperature reduction over land of 1.2C.
The authors describe their approach:
We quantify the maximum possible influence of vegetation on the global climate by conducting two extreme climate model simulations: in a first simulation (‘desert world’), values representative of a desert are used for the land surface parameters for all non-glaciated land regions. At the other extreme, a second simulation is performed (‘green planet’) in which values are used which are most beneficial for the biosphere’s productivity [Kleidon 2000: 471].
They describe the effects of maximum vegetation on the water cycle, stating that over land:
…the hydrological cycle is more active, with precipitation roughly increasing by 100%, evapotranspiration by more than 200% and the mean moisture content of the atmosphere (or precipitable water) increasing by 30%. These increases can be understood by enhanced recycling of soil water as a response of both, (i) more absorbed radiation at the surface so that more energy is available for evapotranspiration and (ii) larger soil water storage capacities (SWCs) which enhance water availability during dry periods. This increased recycling also leads to an overall decrease in continental runoff by about 25% [Kleidon 2000: 476].
Changes in the water cycle result in land surface temperature changes:
The substantial increase in evapotranspiration is associated with differences in the surface energy balance, primarily concerning the partitioning between sensible and latent heat. The latent heat flux increases by the same amount (more than 200%) as evapotranspiration and the sensible heat flux decreases to 30% of its original value. … Subsequently, the increased latent heat flux leads to more efficient cooling of the surface, resulting in temperatures reduced by 1.2 K[13] [Kleidon 2000: 477-478].
Human modification of the hydrological cycle has profoundly affected the flow of liquid water across the Earth’s land surface. Compared to changes to liquid water flow, alteration of water vapor flows through land-use changes has received comparatively less attention, despite compelling evidence that such alteration can influence the functioning of the Earth System.
We show that deforestation is as large a driving force as irrigation in terms of changes in the hydrological cycle. Deforestation has decreased global vapor flows from land by 4% (3,000 km3/yr), a decrease that is quantitatively as large as the increased vapor flow caused by irrigation (2,600 km3/yr). Although the net change in global vapor flows is close to zero, the spatial distributions of deforestation and irrigation are different, leading to major regional transformations of vapor-flow patterns [Gordon 2015: 7612].
Highlighting the significance of Makarieva and Gorshkov’s “biotic pump” hypothesis (above), Sheil and Murdiyarso explain it in layman’s terms in this article for the benefit of a broader public, and examine its validity. They point out that the biotic pump hypothesis offers an explanation for a question not otherwise resolved in conventional climate theory.
Conventional theory offers no clear explanation for how flat lowlands in continental interiors maintain wet climates. Makarieva and Gorshkov show that if only “conventional mechanisms” (including [rain] recycling) apply, then precipitation should decrease exponentially with distance from the oceans. Researchers have previously puzzled over a missing mechanism to account for observed precipitation patterns (Eltahir 1998) [Sheil & Murdiyarso 2009: 342].
They explain the biotic pump hypothesis and how it resolves the puzzle:
Air currents near Earth’s surface flow to where pressure is lowest. According to Makarieva and Gorshkov, these are the areas that possess the highest evaporation rates. In equatorial climates, forests maintain higher evaporation rates than other cover types, including open water. Thus, forests draw in moist air from elsewhere; the larger the forest area, the greater the volumes of moist air drawn in. This additional moisture rises and condenses in turn, generating a positive feedback in which a large proportion of the water condensing as clouds over wet areas is drawn in from elsewhere. The drivers (solar radiation) and basic thermodynamic concepts and relationships are the same as in conventional models, thus most behaviors are identical— the difference lies in how condensation is incorporated.
Makarieva and Gorshkov’s estimates, incorporating volume changes from condensation, imply that when forest cover is sufficient, enough moist air is drawn in to maintain high rainfall inside continents. The numbers now add up: thus, condensation offers a mechanism to explain why continental precipitation does not invariably decline with distance from the ocean [Sheil & Murdiyarso 2009: 342].
Commenting on the relevance of the hypothesis, the authors conclude:
Acceptance of the biotic pump would add to the values that society places on forest cover. By raising regional concerns about water, acceptance of Makarieva and Gorshkov’s biotic pump demands attention from diverse local actors, including many who may otherwise care little for maintaining forest cover [Sheil & Murdiyarso 2009: 346].
The authors examine ecological and geophysical principles to explain how land far inland away from the ocean can remain moist, given that gravity continuously pulls surface and groundwater into the ocean over time.
All freshwater on land originates in the ocean from which it has evaporated, is carried on air flux, and precipitates over the land. Coastal regions benefit from this cycle by their proximity to the ocean, yet in the absence of natural forests in coastal regions precipitation weakens as distance from the ocean increases, leaving inland areas arid. The authors propose the concept of a biotic pump to explain how large continents can be sufficiently moist deep into the interior, and abundant with rivers and lakes.
Air and moisture are pulled horizontally by evapotranspiration over coastal forests. When water vapor from plants condenses, it creates a partial vacuum that pulls water evaporating from the ocean into the continental interior which results in forest rains. By contrast, deserts are unable to pull in ocean evaporation because they lack evaporative force.
Therefore, ongoing deforestation, especially coastal deforestation on a large scale, threatens to cut off rain to the interiors of Earth’s continents, thereby creating new deserts. The Amazonian rainforest is the prime example: Deforestation of the eastern coast of South America has led to changes in the rainforest that is resulting in drying and desertification of the interior, with unprecedented fires and loss of rivers. Historically, Australia’s interior became a desert around the time the first humans arrived on the continent, and the authors speculate that early coastal deforestation was the cause. On the other hand, restoring natural coastal forests can also restore inland water cycles and reverse desertification.
This article illustrates the importance of biological relationships that are ecologically complex and poorly understood. It highlights the significance of the precautionary principle in assessing what we don’t know when altering ecological processes, and taking preventive action in the face of uncertainty.
This paper takes the innovative and paradigm-shifting position that carbon is not the primary consideration in climate; rather, water should be the central focus, integrated with carbon and energy cycles:
Forest-driven water and energy cycles are poorly integrated into regional, national, continental and global decision-making on climate change adaptation, mitigation, land use and water management. This constrains humanity’s ability to protect our planet’s climate and life-sustaining functions. The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. The effects of tree cover on climate at local, regional and continental scales offer benefits that demand wider recognition. The forest- and tree-centered research insights we review and analyze provide a knowledge-base for improving plans, policies and actions. Our understanding of how trees and forests influence water, energy and carbon cycles has important implications, both for the structure of planning, management and governance institutions, as well as for how trees and forests might be used to improve sustainability, adaptation and mitigation efforts [Ellison 2017: Abstract].
Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. [Ellison 2017: Abstract].
Vegetation cover affects the amount of solar energy a land area absorbs and/or releases, thus altering local temperatures and precipitation. Plants regulate local temperatures through shading, albedo and evapotranspiration, which releases latent[9] heat.
The ability of a surface to shed energy through latent or sensible heat is key to determining that surface’s temperature – shifts in the relative balance between the two can lead to increases in surface temperatures (where sensible heat is relatively higher) or decreases (where latent heat is relatively higher) [Swann 2018: 2].
This study shows that changes in vegetation cover in a given place affect not only the local climate, but also the climate system at a continental scale. The results are temperature and precipitation changes in remote parts of the continent relative to where the tree loss occurred, leading to changes in ecosystem productivity in those remote parts. This phenomenon is called ‘ecoclimate teleconnections.’
Plants profoundly influence local climate by controlling the exchange of energy and water with the atmosphere. Changes in and/or losses of plant type or plant functioning can alterthe local climate, but also potentially large scale climate by modifying atmospheric circulation. … the potentially global impact of plant cover change on other ecosystems as communicated by the atmosphere has been under-appreciated and is only beginning to be evaluated [Swann 2018: 2].
Researchers simulated tree die-offs in their model by replacing all trees in a given domain[10] with grass.
Domain-scale tree loss led to changes in local (within same domain) surface properties and fluxes including albedo and evapotranspiration. These changes in surface properties modified local surface climate (e.g., precipitation and temperature), as well as impacted atmospheric circulation. The atmospheric circulation response connects the direct forcing of tree loss on the local atmosphere to other regions, impacting climate and thus resulting in altered Gross Primary Productivity (GPP) across North America [Swann 2018: 3-4].
Furthermore, the severity of the remote effects of tree loss depends not only on the scale of the tree loss, but also on the location of the tree loss. The study found, for example, that tree loss in an area covering most of California had greater effect on GPP in other parts of the continent than did tree loss of a similar scale elsewhere.
Thus, in addition to the magnitude of forest loss, the location of forest loss plays an outsized role in determining the continental scale impact [Swann 2018: 6].
Regenerating the soil carbon sponge is our greatest point of leverage for salvaging the planet from the point of existential climate crisis. “Sponge” refers to the quality of a biologically active soil with high organic matter content to have lots of pore space for water absorption. Jehne states that every additional gram of soil carbon allows the soil to hold 8 additional grams of water. He emphasizes the soil sponge concept because it is the driver of healthy ecosystems, and also within our control to repair and regenerate.
Jehne explains that an average of 342 W/m2 of incident solar radiation enters the troposphere while just 339 W/m2 is reflected back into space due to the greenhouse effect [Jehne 2018: 19:00 min]. This leaves a continuous energy balance of 3 W/m2 heating up the planet. Of the solar radiation returning to space, 24% is released through latent heat fluxes from evapotranspiration [Jehne 2018: 1:34:15]. However, due to land use change, there is 50% less transpiration on Earth than there was some 8,000 years ago. Jehne estimates that increasing transpiration by only 5% would be enough to offset the 3 W/m2 surplus solar energy [Jehne 2018: 1:34:50].
Increasing transpiration is achieved by increasing vegetation cover, which in turn is achieved by regenerating the soil sponge. Jehne explains that conventional agriculture has employed techniques (such as burning, cultivating/tilling, applying fertilizer and pesticides, and use of irrigation and fallow) that quickly oxidize the carbon fixed by plants through photosynthesis. By contrast, regenerative agriculture builds up the soil carbon sponge by facilitating the ecological processes that create stable soil carbon and limit organic matter breakdown.
In addition to the cooling effect from the latent heat flux, transpiration also provides the moisture needed for cloud formation. Jehne states that a 2% increase in cloud cover, given its high albedo, is also enough to reflect the excess 3 W/m2 solar radiation that is otherwise absorbed on Earth [Jehne 2018: 1:39:25]. Furthermore, bacteria released from ecosystems serve as the most effective precipitation nuclei[8] for making rain.
Vegetation cover cools Earth when it intercepts the sun’s energy. This is not just by providing shade, but also through evapotranspiration, which is how plants regulate their own internal temperatures.
For a plant … transpiration[5] is a necessity by which a plant maintains its inner environment within the limit of optimal temperatures. And at the level of landscape, evapotranspiration is the most efficient air conditioning system developed by nature [Eiseltova 2012:10].
The water in plant tissues contains the sun’s energy in the form of latent heat, which is released from plants through evapotranspiration. In the absence of water, solar energy reaching Earth becomes sensible heat – the heat we can feel and measure in rising temperatures.
Without water, the energy of the incoming radiation is transformed into sensible heat and the local area becomes overheated during the day and likewise far cooler at night (as is well known from desert areas, with differences between day and night temperatures typically exceeding 50°C). Water-saturated landscapes provide much more stable environments than do dry terrestrial systems. In landscapes with water – abundant aquatic ecosystems, wetlands and soils with high water retention capacity – about 80% of incoming solar energy is stored as latent heat of water vapour via evapotranspiration, whilst in de-watered landscapes (with a low-water retention capacity) the vast majority of solar energy is transformed into sensible heat (Pokorný et al. 2010b) [Eiseltova 2012: 307].
With respect to landscape management for sustainability, the authors introduce the idea of a “dissipative-ecological-unit,” meaning “the smallest functional unit that is capable of forming internalized cycles of matter and water while dissipating energy” [Eiseltova 2012: 312]. This term emphasizes the importance of small, local water cycles, which occur naturally in undisturbed ecosystems, resulting in “an efficient local resource economy and … relatively even temperatures and moisture conditions” [Eiseltova 2012: 312].
In catchments with a well-developed vegetation cover, water and matter are bound to short-circuited cycles and losses are minimal. In contrast, the increased clearance of forest, exposure of bare land, and drainage of agricultural land have accelerated matter losses from catchments [Eiseltova 2012: 11].
There is an urgent need that agricultural research focuses on how to close water cycles[6] in the landscape and the development of farming systems with a more vertically-layered vegetation structure keeping water and lower temperatures during a sunny day[7] [Eiseltova 2012: 324].
The water cycle is akin to the ‘bloodstream’ of the biosphere. Returning water to the landscape and restoring more natural vegetation cover is the only way to restore landscape sustainability. More attention in present-day science needs to be devoted to the study of the role of vegetation in the water cycle and climate amelioration. Restoration of a more natural vegetation cover over the landscape seems to be the only way forward.
Based on our current scientific knowledge, we can propose two criteria for assessing sustainable land management. These criteria are: the efficiency of an ecosystem to recycle water and matter,and its efficiency to dissipate solar energy. Land managers can substantially contribute to the restoration of the water cycle, climate amelioration and reduction of irreversible matter [soil and nutrient] losses with river water flows to the sea.
It is in the interest of society as a whole that land managers (farmers,foresters) be rewarded for their actions towards sustainable management of their land. Suitable tools to assess the achievements of individual land managers with respect to sustainable management of their land are: (1) continuous monitoring of conductivity – a measure of dissolved load – and flow rates in streams in order to estimate matter losses; and (2) the regular evaluation of satellite thermal channel images to assess temperature damping, i.e., the effectiveness of land use to dissipate solar energy. Restoration of natural ‘cooling structures’ – vegetation with its evapotranspiration and condensation-induced water circulation – is essential to renew landscape sustainability [Eiseltova 2012: 325].
Evapotranspiration – A Driving Force in Landscape Sustainability, Eiseltová 2012
Vegetation cover cools Earth when it intercepts the sun’s energy. This is not just by providing shade, but also through evapotranspiration, which is how plants regulate their own internal temperatures.
For a plant … transpiration[5] is a necessity by which a plant maintains its inner environment within the limit of optimal temperatures. And at the level of landscape, evapotranspiration is the most efficient air conditioning system developed by nature [Eiseltova 2012:10].
The water in plant tissues contains the sun’s energy in the form of latent heat, which is released from plants through evapotranspiration. In the absence of water, solar energy reaching Earth becomes sensible heat – the heat we can feel and measure in rising temperatures.
Without water, the energy of the incoming radiation is transformed into sensible heat and the local area becomes overheated during the day and likewise far cooler at night (as is well known from desert areas, with differences between day and night temperatures typically exceeding 50°C). Water-saturated landscapes provide much more stable environments than do dry terrestrial systems. In landscapes with water – abundant aquatic ecosystems, wetlands and soils with high water retention capacity – about 80% of incoming solar energy is stored as latent heat of water vapour via evapotranspiration, whilst in de-watered landscapes (with a low-water retention capacity) the vast majority of solar energy is transformed into sensible heat (Pokorný et al. 2010b) [Eiseltova 2012: 307].
With respect to landscape management for sustainability, the authors introduce the idea of a “dissipative-ecological-unit,” meaning “the smallest functional unit that is capable of forming internalized cycles of matter and water while dissipating energy” [Eiseltova 2012: 312]. This term emphasizes the importance of small, local water cycles, which occur naturally in undisturbed ecosystems, resulting in “an efficient local resource economy and … relatively even temperatures and moisture conditions” [Eiseltova 2012: 312].
In catchments with a well-developed vegetation cover, water and matter are bound to short-circuited cycles and losses are minimal. In contrast, the increased clearance of forest, exposure of bare land, and drainage of agricultural land have accelerated matter losses from catchments [Eiseltova 2012: 11].
There is an urgent need that agricultural research focuses on how to close water cycles[6] in the landscape and the development of farming systems with a more vertically-layered vegetation structure keeping water and lower temperatures during a sunny day[7] [Eiseltova 2012: 324].
The water cycle is akin to the ‘bloodstream’ of the biosphere. Returning water to the landscape and restoring more natural vegetation cover is the only way to restore landscape sustainability. More attention in present-day science needs to be devoted to the study of the role of vegetation in the water cycle and climate amelioration. Restoration of a more natural vegetation cover over the landscape seems to be the only way forward.
Based on our current scientific knowledge, we can propose two criteria for assessing sustainable land management. These criteria are: the efficiency of an ecosystem to recycle water and matter,and its efficiency to dissipate solar energy. Land managers can substantially contribute to the restoration of the water cycle, climate amelioration and reduction of irreversible matter [soil and nutrient] losses with river water flows to the sea.
It is in the interest of society as a whole that land managers (farmers,foresters) be rewarded for their actions towards sustainable management of their land. Suitable tools to assess the achievements of individual land managers with respect to sustainable management of their land are: (1) continuous monitoring of conductivity – a measure of dissolved load – and flow rates in streams in order to estimate matter losses; and (2) the regular evaluation of satellite thermal channel images to assess temperature damping, i.e., the effectiveness of land use to dissipate solar energy. Restoration of natural ‘cooling structures’ – vegetation with its evapotranspiration and condensation-induced water circulation – is essential to renew landscape sustainability [Eiseltova 2012: 325].
New climate solutions, water cycles and the soil carbon sponge, Jehne 2018
Regenerating the soil carbon sponge is our greatest point of leverage for salvaging the planet from the point of existential climate crisis. “Sponge” refers to the quality of a biologically active soil with high organic matter content to have lots of pore space for water absorption. Jehne states that every additional gram of soil carbon allows the soil to hold 8 additional grams of water. He emphasizes the soil sponge concept because it is the driver of healthy ecosystems, and also within our control to repair and regenerate.
Jehne explains that an average of 342 W/m2 of incident solar radiation enters the troposphere while just 339 W/m2 is reflected back into space due to the greenhouse effect [Jehne 2018: 19:00 min]. This leaves a continuous energy balance of 3 W/m2 heating up the planet. Of the solar radiation returning to space, 24% is released through latent heat fluxes from evapotranspiration [Jehne 2018: 1:34:15]. However, due to land use change, there is 50% less transpiration on Earth than there was some 8,000 years ago. Jehne estimates that increasing transpiration by only 5% would be enough to offset the 3 W/m2 surplus solar energy [Jehne 2018: 1:34:50].
Increasing transpiration is achieved by increasing vegetation cover, which in turn is achieved by regenerating the soil sponge. Jehne explains that conventional agriculture has employed techniques (such as burning, cultivating/tilling, applying fertilizer and pesticides, and use of irrigation and fallow) that quickly oxidize the carbon fixed by plants through photosynthesis. By contrast, regenerative agriculture builds up the soil carbon sponge by facilitating the ecological processes that create stable soil carbon and limit organic matter breakdown.
In addition to the cooling effect from the latent heat flux, transpiration also provides the moisture needed for cloud formation. Jehne states that a 2% increase in cloud cover, given its high albedo, is also enough to reflect the excess 3 W/m2 solar radiation that is otherwise absorbed on Earth [Jehne 2018: 1:39:25]. Furthermore, bacteria released from ecosystems serve as the most effective precipitation nuclei[8] for making rain.
Continental-scale consequences of tree die-offs in North America: identifying where forest loss matters most, Swann 2018
Vegetation cover affects the amount of solar energy a land area absorbs and/or releases, thus altering local temperatures and precipitation. Plants regulate local temperatures through shading, albedo and evapotranspiration, which releases latent[9] heat.
The ability of a surface to shed energy through latent or sensible heat is key to determining that surface’s temperature – shifts in the relative balance between the two can lead to increases in surface temperatures (where sensible heat is relatively higher) or decreases (where latent heat is relatively higher) [Swann 2018: 2].
This study shows that changes in vegetation cover in a given place affect not only the local climate, but also the climate system at a continental scale. The results are temperature and precipitation changes in remote parts of the continent relative to where the tree loss occurred, leading to changes in ecosystem productivity in those remote parts. This phenomenon is called ‘ecoclimate teleconnections.’
Plants profoundly influence local climate by controlling the exchange of energy and water with the atmosphere. Changes in and/or losses of plant type or plant functioning can alterthe local climate, but also potentially large scale climate by modifying atmospheric circulation. … the potentially global impact of plant cover change on other ecosystems as communicated by the atmosphere has been under-appreciated and is only beginning to be evaluated [Swann 2018: 2].
Researchers simulated tree die-offs in their model by replacing all trees in a given domain[10] with grass.
Domain-scale tree loss led to changes in local (within same domain) surface properties and fluxes including albedo and evapotranspiration. These changes in surface properties modified local surface climate (e.g., precipitation and temperature), as well as impacted atmospheric circulation. The atmospheric circulation response connects the direct forcing of tree loss on the local atmosphere to other regions, impacting climate and thus resulting in altered Gross Primary Productivity (GPP) across North America [Swann 2018: 3-4].
Furthermore, the severity of the remote effects of tree loss depends not only on the scale of the tree loss, but also on the location of the tree loss. The study found, for example, that tree loss in an area covering most of California had greater effect on GPP in other parts of the continent than did tree loss of a similar scale elsewhere.
Thus, in addition to the magnitude of forest loss, the location of forest loss plays an outsized role in determining the continental scale impact [Swann 2018: 6].
Trees, forests and water: cool insights for a hot world, Ellison 2017[11]
This paper takes the innovative and paradigm-shifting position that carbon is not the primary consideration in climate; rather, water should be the central focus, integrated with carbon and energy cycles:
Forest-driven water and energy cycles are poorly integrated into regional, national, continental and global decision-making on climate change adaptation, mitigation, land use and water management. This constrains humanity’s ability to protect our planet’s climate and life-sustaining functions. The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. The effects of tree cover on climate at local, regional and continental scales offer benefits that demand wider recognition. The forest- and tree-centered research insights we review and analyze provide a knowledge-base for improving plans, policies and actions. Our understanding of how trees and forests influence water, energy and carbon cycles has important implications, both for the structure of planning, management and governance institutions, as well as for how trees and forests might be used to improve sustainability, adaptation and mitigation efforts [Ellison 2017: Abstract].
Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. [Ellison 2017: Abstract].
Biotic pump of atmospheric moisture as driver of the hydrological cycle on land, Makarieva and Gorshkov 2007[12]
The authors examine ecological and geophysical principles to explain how land far inland away from the ocean can remain moist, given that gravity continuously pulls surface and groundwater into the ocean over time.
All freshwater on land originates in the ocean from which it has evaporated, is carried on air flux, and precipitates over the land. Coastal regions benefit from this cycle by their proximity to the ocean, yet in the absence of natural forests in coastal regions precipitation weakens as distance from the ocean increases, leaving inland areas arid. The authors propose the concept of a biotic pump to explain how large continents can be sufficiently moist deep into the interior, and abundant with rivers and lakes.
Air and moisture are pulled horizontally by evapotranspiration over coastal forests. When water vapor from plants condenses, it creates a partial vacuum that pulls water evaporating from the ocean into the continental interior which results in forest rains. By contrast, deserts are unable to pull in ocean evaporation because they lack evaporative force.
Therefore, ongoing deforestation, especially coastal deforestation on a large scale, threatens to cut off rain to the interiors of Earth’s continents, thereby creating new deserts. The Amazonian rainforest is the prime example: Deforestation of the eastern coast of South America has led to changes in the rainforest that is resulting in drying and desertification of the interior, with unprecedented fires and loss of rivers. Historically, Australia’s interior became a desert around the time the first humans arrived on the continent, and the authors speculate that early coastal deforestation was the cause. On the other hand, restoring natural coastal forests can also restore inland water cycles and reverse desertification.
This article illustrates the importance of biological relationships that are ecologically complex and poorly understood. It highlights the significance of the precautionary principle in assessing what we don’t know when altering ecological processes, and taking preventive action in the face of uncertainty.
How Forests Attract Rain: An Examination of a New Hypothesis, Sheil and Murdiyarso 2009
Highlighting the significance of Makarieva and Gorshkov’s “biotic pump” hypothesis (above), Sheil and Murdiyarso explain it in layman’s terms in this article for the benefit of a broader public, and examine its validity. They point out that the biotic pump hypothesis offers an explanation for a question not otherwise resolved in conventional climate theory.
Conventional theory offers no clear explanation for how flat lowlands in continental interiors maintain wet climates. Makarieva and Gorshkov show that if only “conventional mechanisms” (including [rain] recycling) apply, then precipitation should decrease exponentially with distance from the oceans. Researchers have previously puzzled over a missing mechanism to account for observed precipitation patterns (Eltahir 1998) [Sheil & Murdiyarso 2009: 342].
They explain the biotic pump hypothesis and how it resolves the puzzle:
Air currents near Earth’s surface flow to where pressure is lowest. According to Makarieva and Gorshkov, these are the areas that possess the highest evaporation rates. In equatorial climates, forests maintain higher evaporation rates than other cover types, including open water. Thus, forests draw in moist air from elsewhere; the larger the forest area, the greater the volumes of moist air drawn in. This additional moisture rises and condenses in turn, generating a positive feedback in which a large proportion of the water condensing as clouds over wet areas is drawn in from elsewhere. The drivers (solar radiation) and basic thermodynamic concepts and relationships are the same as in conventional models, thus most behaviors are identical— the difference lies in how condensation is incorporated.
Makarieva and Gorshkov’s estimates, incorporating volume changes from condensation, imply that when forest cover is sufficient, enough moist air is drawn in to maintain high rainfall inside continents. The numbers now add up: thus, condensation offers a mechanism to explain why continental precipitation does not invariably decline with distance from the ocean [Sheil & Murdiyarso 2009: 342].
Commenting on the relevance of the hypothesis, the authors conclude:
Acceptance of the biotic pump would add to the values that society places on forest cover. By raising regional concerns about water, acceptance of Makarieva and Gorshkov’s biotic pump demands attention from diverse local actors, including many who may otherwise care little for maintaining forest cover [Sheil & Murdiyarso 2009: 346].
Human modification of global water vapor flows from the land surface, Gordon 2005
Human modification of the hydrological cycle has profoundly affected the flow of liquid water across the Earth’s land surface. Compared to changes to liquid water flow, alteration of water vapor flows through land-use changes has received comparatively less attention, despite compelling evidence that such alteration can influence the functioning of the Earth System.
We show that deforestation is as large a driving force as irrigation in terms of changes in the hydrological cycle. Deforestation has decreased global vapor flows from land by 4% (3,000 km3/yr), a decrease that is quantitatively as large as the increased vapor flow caused by irrigation (2,600 km3/yr). Although the net change in global vapor flows is close to zero, the spatial distributions of deforestation and irrigation are different, leading to major regional transformations of vapor-flow patterns [Gordon 2015: 7612].
A green planet versus a desert world: estimating the maximum effect of vegetation on the land surface climate, Kleidon 2000
This climate model simulation illustrates how the biosphere affects the climate system. With “maximum vegetation,” more water is absorbed in the ground, allowing for evaporation to cool the land surface while also recycling more rain. This simulation resulted in an average temperature reduction over land of 1.2C.
The authors describe their approach:
We quantify the maximum possible influence of vegetation on the global climate by conducting two extreme climate model simulations: in a first simulation (‘desert world’), values representative of a desert are used for the land surface parameters for all non-glaciated land regions. At the other extreme, a second simulation is performed (‘green planet’) in which values are used which are most beneficial for the biosphere’s productivity [Kleidon 2000: 471].
They describe the effects of maximum vegetation on the water cycle, stating that over land:
…the hydrological cycle is more active, with precipitation roughly increasing by 100%, evapotranspiration by more than 200% and the mean moisture content of the atmosphere (or precipitable water) increasing by 30%. These increases can be understood by enhanced recycling of soil water as a response of both, (i) more absorbed radiation at the surface so that more energy is available for evapotranspiration and (ii) larger soil water storage capacities (SWCs) which enhance water availability during dry periods. This increased recycling also leads to an overall decrease in continental runoff by about 25% [Kleidon 2000: 476].
Changes in the water cycle result in land surface temperature changes:
The substantial increase in evapotranspiration is associated with differences in the surface energy balance, primarily concerning the partitioning between sensible and latent heat. The latent heat flux increases by the same amount (more than 200%) as evapotranspiration and the sensible heat flux decreases to 30% of its original value. … Subsequently, the increased latent heat flux leads to more efficient cooling of the surface, resulting in temperatures reduced by 1.2 K[13] [Kleidon 2000: 477-478].
Historical deforestation locally increased the intensity of hot days in northern mid-latitudes, LeJeune 2018
Deforestation contributes to climate change on a global scale through carbon emissions (biogeochemical effects), and on a local/regional scale through biogeophysical effects related to albedo, evapotranspiration and roughness, affecting surface energy budgets.
Here, we show that historical deforestation has led to a substantial local warming of hot days over the northern mid-latitudes – a finding that contrasts with most previous model results. Based on observation-constrained state-of-the-art climate-model experiments, we estimate that moderate reductions in tree cover in these regions have contributed at least one-third of the local present-day warming of the hottest day of the year since pre-industrial time, and were responsible for most of this warming before 1980 [LeJeune 2018: 1].
The study uses observational data to constrain the outcome of a climate model simulating the effects of deforestation on regional temperatures. The authors found that during most of the 20th century, the biogeophysical effects of deforestation were the main cause of regional temperature increases, and that by 1980 deforestation in northern mid-latitudes had declined. By that time other forcings began to take on a proportionally greater role in regional temperature increases.
Twentieth Century regional climate change during the summer in the central United States attributed to agricultural intensification, Alter 2018
Noting that “major increases in crop productivity and changes in regional climate are generally collocated in time and space over the central United States” [Alter 2018: 1587], the study tested the hypothesis that there is a causal relationship – that historical agricultural intensification has affected regional summer climate in this area.
… from 1950 to 2010, the amount of corn harvested annually in the Corn Belt increased by 400%, from 2 billion to 10 billion bushels (National Agricultural Statistics Service, 2016) [Alter 2018: 1586].
and
From 1910 to 1949 (pre-agricultural development, pre-DEV) to 1970–2009 (full agricultural development, full-DEV), the central United States experienced large-scale increases in rainfall of up to 35% and decreases in surface air temperature of up to 1°C during the boreal summer months of July and August, when crop water use in the Corn Belt is at its peak [Alter 2018: 1586].
The authors used a regional climate model to test their hypothesis by comparing a set of simulations where “enhanced photosynthesis over cropland [serves] as a proxy for agricultural intensification” [Alter 2018: 1589] to a control simulation with no agricultural intensification. They found that:
Over the region that has experienced significant increases in observed rainfall (region of significant change—ROSC), the mean rainfall increase is ~7% (0.20mm/d) for the simulations and ~15% (0.37mm/d) for the observations. Thus, it seems that agricultural intensification has been a major contributor to the observed increase in summer rainfall in the central United States [Alter 2018: 1589].
Strikingly, these increases in rainfall are also very consistent: Agricultural intensification enhances simulated rainfall across the aforementioned swath in the central United States during at least 62% of the 150 ensemble years (significant at the 5% level using the chi-square test). In the observational data, a similar consistency in precipitation enhancement is evident when comparing the pre-DEV and full-DEV time periods. This suggests that the changes in rainfall due to agricultural intensification are not the result of occasional increases but instead are indicative of a more systematic change in the summer rainfall regime of the central United States [Alter 2018: 1589].
This study usefully contributes evidence that vegetation cover affects local and regional climates, while drawing conclusions, however, that are not necessarily helpful to understanding how to mitigate and adapt to climate change. The study’s findings suggest that agricultural intensification can potentially mitigate local climate change effects in the future, but it is unlikely that the methods that drove agricultural intensification in the 20th Century will continue to work in a changing climate. The reason that these methods are now obsolete is that they strip the soils of the organic material and living organisms necessary for the resilience of plants, and their ability to cope with droughts, floods, heat and other challenging conditions.
The model here uses “enhanced photosynthesis” as a proxy for agricultural intensification. While the increase in yield between early and late 20th Century Corn Belt production represents an increase in photosynthesis, high-input agriculture is but one pathway to enhanced photosynthesis. Moreover, it is an extremely problematicone with respect to climate change, given the high energy costs of fertilizer, pesticides and fuel, and the damage to the soils from these practices.
Instead, a useful lesson to draw from this study is simply that enhanced photosynthesis itself can mitigate climate change regionally. In the context of agricultural production in the era of climate change, enhanced photosynthesis might best be accomplished through ecological intensification, a strategy for improving resilience within an agro-ecosystem, and thereby greater photosynthesis and more reliable crop production.
Intermediate tree cover can maximize groundwater recharge in the seasonally dry tropics, Ilstedt 2016
Responding to a common belief that trees lower groundwater infiltration due to transpiration, and a contrasting view that trees increase groundwater infiltration by increasing organic matter and soil porosity, these authors test an “optimum tree cover theory.”
They find that “intermediate” tree cover maximizes groundwater recharge in the tropics, resulting in a 2-14% increase in total annual water input from rainfall. However, the tree species used in this study consume more water compared to many other tree species in the semi-arid tropics. Therefore, the results here may be conservative in terms of the potential of trees to increase groundwater recharge. Furthermore, the study doesn’t consider the potential effects of greater transpiration from increased tree cover on local rainfall patterns.
Critical impact of vegetation physiology on the continental hydrologic cycle in response to increasing CO2, Lemordant 2018
This study finds that the physiological response of plants to increased atmospheric CO2 affects the global hydrological cycle even more than does the greenhouse effect and changes in precipitation. The authors conclude:
This highlights the key role of vegetation in controlling future terrestrial hydrologic response and emphasizes that the carbon and water cycles are intimately coupled over land [LeMordant 2018: 1].
With increasing [CO2] at the leaf surface, the density of stomata at the leaf surface is decreased and their individual opening is reduced and therefore less water is transpired per unit leaf area. In other words, leaf-level water use efficiency increases, potentially increasing surface soil moisture and runoff. On the other hand, leaf biomass tends to also increase with increasing [CO2] … generating a larger evaporative surface that can partly offset the reduction in stomatal conductance and negate the soil water savings. Our objective is therefore to quantify how such plant [CO2] effects influence future hydrological variable responses compared with radiative effects ––the atmospheric impact of the “greenhouse effect.” Radiative effects impact precipitation, i.e., water supply, and evaporative demand, through increase in radiation, temperature, and atmospheric dryness as estimated by the vapor pressure deficit (VPD), i.e., saturation minus actual vapor pressure [LeMordant 2018: 1].
Our study illustrates how deeply the physiological effects [on vegetation] due to increasing atmospheric [CO2] impact the continental water cycle. Contrary to previous wisdom, changes in precipitation and radiation [greenhouse effect] do not play the primary role in future drying and moistening in most regions. Rather, biosphere physiological effects and related biosphere–atmosphere interactions are key for predicting future continental water stress as represented by ET [evapotranspiration], long-term runoff, EF, or leaf area index. In turn, vegetation water stress largely regulates land carbon uptake, further emphasizing how tightly the future carbon and water cycles are coupled so that they cannot be evaluated in isolation [LeMordant 2018: 5].
Weakening of Indian summer monsoon rainfall due to changes in land use land cover, Paul 2016
The Indian summer monsoon rainfall has decreased since 1950, and several hypotheses have been proposed to explain why. Most of these hypotheses involving weakening temperature gradients over the continent. This study explores the potential link between a weakening monsoon and widespread land use land cover (LULC) change from woody savanna to cropland in recent decades. Citing earlier studies, the authors note that:
Precipitation resulting from local land surface Evapotranspiration (ET) is known as recycled precipitation. Pathak et al. found that evapotranspiration from land surface vegetation plays a major role during the end of a monsoon. They observed that, during the initial phase of a monsoon, oceanic sources play a major role, and the soil is recharged with moisture. However, during the latter half of a summer monsoon (August and September), land surface ET increases as recycled precipitation increases, a pattern that is more prominent in the Ganga Basin and Northeast India. This recycled precipitation accounts for approximately 20–25% of the rainfall in North India (Ganga Basin) and Northeast India during August and September. Hence, deforestation associated with changes in LULC may affect ET and may significantly affect monsoon rainfall [Paul 2016: 1-2].
Summarizing their own study, the authors conclude:
Here, we performed a sensitivity analysis to quantify the impacts of large-scale conversion from woody savannah to crop land in India on monsoon precipitation. We found such a change results in decreased ET and subsequently a decrease in recycled precipitation leading to a decline in monsoon precipitation. This decline is similar in extent to the observed recent decadal weakening of monsoon precipitation. However, other reasons may account for this observed weakening, such as the warming of Indian Ocean SST [sea surface temperature] [Paul 2016: 5-6].
To better clarify causality of the weakening monsoon, given other potential factors, the authors propose that:
The future scope of this present work is to perform detection and attribution studies for potential declines of Indian monsoons with model runs forced with SST warming only, aerosol forcing only, LULC changes only and all controlling factors together [Paul 2016: 6].
Tropical reforestation and climate change: beyond carbon, Locatelli 2015
When managed with both climate adaptation and mitigation in mind, tropical reforestation (TR) can serve multiple synergistic functions. TR mitigates regional and global climate change, not only by sequestering carbon but also through biophysical cooling (via evapotranspiration), by recycling rainfall regionally, and by reducing pressure on old growth forests.
Furthermore, TR helps local communities adapt to climate change by recharging stream flow in the dry season, reducing the severity of floods, protecting slopes against landslides and, through mangrove reforestation, reducing the impact of coastal storms and waves. Reforestation also creates livelihood opportunities through the sustainable harvest of forest products, and creates shelter and habitat for species vulnerable to climate change. However, to achieve this broad range of benefits, “reforestation practices should be designed to avoid the implementation of one strategy (mitigation or adaptation) to the detriment of the other.” Arguing for the application of what they term “climate-smart reforestation,” the authors recommend the following:
The challenge for climate-smart reforestation is to implement an effective combination of approaches to meet all three objectives: societal adaptation, climate mitigation, and ecological resilience [Locatelli 2015: 4].
However, as most policies consider the three objectives of climate-smart reforestation separately, they often overlook possible trade-offs and synergies. For example,reforestation projects managed with a carbon purpose could have detrimental consequences on water availability in the semi-arid tropics (Trabucco et al. 2008) or on biodiversity (O’Connor 2008). By contrast, reforestation that is explicitly climate-smart uses a multi-objective planning focus that enables different objectives to reinforce each other so that their interactions produce synergies rather than trade-offs. For example, tree regeneration in Tanzania under the Ngitili resource management system achieves carbon storage together with improved watershed conservation and greater provision of natural resources (water, food, and fodder) for livelihoods (Duguma et al. 2014). A proposed adaptation project in Colombia aims to reforest with flood-resistant native tree species to reduce flood impacts on downstream communities (UNDP 2012). A project in Costa Rica is testing different mixes of species and silvicultural practices to reduce vulnerability to storms and fires while also achieving carbon storage (Locatelli et al. 2011) [Locatelli 2015: 4-5].
This article underscores a key concept of this compendium – that functioning ecosystems (whether old growth or restored forests, for example) provide multiple, interwoven functions that support human and biodiverse life by regulating local, regional and global climate conditions.
Water-retention potential of Europe’s forests: A European overview to support natural water-retention measures, European Environment Agency (EEA) 2015
The importance of water retention (the rainfall absorbed or used within an ecosystem) for mitigating flood and drought conditions and contributing to clean drinking water, for example, has been increasingly recognized in Europe in the past decade. Along with wetland preservation, better agriculture practices and other measures, preserving and re-growing forests are seen as key to enhanced natural water retention. Forests cover a third of Europe, and:
can soak up excess rainwater, preventing run-offs and damage from flooding. By releasing water in the dry season, forests can help to provide clean water and mitigate the effects of droughts [EEA 2015: 6].
In recognition of the important water management role of forests and other natural ecosystems, new policy instruments have proposed Natural Water-Retention Measures (NWRMs).
Natural Water-Retention Measures (NWRMs) are defined as ‘measures to protect and manage water resources and to address water-related challenges by restoring or maintaining ecosystems, natural features and characteristics of water bodies using natural means and processes’ (European Commission and Directorate-General for the Environment 2014). … The main focus is to enhance and preserve the water retention capacity of aquifers, soil and ecosystems and improve their status [EEA 2015: 9].
This EEA study found that:
In water-basins where the forest cover is 30%, water retention is 25% higher than in basins where the forest cover is only 10%. In basins where the forest cover is 70%, water retention is 50% higher than in basins where the forest cover is only 10%. … Coniferous forests in general retain 10% more water than broadleaved forests or mixed forests [EEA 2015: 5].
Why Climate Change Makes Riparian Restoration More Important than Ever: Recommendations for Practice and Research, Seavy 2009
Riparian[14] ecosystems are naturally resilient, provide linear habitat connectivity, link aquatic and terrestrial ecosystems, and create thermal refugia for wildlife: all characteristics that can contribute to ecological adaptation to climate change [Seavy 2009: 330].
Arguing for the restoration of riparian areas because of their ecological significance and inherent resilience, these authors articulate the importance of both surface and groundwater – protected within a biodiverse ecosystem – for its cooling effect.
Because riparian areas have higher water content than surrounding upland areas, they absorb heat and buffer organisms against extreme temperatures (Naiman et al. 2000). During previous periods of climate change, riparian areas served as refugia because they provided microclimates that protected plant biodiversity (Bakker 1984, Meave and Kellman 1994). Riparian vegetation can maintain cooler water temperatures by shading water from sunlight (Sridhar et al. 2004, Cassie 2006) and the infusion of cold groundwater into warmer surface waters creates and maintains pockets of cool water (Chu et al. 2008). Thus, riparian areas provide thermal refugia for animals with thermoregulatory limitations [Seavy 2009: 332].
Water and vegetation are climate heroes, co-starring in a story about as old as terrestrial life on Earth yet under-recognized in mainstream climate politics. Not only does the vegetation embedded in ecosystems act as a giant CO2-absorption machine, constantly removing the greenhouse gas from the air and storing much of it in soil and biomass, but vegetation also tames the energetic flow of liquid and gaseous water around the planet, mitigating drought and flood conditions. Plants facilitate the recharge of groundwater, while also recharging the skies with moisture for rain. And through this plant-water partnership, vegetation also cools the Earth.
Water, thanks to its high heat-carrying capacity, is able to redistribute much of the solar heat energy received by the Earth through the water cycle: through evapotranspiration and condensation. Thus the evapotranspiration and condensation of water plays an instrumental role in climate control with regard to temperature distribution in time and space. That is, it helps reduce the peaks and modulate the amplitudes of high and low temperatures on the land surface – making conditions on Earth suitable for life [Eiseltova 2012: 306].
Like all living things, plants need water to survive. Yet, the average plant uses less than 5% of the water taken up by its roots for its own cell production and growth [McElrone 2013]. The remainder is essentially a coolant in a plant’s private air conditioning system. Through the process of transpiration, water absorbed by roots and transported up the stem eventually makes it to the stomatal openings on the leaves, vaporizing just below the surface and cooling the plant in a release of latent heat.
The average plant uses less than 5% of the water taken up by its roots for its own cell production and growth [McElrone 2013]. The remainder is essentially a coolant in a plant’s private air conditioning system.
Latent heat is the energy absorbed by water that transforms liquid into vapor. The water in a leaf intercepting sunlight absorbs thermal energy from the sun and then releases it in vapor in a process that involves no temperature change. Thus, living plants are never hot to the touch. By contrast, solar energy alighting on a mineral surface is absorbed as sensible heat[3], raising the temperature of that surface.
An average tree can transpire hundreds of liters of water per day. Every 100 liters of water transpired equates in cooling power to the daily output of two central air-conditioning units for an average home [Ellison 2017:54]. Multiplied millions of times over a given landscape, the cooling effect of transpiration is significant.
Imagine yourself in a city, where on a hot day the asphalt is too hot even to touch, and the heat radiating from it permeates everything around it.[4] Next, imagine yourself on that same hot day walking into a city park full of trees, bushes and grasses: not only can you take your shoes off on the cool grass and park yourself in some shade, but with enough trees around you can also feel a distinct coolness in the air even out of the shade.
Similarly, vegetation cover has been shown to affect local, regional and continental climates [LeJeune 2018, Eiseltova 2012, Locatelli 2015, Alter 2018, Paul 2016, Ellison 2017, Swann 2018, Makarieva 2007]. As noted, vegetation contributes to rain through transpiration, which augments atmospheric vapor flows, and through the release of biotic aerosols, which coalesce water droplets in clouds into drops of rain [Hassett 2017, Ellison 2017]. Furthermore, by increasing groundwater retention,,vegetation mitigates local flood and drought conditions and filters/cleans drinking water [EEA 2015]. The cooling action of forests also creates sanctuary from the intensifying heat of climate change for animals [Betts 2018] and people alike.
Less obvious perhaps, though equally significant, is that vegetation cover also regulates temperatures and climate on a global scale [Ellison 2017, Lemordant 2018, Gordon 2005, Kleidon 2000, Locatelli 2015], thus influencing weather patterns on Earth. For example, Gordon [2005] notes that deforestation has reduced global vapor flows by more than 4%, changing rainfall distribution patterns. Kleidon [2000] shows an average global temperature difference of 1.2C between hypothetical conditions of maximum and minimum global vegetation, regardless of any greenhouse gas effect. (For comparison, 1.2C is roughly the level of global warming since the start of the Industrial Revolution due to land use change and the greenhouse gas effect.) Ellison [2017: 52] notes that land cover change accounts for some 18% of global warming trends.
Further evidence of vegetation’s significant effect on the global climate is Lemordant’s [2018] finding that the physiological response of plants to increased atmospheric CO2 affects the global hydrological cycle even more than do the greenhouse effect and changes in precipitation. That’s because plants shrink their stomata to limit CO2 intake and consequently limit the release of water vapor from those same stomata. Stomatal shrinkage results in more water left in the soil and less water entering the atmosphere through transpiration.
In summary, Ellison et al [2017] state that,
The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles [Ellison 2017: 51].
In their prescriptions for land management policies, both Locatelli [2015] and Eiseltova [2012] embrace the concept that vegetation (and forests in particular) exert control in diverse and dynamic ways on local, regional and global climate systems. Locatelli argues for “climate-smart” tropical reforestation that not only enhancesthe carbon sequestration process, but also helps local communities adapt to climate change by recharging stream flow in the dry season, reducing the severity of floods, protecting slopes against landslides and, through mangrove reforestation, reducing the impact of coastal storms and waves.
Similarly, Eiseltova et al [2012: 324] argue that “there is an urgent need that agricultural research focuses on how to close water cycles in the landscape and the development of farming systems with a more vertically-layered vegetation structure keeping water and lower temperatures during a sunny day.” These authors propose “two criteria for assessing sustainable land management”:
These criteria are: the efficiency of an ecosystem to recycle water and matter; and its efficiency to dissipate solar energy. It is land managers that can substantially contribute to the restoration of the water cycle, climate amelioration and reduction of irreversible matter losses with river water flows to the sea [Eiseltova 2012: 325].
The collection of article summaries that follow reveals a cascade of recent discoveries about the relevance of forests and other vegetative ecosystems, vis a vis regional and global water cycles,in discussions of climate mitigation and adaptation. What is both fascinating and critical here is, as Locatelli et al allude to, the multi-functionality of extensive, integral vegetative ecosystems with respect to climate change. This multifunctionality is due to the roles vegetation plays both locally, in adapting to temperature and weather extremes and contributing to forestry/farming-related livelihood opportunities, and globally, through CO2 sequestration, global average temperature reduction, and evenness of rainfall distribution.
What is both fascinating and critical here is, as Locatelli et al. allude to, the multi-functionality of extensive, integral vegetative ecosystems with respect to climate change.
Water Article Summaries
Evapotranspiration – A Driving Force in Landscape Sustainability, Eiseltová 2012
Vegetation cover cools Earth when it intercepts the sun’s energy. This is not just by providing shade, but also through evapotranspiration, which is how plants regulate their own internal temperatures.
For a plant … transpiration[5] is a necessity by which a plant maintains its inner environment within the limit of optimal temperatures. And at the level of landscape, evapotranspiration is the most efficient air conditioning system developed by nature [Eiseltova 2012:10].
The water in plant tissues contains the sun’s energy in the form of latent heat, which is released from plants through evapotranspiration. In the absence of water, solar energy reaching Earth becomes sensible heat – the heat we can feel and measure in rising temperatures.
Without water, the energy of the incoming radiation is transformed into sensible heat and the local area becomes overheated during the day and likewise far cooler at night (as is well known from desert areas, with differences between day and night temperatures typically exceeding 50°C). Water-saturated landscapes provide much more stable environments than do dry terrestrial systems. In landscapes with water – abundant aquatic ecosystems, wetlands and soils with high water retention capacity – about 80% of incoming solar energy is stored as latent heat of water vapour via evapotranspiration, whilst in de-watered landscapes (with a low-water retention capacity) the vast majority of solar energy is transformed into sensible heat (Pokorný et al. 2010b) [Eiseltova 2012: 307].
With respect to landscape management for sustainability, the authors introduce the idea of a “dissipative-ecological-unit,” meaning “the smallest functional unit that is capable of forming internalized cycles of matter and water while dissipating energy” [Eiseltova 2012: 312]. This term emphasizes the importance of small, local water cycles, which occur naturally in undisturbed ecosystems, resulting in “an efficient local resource economy and … relatively even temperatures and moisture conditions” [Eiseltova 2012: 312].
In catchments with a well-developed vegetation cover, water and matter are bound to short-circuited cycles and losses are minimal. In contrast, the increased clearance of forest, exposure of bare land, and drainage of agricultural land have accelerated matter losses from catchments [Eiseltova 2012: 11].
There is an urgent need that agricultural research focuses on how to close water cycles[6] in the landscape and the development of farming systems with a more vertically-layered vegetation structure keeping water and lower temperatures during a sunny day[7] [Eiseltova 2012: 324].
The water cycle is akin to the ‘bloodstream’ of the biosphere. Returning water to the landscape and restoring more natural vegetation cover is the only way to restore landscape sustainability. More attention in present-day science needs to be devoted to the study of the role of vegetation in the water cycle and climate amelioration. Restoration of a more natural vegetation cover over the landscape seems to be the only way forward.
Based on our current scientific knowledge, we can propose two criteria for assessing sustainable land management. These criteria are: the efficiency of an ecosystem to recycle water and matter,and its efficiency to dissipate solar energy. Land managers can substantially contribute to the restoration of the water cycle, climate amelioration and reduction of irreversible matter [soil and nutrient] losses with river water flows to the sea.
It is in the interest of society as a whole that land managers (farmers,foresters) be rewarded for their actions towards sustainable management of their land. Suitable tools to assess the achievements of individual land managers with respect to sustainable management of their land are: (1) continuous monitoring of conductivity – a measure of dissolved load – and flow rates in streams in order to estimate matter losses; and (2) the regular evaluation of satellite thermal channel images to assess temperature damping, i.e., the effectiveness of land use to dissipate solar energy. Restoration of natural ‘cooling structures’ – vegetation with its evapotranspiration and condensation-induced water circulation – is essential to renew landscape sustainability [Eiseltova 2012: 325].
New climate solutions, water cycles and the soil carbon sponge, Jehne 2018
Regenerating the soil carbon sponge is our greatest point of leverage for salvaging the planet from the point of existential climate crisis. “Sponge” refers to the quality of a biologically active soil with high organic matter content to have lots of pore space for water absorption. Jehne states that every additional gram of soil carbon allows the soil to hold 8 additional grams of water. He emphasizes the soil sponge concept because it is the driver of healthy ecosystems, and also within our control to repair and regenerate.
Jehne explains that an average of 342 W/m2 of incident solar radiation enters the troposphere while just 339 W/m2 is reflected back into space due to the greenhouse effect [Jehne 2018: 19:00 min]. This leaves a continuous energy balance of 3 W/m2 heating up the planet. Of the solar radiation returning to space, 24% is released through latent heat fluxes from evapotranspiration [Jehne 2018: 1:34:15]. However, due to land use change, there is 50% less transpiration on Earth than there was some 8,000 years ago. Jehne estimates that increasing transpiration by only 5% would be enough to offset the 3 W/m2 surplus solar energy [Jehne 2018: 1:34:50].
Increasing transpiration is achieved by increasing vegetation cover, which in turn is achieved by regenerating the soil sponge. Jehne explains that conventional agriculture has employed techniques (such as burning, cultivating/tilling, applying fertilizer and pesticides, and use of irrigation and fallow) that quickly oxidize the carbon fixed by plants through photosynthesis. By contrast, regenerative agriculture builds up the soil carbon sponge by facilitating the ecological processes that create stable soil carbon and limit organic matter breakdown.
In addition to the cooling effect from the latent heat flux, transpiration also provides the moisture needed for cloud formation. Jehne states that a 2% increase in cloud cover, given its high albedo, is also enough to reflect the excess 3 W/m2 solar radiation that is otherwise absorbed on Earth [Jehne 2018: 1:39:25]. Furthermore, bacteria released from ecosystems serve as the most effective precipitation nuclei[8] for making rain.
Continental-scale consequences of tree die-offs in North America: identifying where forest loss matters most, Swann 2018
Vegetation cover affects the amount of solar energy a land area absorbs and/or releases, thus altering local temperatures and precipitation. Plants regulate local temperatures through shading, albedo and evapotranspiration, which releases latent[9] heat.
The ability of a surface to shed energy through latent or sensible heat is key to determining that surface’s temperature – shifts in the relative balance between the two can lead to increases in surface temperatures (where sensible heat is relatively higher) or decreases (where latent heat is relatively higher) [Swann 2018: 2].
This study shows that changes in vegetation cover in a given place affect not only the local climate, but also the climate system at a continental scale. The results are temperature and precipitation changes in remote parts of the continent relative to where the tree loss occurred, leading to changes in ecosystem productivity in those remote parts. This phenomenon is called ‘ecoclimate teleconnections.’
Plants profoundly influence local climate by controlling the exchange of energy and water with the atmosphere. Changes in and/or losses of plant type or plant functioning can alterthe local climate, but also potentially large scale climate by modifying atmospheric circulation. … the potentially global impact of plant cover change on other ecosystems as communicated by the atmosphere has been under-appreciated and is only beginning to be evaluated [Swann 2018: 2].
Researchers simulated tree die-offs in their model by replacing all trees in a given domain[10] with grass.
Domain-scale tree loss led to changes in local (within same domain) surface properties and fluxes including albedo and evapotranspiration. These changes in surface properties modified local surface climate (e.g., precipitation and temperature), as well as impacted atmospheric circulation. The atmospheric circulation response connects the direct forcing of tree loss on the local atmosphere to other regions, impacting climate and thus resulting in altered Gross Primary Productivity (GPP) across North America [Swann 2018: 3-4].
Furthermore, the severity of the remote effects of tree loss depends not only on the scale of the tree loss, but also on the location of the tree loss. The study found, for example, that tree loss in an area covering most of California had greater effect on GPP in other parts of the continent than did tree loss of a similar scale elsewhere.
Thus, in addition to the magnitude of forest loss, the location of forest loss plays an outsized role in determining the continental scale impact [Swann 2018: 6].
Trees, forests and water: cool insights for a hot world, Ellison 2017[11]
This paper takes the innovative and paradigm-shifting position that carbon is not the primary consideration in climate; rather, water should be the central focus, integrated with carbon and energy cycles:
Forest-driven water and energy cycles are poorly integrated into regional, national, continental and global decision-making on climate change adaptation, mitigation, land use and water management. This constrains humanity’s ability to protect our planet’s climate and life-sustaining functions. The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. The effects of tree cover on climate at local, regional and continental scales offer benefits that demand wider recognition. The forest- and tree-centered research insights we review and analyze provide a knowledge-base for improving plans, policies and actions. Our understanding of how trees and forests influence water, energy and carbon cycles has important implications, both for the structure of planning, management and governance institutions, as well as for how trees and forests might be used to improve sustainability, adaptation and mitigation efforts [Ellison 2017: Abstract].
Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. [Ellison 2017: Abstract].
Biotic pump of atmospheric moisture as driver of the hydrological cycle on land, Makarieva and Gorshkov 2007[12]
The authors examine ecological and geophysical principles to explain how land far inland away from the ocean can remain moist, given that gravity continuously pulls surface and groundwater into the ocean over time.
All freshwater on land originates in the ocean from which it has evaporated, is carried on air flux, and precipitates over the land. Coastal regions benefit from this cycle by their proximity to the ocean, yet in the absence of natural forests in coastal regions precipitation weakens as distance from the ocean increases, leaving inland areas arid. The authors propose the concept of a biotic pump to explain how large continents can be sufficiently moist deep into the interior, and abundant with rivers and lakes.
Air and moisture are pulled horizontally by evapotranspiration over coastal forests. When water vapor from plants condenses, it creates a partial vacuum that pulls water evaporating from the ocean into the continental interior which results in forest rains. By contrast, deserts are unable to pull in ocean evaporation because they lack evaporative force.
Therefore, ongoing deforestation, especially coastal deforestation on a large scale, threatens to cut off rain to the interiors of Earth’s continents, thereby creating new deserts. The Amazonian rainforest is the prime example: Deforestation of the eastern coast of South America has led to changes in the rainforest that is resulting in drying and desertification of the interior, with unprecedented fires and loss of rivers. Historically, Australia’s interior became a desert around the time the first humans arrived on the continent, and the authors speculate that early coastal deforestation was the cause. On the other hand, restoring natural coastal forests can also restore inland water cycles and reverse desertification.
This article illustrates the importance of biological relationships that are ecologically complex and poorly understood. It highlights the significance of the precautionary principle in assessing what we don’t know when altering ecological processes, and taking preventive action in the face of uncertainty.
How Forests Attract Rain: An Examination of a New Hypothesis, Sheil and Murdiyarso 2009
Highlighting the significance of Makarieva and Gorshkov’s “biotic pump” hypothesis (above), Sheil and Murdiyarso explain it in layman’s terms in this article for the benefit of a broader public, and examine its validity. They point out that the biotic pump hypothesis offers an explanation for a question not otherwise resolved in conventional climate theory.
Conventional theory offers no clear explanation for how flat lowlands in continental interiors maintain wet climates. Makarieva and Gorshkov show that if only “conventional mechanisms” (including [rain] recycling) apply, then precipitation should decrease exponentially with distance from the oceans. Researchers have previously puzzled over a missing mechanism to account for observed precipitation patterns (Eltahir 1998) [Sheil & Murdiyarso 2009: 342].
They explain the biotic pump hypothesis and how it resolves the puzzle:
Air currents near Earth’s surface flow to where pressure is lowest. According to Makarieva and Gorshkov, these are the areas that possess the highest evaporation rates. In equatorial climates, forests maintain higher evaporation rates than other cover types, including open water. Thus, forests draw in moist air from elsewhere; the larger the forest area, the greater the volumes of moist air drawn in. This additional moisture rises and condenses in turn, generating a positive feedback in which a large proportion of the water condensing as clouds over wet areas is drawn in from elsewhere. The drivers (solar radiation) and basic thermodynamic concepts and relationships are the same as in conventional models, thus most behaviors are identical— the difference lies in how condensation is incorporated.
Makarieva and Gorshkov’s estimates, incorporating volume changes from condensation, imply that when forest cover is sufficient, enough moist air is drawn in to maintain high rainfall inside continents. The numbers now add up: thus, condensation offers a mechanism to explain why continental precipitation does not invariably decline with distance from the ocean [Sheil & Murdiyarso 2009: 342].
Commenting on the relevance of the hypothesis, the authors conclude:
Acceptance of the biotic pump would add to the values that society places on forest cover. By raising regional concerns about water, acceptance of Makarieva and Gorshkov’s biotic pump demands attention from diverse local actors, including many who may otherwise care little for maintaining forest cover [Sheil & Murdiyarso 2009: 346].
Human modification of global water vapor flows from the land surface, Gordon 2005
Human modification of the hydrological cycle has profoundly affected the flow of liquid water across the Earth’s land surface. Compared to changes to liquid water flow, alteration of water vapor flows through land-use changes has received comparatively less attention, despite compelling evidence that such alteration can influence the functioning of the Earth System.
We show that deforestation is as large a driving force as irrigation in terms of changes in the hydrological cycle. Deforestation has decreased global vapor flows from land by 4% (3,000 km3/yr), a decrease that is quantitatively as large as the increased vapor flow caused by irrigation (2,600 km3/yr). Although the net change in global vapor flows is close to zero, the spatial distributions of deforestation and irrigation are different, leading to major regional transformations of vapor-flow patterns [Gordon 2015: 7612].
A green planet versus a desert world: estimating the maximum effect of vegetation on the land surface climate, Kleidon 2000
This climate model simulation illustrates how the biosphere affects the climate system. With “maximum vegetation,” more water is absorbed in the ground, allowing for evaporation to cool the land surface while also recycling more rain. This simulation resulted in an average temperature reduction over land of 1.2C.
The authors describe their approach:
We quantify the maximum possible influence of vegetation on the global climate by conducting two extreme climate model simulations: in a first simulation (‘desert world’), values representative of a desert are used for the land surface parameters for all non-glaciated land regions. At the other extreme, a second simulation is performed (‘green planet’) in which values are used which are most beneficial for the biosphere’s productivity [Kleidon 2000: 471].
They describe the effects of maximum vegetation on the water cycle, stating that over land:
…the hydrological cycle is more active, with precipitation roughly increasing by 100%, evapotranspiration by more than 200% and the mean moisture content of the atmosphere (or precipitable water) increasing by 30%. These increases can be understood by enhanced recycling of soil water as a response of both, (i) more absorbed radiation at the surface so that more energy is available for evapotranspiration and (ii) larger soil water storage capacities (SWCs) which enhance water availability during dry periods. This increased recycling also leads to an overall decrease in continental runoff by about 25% [Kleidon 2000: 476].
Changes in the water cycle result in land surface temperature changes:
The substantial increase in evapotranspiration is associated with differences in the surface energy balance, primarily concerning the partitioning between sensible and latent heat. The latent heat flux increases by the same amount (more than 200%) as evapotranspiration and the sensible heat flux decreases to 30% of its original value. … Subsequently, the increased latent heat flux leads to more efficient cooling of the surface, resulting in temperatures reduced by 1.2 K[13] [Kleidon 2000: 477-478].
Historical deforestation locally increased the intensity of hot days in northern mid-latitudes, LeJeune 2018
Deforestation contributes to climate change on a global scale through carbon emissions (biogeochemical effects), and on a local/regional scale through biogeophysical effects related to albedo, evapotranspiration and roughness, affecting surface energy budgets.
Here, we show that historical deforestation has led to a substantial local warming of hot days over the northern mid-latitudes – a finding that contrasts with most previous model results. Based on observation-constrained state-of-the-art climate-model experiments, we estimate that moderate reductions in tree cover in these regions have contributed at least one-third of the local present-day warming of the hottest day of the year since pre-industrial time, and were responsible for most of this warming before 1980 [LeJeune 2018: 1].
The study uses observational data to constrain the outcome of a climate model simulating the effects of deforestation on regional temperatures. The authors found that during most of the 20th century, the biogeophysical effects of deforestation were the main cause of regional temperature increases, and that by 1980 deforestation in northern mid-latitudes had declined. By that time other forcings began to take on a proportionally greater role in regional temperature increases.
Twentieth Century regional climate change during the summer in the central United States attributed to agricultural intensification, Alter 2018
Noting that “major increases in crop productivity and changes in regional climate are generally collocated in time and space over the central United States” [Alter 2018: 1587], the study tested the hypothesis that there is a causal relationship – that historical agricultural intensification has affected regional summer climate in this area.
… from 1950 to 2010, the amount of corn harvested annually in the Corn Belt increased by 400%, from 2 billion to 10 billion bushels (National Agricultural Statistics Service, 2016) [Alter 2018: 1586].
and
From 1910 to 1949 (pre-agricultural development, pre-DEV) to 1970–2009 (full agricultural development, full-DEV), the central United States experienced large-scale increases in rainfall of up to 35% and decreases in surface air temperature of up to 1°C during the boreal summer months of July and August, when crop water use in the Corn Belt is at its peak [Alter 2018: 1586].
The authors used a regional climate model to test their hypothesis by comparing a set of simulations where “enhanced photosynthesis over cropland [serves] as a proxy for agricultural intensification” [Alter 2018: 1589] to a control simulation with no agricultural intensification. They found that:
Over the region that has experienced significant increases in observed rainfall (region of significant change—ROSC), the mean rainfall increase is ~7% (0.20mm/d) for the simulations and ~15% (0.37mm/d) for the observations. Thus, it seems that agricultural intensification has been a major contributor to the observed increase in summer rainfall in the central United States [Alter 2018: 1589].
Strikingly, these increases in rainfall are also very consistent: Agricultural intensification enhances simulated rainfall across the aforementioned swath in the central United States during at least 62% of the 150 ensemble years (significant at the 5% level using the chi-square test). In the observational data, a similar consistency in precipitation enhancement is evident when comparing the pre-DEV and full-DEV time periods. This suggests that the changes in rainfall due to agricultural intensification are not the result of occasional increases but instead are indicative of a more systematic change in the summer rainfall regime of the central United States [Alter 2018: 1589].
This study usefully contributes evidence that vegetation cover affects local and regional climates, while drawing conclusions, however, that are not necessarily helpful to understanding how to mitigate and adapt to climate change. The study’s findings suggest that agricultural intensification can potentially mitigate local climate change effects in the future, but it is unlikely that the methods that drove agricultural intensification in the 20th Century will continue to work in a changing climate. The reason that these methods are now obsolete is that they strip the soils of the organic material and living organisms necessary for the resilience of plants, and their ability to cope with droughts, floods, heat and other challenging conditions.
The model here uses “enhanced photosynthesis” as a proxy for agricultural intensification. While the increase in yield between early and late 20th Century Corn Belt production represents an increase in photosynthesis, high-input agriculture is but one pathway to enhanced photosynthesis. Moreover, it is an extremely problematicone with respect to climate change, given the high energy costs of fertilizer, pesticides and fuel, and the damage to the soils from these practices.
Instead, a useful lesson to draw from this study is simply that enhanced photosynthesis itself can mitigate climate change regionally. In the context of agricultural production in the era of climate change, enhanced photosynthesis might best be accomplished through ecological intensification, a strategy for improving resilience within an agro-ecosystem, and thereby greater photosynthesis and more reliable crop production.
Intermediate tree cover can maximize groundwater recharge in the seasonally dry tropics, Ilstedt 2016
Responding to a common belief that trees lower groundwater infiltration due to transpiration, and a contrasting view that trees increase groundwater infiltration by increasing organic matter and soil porosity, these authors test an “optimum tree cover theory.”
They find that “intermediate” tree cover maximizes groundwater recharge in the tropics, resulting in a 2-14% increase in total annual water input from rainfall. However, the tree species used in this study consume more water compared to many other tree species in the semi-arid tropics. Therefore, the results here may be conservative in terms of the potential of trees to increase groundwater recharge. Furthermore, the study doesn’t consider the potential effects of greater transpiration from increased tree cover on local rainfall patterns.
Critical impact of vegetation physiology on the continental hydrologic cycle in response to increasing CO2, Lemordant 2018
This study finds that the physiological response of plants to increased atmospheric CO2 affects the global hydrological cycle even more than does the greenhouse effect and changes in precipitation. The authors conclude:
This highlights the key role of vegetation in controlling future terrestrial hydrologic response and emphasizes that the carbon and water cycles are intimately coupled over land [LeMordant 2018: 1].
With increasing [CO2] at the leaf surface, the density of stomata at the leaf surface is decreased and their individual opening is reduced and therefore less water is transpired per unit leaf area. In other words, leaf-level water use efficiency increases, potentially increasing surface soil moisture and runoff. On the other hand, leaf biomass tends to also increase with increasing [CO2] … generating a larger evaporative surface that can partly offset the reduction in stomatal conductance and negate the soil water savings. Our objective is therefore to quantify how such plant [CO2] effects influence future hydrological variable responses compared with radiative effects ––the atmospheric impact of the “greenhouse effect.” Radiative effects impact precipitation, i.e., water supply, and evaporative demand, through increase in radiation, temperature, and atmospheric dryness as estimated by the vapor pressure deficit (VPD), i.e., saturation minus actual vapor pressure [LeMordant 2018: 1].
Our study illustrates how deeply the physiological effects [on vegetation] due to increasing atmospheric [CO2] impact the continental water cycle. Contrary to previous wisdom, changes in precipitation and radiation [greenhouse effect] do not play the primary role in future drying and moistening in most regions. Rather, biosphere physiological effects and related biosphere–atmosphere interactions are key for predicting future continental water stress as represented by ET [evapotranspiration], long-term runoff, EF, or leaf area index. In turn, vegetation water stress largely regulates land carbon uptake, further emphasizing how tightly the future carbon and water cycles are coupled so that they cannot be evaluated in isolation [LeMordant 2018: 5].
Weakening of Indian summer monsoon rainfall due to changes in land use land cover, Paul 2016
The Indian summer monsoon rainfall has decreased since 1950, and several hypotheses have been proposed to explain why. Most of these hypotheses involving weakening temperature gradients over the continent. This study explores the potential link between a weakening monsoon and widespread land use land cover (LULC) change from woody savanna to cropland in recent decades. Citing earlier studies, the authors note that:
Precipitation resulting from local land surface Evapotranspiration (ET) is known as recycled precipitation. Pathak et al. found that evapotranspiration from land surface vegetation plays a major role during the end of a monsoon. They observed that, during the initial phase of a monsoon, oceanic sources play a major role, and the soil is recharged with moisture. However, during the latter half of a summer monsoon (August and September), land surface ET increases as recycled precipitation increases, a pattern that is more prominent in the Ganga Basin and Northeast India. This recycled precipitation accounts for approximately 20–25% of the rainfall in North India (Ganga Basin) and Northeast India during August and September. Hence, deforestation associated with changes in LULC may affect ET and may significantly affect monsoon rainfall [Paul 2016: 1-2].
Summarizing their own study, the authors conclude:
Here, we performed a sensitivity analysis to quantify the impacts of large-scale conversion from woody savannah to crop land in India on monsoon precipitation. We found such a change results in decreased ET and subsequently a decrease in recycled precipitation leading to a decline in monsoon precipitation. This decline is similar in extent to the observed recent decadal weakening of monsoon precipitation. However, other reasons may account for this observed weakening, such as the warming of Indian Ocean SST [sea surface temperature] [Paul 2016: 5-6].
To better clarify causality of the weakening monsoon, given other potential factors, the authors propose that:
The future scope of this present work is to perform detection and attribution studies for potential declines of Indian monsoons with model runs forced with SST warming only, aerosol forcing only, LULC changes only and all controlling factors together [Paul 2016: 6].
Tropical reforestation and climate change: beyond carbon, Locatelli 2015
When managed with both climate adaptation and mitigation in mind, tropical reforestation (TR) can serve multiple synergistic functions. TR mitigates regional and global climate change, not only by sequestering carbon but also through biophysical cooling (via evapotranspiration), by recycling rainfall regionally, and by reducing pressure on old growth forests.
Furthermore, TR helps local communities adapt to climate change by recharging stream flow in the dry season, reducing the severity of floods, protecting slopes against landslides and, through mangrove reforestation, reducing the impact of coastal storms and waves. Reforestation also creates livelihood opportunities through the sustainable harvest of forest products, and creates shelter and habitat for species vulnerable to climate change. However, to achieve this broad range of benefits, “reforestation practices should be designed to avoid the implementation of one strategy (mitigation or adaptation) to the detriment of the other.” Arguing for the application of what they term “climate-smart reforestation,” the authors recommend the following:
The challenge for climate-smart reforestation is to implement an effective combination of approaches to meet all three objectives: societal adaptation, climate mitigation, and ecological resilience [Locatelli 2015: 4].
However, as most policies consider the three objectives of climate-smart reforestation separately, they often overlook possible trade-offs and synergies. For example,reforestation projects managed with a carbon purpose could have detrimental consequences on water availability in the semi-arid tropics (Trabucco et al. 2008) or on biodiversity (O’Connor 2008). By contrast, reforestation that is explicitly climate-smart uses a multi-objective planning focus that enables different objectives to reinforce each other so that their interactions produce synergies rather than trade-offs. For example, tree regeneration in Tanzania under the Ngitili resource management system achieves carbon storage together with improved watershed conservation and greater provision of natural resources (water, food, and fodder) for livelihoods (Duguma et al. 2014). A proposed adaptation project in Colombia aims to reforest with flood-resistant native tree species to reduce flood impacts on downstream communities (UNDP 2012). A project in Costa Rica is testing different mixes of species and silvicultural practices to reduce vulnerability to storms and fires while also achieving carbon storage (Locatelli et al. 2011) [Locatelli 2015: 4-5].
This article underscores a key concept of this compendium – that functioning ecosystems (whether old growth or restored forests, for example) provide multiple, interwoven functions that support human and biodiverse life by regulating local, regional and global climate conditions.
Water-retention potential of Europe’s forests: A European overview to support natural water-retention measures, European Environment Agency (EEA) 2015
The importance of water retention (the rainfall absorbed or used within an ecosystem) for mitigating flood and drought conditions and contributing to clean drinking water, for example, has been increasingly recognized in Europe in the past decade. Along with wetland preservation, better agriculture practices and other measures, preserving and re-growing forests are seen as key to enhanced natural water retention. Forests cover a third of Europe, and:
can soak up excess rainwater, preventing run-offs and damage from flooding. By releasing water in the dry season, forests can help to provide clean water and mitigate the effects of droughts [EEA 2015: 6].
In recognition of the important water management role of forests and other natural ecosystems, new policy instruments have proposed Natural Water-Retention Measures (NWRMs).
Natural Water-Retention Measures (NWRMs) are defined as ‘measures to protect and manage water resources and to address water-related challenges by restoring or maintaining ecosystems, natural features and characteristics of water bodies using natural means and processes’ (European Commission and Directorate-General for the Environment 2014). … The main focus is to enhance and preserve the water retention capacity of aquifers, soil and ecosystems and improve their status [EEA 2015: 9].
This EEA study found that:
In water-basins where the forest cover is 30%, water retention is 25% higher than in basins where the forest cover is only 10%. In basins where the forest cover is 70%, water retention is 50% higher than in basins where the forest cover is only 10%. … Coniferous forests in general retain 10% more water than broadleaved forests or mixed forests [EEA 2015: 5].
Why Climate Change Makes Riparian Restoration More Important than Ever: Recommendations for Practice and Research, Seavy 2009
Riparian[14] ecosystems are naturally resilient, provide linear habitat connectivity, link aquatic and terrestrial ecosystems, and create thermal refugia for wildlife: all characteristics that can contribute to ecological adaptation to climate change [Seavy 2009: 330].
Arguing for the restoration of riparian areas because of their ecological significance and inherent resilience, these authors articulate the importance of both surface and groundwater – protected within a biodiverse ecosystem – for its cooling effect.
Because riparian areas have higher water content than surrounding upland areas, they absorb heat and buffer organisms against extreme temperatures (Naiman et al. 2000). During previous periods of climate change, riparian areas served as refugia because they provided microclimates that protected plant biodiversity (Bakker 1984, Meave and Kellman 1994). Riparian vegetation can maintain cooler water temperatures by shading water from sunlight (Sridhar et al. 2004, Cassie 2006) and the infusion of cold groundwater into warmer surface waters creates and maintains pockets of cool water (Chu et al. 2008). Thus, riparian areas provide thermal refugia for animals with thermoregulatory limitations [Seavy 2009: 332].
In this third issue of the Compendium of Scientific and Practical Findings Supporting Eco-Restoration to Address Global Warming by Biodiversity for a Livable Climate (“Bio4Climate”), we focus on the pivotal roles of water cycles and soil ecology in stabilizing ecosystems and the climate.
“The world is divided politically, but ecologically it is tightly interwoven.” – Carl Sagan, 1980, Cosmos
The magnitude of troubles ailing humanity is dizzying, if not terrifying – any 10 minutes of exposure to the daily news can attest to this. It’s hard to untangle the problems from each other, or to connect causes to effects, let alone to identify solutions that will work. That may well be because we tend to focus on symptoms rather than root causes. Wealth inequality, climate change and perpetual war are not inevitable conditions, but they are natural outcomes of the systems that produce them. With this in mind, here we explore solutions to the climate crisis from a systems perspective, meaning we seek to understand what complex systems are, how they work, and then to place what we observe of the world into this context.
Complex systems exist and behave the way they do based on the relationships among their component parts, as well as their interactions with other overlapping systems and subsystems. The Earth system is made up of a few major sub-systems: atmosphere, biosphere, hydrosphere, lithosphere, and human socio-economic systems [Donner 2009], within which countless other systems operate and interact. Elements, like carbon, cycle through systems as inputs and outputs, connecting systems to each other, and also collecting in various stocks, depending on rates of input and output.
Ecosystems, and indeed the global biosphere, are prototypical examples of complex adaptive systems, in which macroscopic system properties such as trophic structure, diversity–productivity relationships, and patterns of nutrient flux emerge from interactions among components, and may feed back to influence the subsequent development of those interactions [Levin 1998:431].
In other words, every system is greater than the sum of its parts. This is due to the relationships among parts giving rise to distinct patterns of behavior expressed by the system itself, often referred to as emergent properties, which in turn affect component parts. The human body is an example of a complex system, where all the organs working together give life to a person, and the behavior or the person affects the health of her organs. At the same time, individuals influence larger complex systems of which we are a part, such as our labor and spending in an economy; we are in turn influenced by economic volatility.
To understand global warming in the context of the Earth system, then, is to focus on how many components of the system interact to produce this outcome. It is to understand not only that greenhouse gas emissions trap heat in the atmosphere, but also that vegetation cools the Earth through evapotranspiration thereby generating rainfall that would otherwise be absent, while also drawing carbon out of the atmosphere. It is to further understand that vegetation is better protected and more productive in the presence of a greater degree of biodiversity. It is to accept and appreciate the vast complexity of billions of simultaneous processes that cannot be fully controlled, and yet also to recognize the larger patterns that restore balance to the systems sustaining all life. And it is to more fully account for how human systems interact with the other Earth systems.
As many studies in this Compendium show, Earth abounds with connections and causes that may surprise us. For instance, groundwater depletion is a source of CO2 emissions [Wood 2017], mushrooms cause rain [Hassett 2015], termite mounds mitigate drought effects [Bonachela 2017], and the Earth’s vast biosphere evolved into being thanks to a fungi-plant partnership [Mills 2017]. Understanding the planet as a complex system, encompassing myriad living and non-living subsystems, opens up our awareness to the interdependence among seemingly unrelated things and processes, and to the possibility of indirect effects and unintended consequences.
A systems framework also helps us understand the urgency of the crisis, given dynamics of complex systems that can lead to abrupt, transformational changes in the system. For instance, there are often time and space lags between cause and effect, as well as indirect effects, obscuring our awareness of the causes and consequences of our actions. Each time we drive a car, for example, we contribute to air pollution, acid rain, and climate change, however slightly. Yet, in the moment of driving, we are spared any immediately perceptible evidence of these effects. “With respect to climate change, greenhouse gases have accumulated in piecemeal fashion, with each car, cow, power plant, etc., having a minor effect. However, combining these small-scale impacts, through space and time, has manifested in large-scale effects that affect the entire planet” [Ingwersen 2013:4]
In addition, complex systems are influenced by positive or negative feedback loops, which either amplify a change or control it, respectively. Due at least in part to positive feedback mechanisms, complex systems exhibit nonlinear responses, meaning “that a very small change in some parameters can cause great qualitative differences in the resulting behavior”, as opposed to “the response of a linear system to small changes in its parameters or to changes in external forcing,” which “is usually smooth and proportionate to the stimulation” [Rial 2004:12].
Nonlinear behavior is triggered when the trajectory of a gradual change crosses a “tipping point,” or threshold, beyond which the system no longer maintains its equilibrium, and it changes abruptly into a new state. It’s akin to the age-old expression, “the straw that broke the camel’s back” – in other words, though increasingly strained the camel bore the weight of more and more straw, but only up to a point. An example of nonlinear change is the Arctic Sea, which hasn’t been ice free for more than 100,000 years but is now declining by 13.2% per decade [NASA] and could lose its summer ice entirely within a matter of years.
Understanding the climate crisis as a symptom of the global destruction of multiple interacting earth systems, rather than simply as the result of a buildup of greenhouse gases, leads us to different solutions. As Rockstrom et al [2009] suggest, in the interest of preserving the stable Holocene climate system we have known since before the dawn of agriculture, humanity’s response to climate change must account for multiple Earth system thresholds that are not to be crossed. “Since the industrial revolution (the advent of the Anthropocene), humans are effectively pushing the planet outside the Holocene range of variability for many key Earth System processes [Steffen et al. 2004].
Without such pressures, the Holocene state may be maintained for thousands of years into the future” [Rockstrom 2009:2]. The authors identify several other Earth system processes, including: ocean acidification, ozone depletion, aerosol loading, biodiversity loss, land-use change, nutrient and chemical pollution, and freshwater use, where “transgressing one or more planetary boundaries may be deleterious or even catastrophic due to the risk of crossing thresholds that will trigger non-linear, abrupt environmental change within continental- to planetary-scale systems” [Rockstrom 2009:1].
In other words, it’s not enough to solve the climate crisis only by switching to 100% renewable energy if the many other processes leading to ecological collapse are left unchecked. The explanation Rockstrom et al. offer sensitizes us to the strong interdependence among earth systems and processes, showing, for instance, that global warming can interact with (exacerbate or be exacerbated by) biodiversity loss, and with the human systems. How we respond to climate change can exert a positive (amplifying) feedback, for example, by ramping up our energy-intensive industrial defenses (artificial seawalls, air conditioners, geoengineering), or a negative (controlling) feedback – restoring coastal wetlands to mitigate hurricane damage, and increasing urban tree canopy to cool cities.
Donella Meadows, a systems thinker and sustainability advocate, offers an approach to intervening in these complex and overlapping systems to influence outcomes. She notes that we often look for leverage points in the wrong places, such as in the parameters. Parameters are what control the rate of flux into or out of a system, such as CO2 into or out of the atmosphere, and are often controlled through policy changes, such as changes in tax rates, minimum wage, and air quality standards, for instance [Meadows 1999]. Yet “if the system is chronically stagnant, parameter changes rarely kick-start it” [Meadows 1999: 8] because they are usually too small to trigger change in the overall goals or design of the system.
Meadows explains that a more powerful leverage point is the system’s guiding paradigm, such as that of our socio-economic systems that infinite growth will necessarily improve the human condition. Perhaps what’s needed is to reveal the current paradigm’s blind spot: that the planet’s resources, which fuel economic growth and absorb its waste products, are finite. Or a guiding paradigm of our political systems that downplays the role of biodiversity and ecosystem restoration/conservation by relying on emissions reductions to solve the climate crisis.
Paradigm change for an individual can happen in an instant, Meadows explains; for a whole society, it’s more complicated, though still possible. “In a nutshell, you keep pointing at the anomalies and failures in the old paradigm, you keep coming yourself, and loudly and with assurance from the new one, you insert people with the new paradigm in places of public visibility and power. You don’t waste time with reactionaries; rather you work with active change agents and with the vast middle ground of people who are open-minded” [Meadows 1999:18].
In our daily lives, we are well sensitized to the processes of our socio-economic systems – working for a paycheck, taking care of our families, and tending to our social networks. This is normal – it’s called living one’s life. We are much less sensitized to how our human systems interact with Earth systems, because any one person doesn’t necessarily need to consider this link to ensure his/her near-term survival or wellbeing. An exception may be farmers, whose wellbeing does depend on the land, and who are thus more likely to be in tune with Earth systems in a local context.
In general, though, we (at least in the West) rely directly on cars, buses, pavement, electricity, refrigerators, grocery stores, and plastic packaging, and only indirectly on the ecosystem processes that make these technologies possible. This is a glaring blind spot in one of our guiding paradigms – that our technology can save us. It’s a failure to visualize the world as a complex system with all its components and subsystems open and interacting, and to clearly perceive how the ingrained patterns of our daily lives, manifesting from the design of our socio-economic systems, are driving the cycles of Earth’s systems beyond the limits of equilibrium.
This is a glaring blind spot in one of our guiding paradigms – that our technology can save us. It’s a failure to visualize the world as a complex system with all its components and subsystems open and interacting, and to clearly perceive how the ingrained patterns of our daily lives, manifesting from the design of our socio-economic systems, are driving the cycles of Earth’s systems beyond the limits of equilibrium.
In short, the climate crisis we face isn’t just about greenhouse gases, biodiversity loss, poor soil health, or depleted aquifers, nor is it only about the food system, industrial society, poor individual choices, the military industrial complex, or unaccountable corporations, or any of a long list of ills.It’s about all of them in constant interactions, and our solutions need to account for that.
Global Warming has been a message of warning since climate research and discussions began roughly two hundred years ago in western science. Today, the predominance of the future tense in the climate dialogue has set the tone and expectations that however many times the “window of opportunity” for meaningful climate action were to close, it would surely open again. Casting targets in the seemingly distant future such as 2050 (more seemingly distant in 1992 than today), provided a psychological cushion that eased us into a dreamlike state where even repeatedly failed international negotiations still left time for reprieve and salvation.
Biodiversity for a Livable Climate has, to a certain extent, been complicit in this silence-in-urgency. There has been so much fear, confusion and denial of climate reality that we as an organization wanted to calm the conversation over the past three years to provide a safer place, a place of openness to new perspectives, to consider alternatives that most people hadn’t yet addressed nor imagined.
On the one hand, we feel that this strategy has shown some success. By emphasizing the positive, particularly amidst the frenzy induced by the 2016 election cycle, we have continued to introduce biodiversity and eco-restoration strategies to thousands of people, provided many examples of positive action, and helped build support for regenerative land management on many millions of acres. There is a palpable difference in the conversation and research today than there was only two years ago, and we should all celebrate that (and that’s part of the purpose of this compendium).
On the other hand, the positive feedback loops in climate,[12] as witnessed by the dramatic accelerations of weather crises, and many other environmental degradations such as extinctions (including unprecedented disappearance of insects), species migrations, plankton loss, and disrupted lifecycle timing, raise the urgency to such a level of alarm that even wanting to focus on the positive developments must make room for acknowledging our current dire situation.
Over the past thirty years, if we communicators of catastrophe have learned anything, it’s that it’s both extremely difficult and delicate to scare people into positive action. At the outset of the climate endeavor the assumption was that if we just give people the facts, we would all pretty much line up in a march of rational behavior.
As it turns out, in some senses the least of our problems with eco-collapse is the eco-collapse itself. The primary problem has been human psychology and culture, particularly the culture of civilizations, which relies on overshoot and inevitable collapse. We have met the enemy, and more truly than perhaps even Walt Kelly and the organizers of Earth Day 1970 realized, the enemy is deeply us [Kelly 1970].
While there are likely already some very unpleasant surprises in the pipeline, and outcomes are by no means certain, there is good news as well: the urgency is addressable. The tectonic level of destruction that we’ve set up in but a blink of geological time is reversible. The solutions are largely already known. What we’re struggling with is the psyche of that most puzzling of species, homo sapiens.
At this point, therefore, it behooves us to briefly outline the current urgency to paint what we hope is a clearer picture, based on a realistic foundation that rejects wishful thinking in favor of action moving forward.
Again, there is some good news: the zeitgeist, the spirit of the times which emerges from the sum total of human culture and the planetary context in which it must function, is finally setting a determined course towards a global systems approach. The zeitgeist has, in the nick of time, begun to address root causes of ecosystem collapse and not just obvious symptoms, as painful as they are. Symptoms are as varied as Nature itself, but root causes – anthropogenic degradation and desertification of the biosphere – are surprisingly straightforward.
We can solve this, and indeed, as we put our dominance of Nature behind us we already are. Nature will be happy to collaborate if only we were to pledge our allegiance to a few immutable laws of biology established over billions of years: repeal just isn’t in the cards. That’s the heart of what this Compendium is about. The time is short, the burdens daunting, but for the sake of preserving and protecting life on Earth we have little choice.
===
In 1958 Charles Keeling began recording the now-classic curve of carbon dioxide concentrations at the Scripps Mauna Loa Observatory in Hawaii (the sawtooth annual variations of 6-8 ppm are a result of increased uptake of CO2 during the summer growing season in the Northern Hemisphere followed by the release of CO2 during the winter).
In that same year, Oscar-winning director Frank Capra released a film on the weather for the Bell Laboratory Science Series. Towards the end of the film there is a brief dramatic clip of global warming, collapsing ice and belching industry, indicating that we had a surprisingly good idea of what climate effects were about in 1958. It is well worth a look, both for the climate observations and the sense of the scientific over-confidence of the post-atomic age (start at around 47:50 of the full version). [Bell Labs 1958]
The sense of climate urgency in 1958, however, subsumed in the enthusiasm over the aspirations of science and overshadowed by the anxiety of the cold war, was understandably remote.
Fast forward to 1988 and James Hansen’s testimony before Congress. As reported in the New York Times,
If the current pace of the buildup of these gases continues, the effect is likely to be a warming of 3 to 9 degrees Fahrenheit from the year 2025 to 2050, according to these projections. This rise in temperature is not expected to be uniform around the globe but to be greater in the higher latitudes, reaching as much as 20 degrees, and lower at the Equator.
The rise in global temperature is predicted to cause a thermal expansion of the oceans and to melt glaciers and polar ice, thus causing sea levels to rise by one to four feet by the middle of the next century. Scientists have already detected a slight rise in sea levels. At the same time, heat would cause inland waters to evaporate more rapidly, thus lowering the level of bodies of water such as the Great Lakes. [http://www.nytimes.com/1988/06/24/us/global-warming-has-begun-expert-tells-senate.html?pagewanted=all]
These predictions, using basic data and newly evolving models, were stark, even from today’s perspective. Hansen and others initiated the beginnings of a global emergency mindset. But despite leading to a series of international conferences and treaties, they were caught in a mire of political and economic resistance that has thus far proved an almost immovable force. While progress has been made on many fronts in the efforts to address greenhouse gases, none has yet yielded differences which will change the course of climate events. That will require a systems approach to the natural world that is only now in the process of coming to fruition, and which is the topic of this Compendium.
Hansen has gone on to become one of the most powerful and courageous voices in the mainstream climate science arena, stepping beyond his role as scientist and into the public and political arenas. He has given measured but insistent voice to the current urgency:
Global temperature is a fundamental climate metric highly correlated with sea level, which implies that keeping shorelines near their present location requires keeping global temperature within or close to its preindustrial Holocene range. However, global temperature excluding short-term variability now exceeds 1o C relative to the 1880–1920 mean and annual 2016 global temperature was almost 1.3o C. We show that global temperature has risen well out of the Holocene range and Earth is now as warm as it was during the prior (Eemian) interglacial period, when sea level reached 6–9m higher than today. Further, Earth is out of energy balance with present atmospheric composition, implying that more warming is in the pipeline, and we show that the growth rate of greenhouse gas climate forcing has accelerated markedly in the past decade. The rapidity of ice sheet and sea level response to global temperature is difficult to predict, but is dependent on the magnitude of warming. Targets for limiting global warming thus, at minimum, should aim to avoid leaving global temperature at Eemian or higher levels for centuries. Such targets now require “negative emissions”, i.e., extraction of CO2 from the air. If phasedown of fossil fuel emissions begins soon, improved agricultural and forestry practices, including reforestation and steps to improve soil fertility and increase its carbon content, may provide much of the necessary CO2 extraction. In that case, the magnitude and duration of global temperature excursion above the natural range of the current interglacial (Holocene) could be limited and irreversible climate impacts could be minimized. In contrast, continued high fossil fuel emissions today place a burden on young people to undertake massive technological CO2 extraction if they are to limit climate change and its consequences. Proposed methods of extraction such as bioenergy with carbon capture and storage (BECCS) or air capture of CO2 have minimal requirement of negative CO2 emissions estimated costs of USD 89–535 trillion this century and also have large risks and uncertain feasibility. Continued high fossil fuel emissions unarguably sentences young people to either a massive, implausible cleanup or growing deleterious climate impacts or both. [Hansen et al., 2017]
A dire picture indeed.
Notwithstanding Hansen’s steady voice, there are reasons for more urgent statements still. Positive feedbacks, i.e., climate accelerations, are asserting prominence, particularly in the striking accumulations of high temperature records and unprecedented weather calamities occurring during 2017.
On July 19, 2012, popular climate author Bill McKibben wrote “Global Warming’s Terrifying New Math: Three simple numbers that add up to global catastrophe – and that make clear who the real enemy is.” [McKibben 2012] The real enemy according to McKibben is the purveyors of fossil fuels.
McKibben puts forth what he calls three terrifying numbers:
2 degrees C, a dubious safe limit
565 Gt, the maximum additional carbon by mid-century to keep within that safe limit
2,795 Gt, the carbon in already-proven coal, gas and oil reserves
Given the rate at which we were burning fossil fuels, the math doesn’t offer a good prognosis, and McKibben concludes that, “So far, as I said at the start, environmental efforts to tackle global warming have failed.” [McKibben 2012]
Over five years later on December 1, 2017, McKibben wrote “Winning Slowly Is the Same as Losing: The technology exists to combat climate change – what will it take to get our leaders to act?” [McKibben 2017] His sense of alarm is clear:
If we don’t win very quickly on climate change, then we will never win. That’s the core truth about global warming. It’s what makes it different from every other problem our political systems have faced. . . . It won’t stand still.
McKibben is also sensible with respect to his perspective on Donald Trump’s administration, i.e., that we were failing on climate long before the new president arrived, with the possible implication that we may even be avoiding our own failures by obsessing with his:
[W]e weren’t moving fast enough to catch up with physics before Trump. In fact, it’s even possible that Trump – by jumping the climate shark so spectacularly – may runsome small risk of disrupting the fossil-fuel industry’s careful strategy.
While McKibben hails the ongoing progress in opposing the fossil fuels industry, the urgency of the timing and limited effectiveness of the actions weighs heavily upon him:
At 350.org, we’re rolling out a vast Fossil Free campaign across the globe this winter, joining organizations like the Sierra Club to pressure governments to sign up for 100 percent renewable energy, blocking new pipelines and frack wells as fast as the industry can propose them, and calling out the banks and hedge funds that underwrite the past. It’s working – just in the last few weeks Norway’s sovereign wealth fund, the largest in the world, announced plans to divest from fossil fuels, and the Nebraska Public Service Commission threw yet more roadblocks in front of the Keystone pipeline.
But the question is, is it working fast enough? Paraphrasing the great abolitionist leader Theodore Parker, Martin Luther King Jr. used to regularly end his speeches with the phrase “the arc of the moral universe is long but it bends toward justice.” The line was a favorite of Obama’s too, and for all three men it meant the same thing: “This may take a while, but we’re going to win.” For most political fights, it is the simultaneously frustrating and inspiring truth. But not for climate change. The arc of the physical universe appears to be short, and it bends toward heat. Win soon or suffer the consequences.
From Bio4Climate’s perspective, McKibben’s frank assessment and even despair are laudable. Admitting failure is an opening to new solutions. What is discouraging is his reluctance to at least consider the full potential of eco-restoration to overcome both geophysical and political obstacles to positive action on multiple pressing environmental issues, including climate.[13]
On June 20, 2017 an Australian organization, Breakthrough [Breakthrough 2017],[14] produced a no-nonsense report entitled Disaster Alley: Climate Change, Conflict & Risk.[15] Among its blunt points:
From tropical coral reefs to the polar ice sheets, global warming is already dangerous. The world is perilously close to, or passed, tipping points which will create major changes in global climate systems.
The world now faces existential climate-change risks which may result in “outright chaos” and an end to human civilisation as we know it.
These risks are either not understood or wilfully ignored across the public and private sectors, with very few exceptions.
Global warming will drive increasingly severe humanitarian crises, forced migration, political instability and conflict. The Asia–Pacific region, including Australia, is considered to be “Disaster Alley” where some of the worst impacts will be experienced.
Building more resilient communities in the most vulnerable nations by high-level financial commitments and development assistance can help protect peoples in climate hotspots and zones of potential instability and conflict.
Australia’s political, bureaucratic and corporate leaders are abrogating their fiduciary responsibilities to safeguard the people and their future well-being. They are ill-prepared for the real risks of climate change at home and in the region.
The Australian government must ensure Australian Defence Force and emergency services preparedness, mission and operational resilience, and capacity for humanitarian aid and disaster relief, across the full range of projected climate change scenarios.
It is essential to now strongly advocate a global climate emergency response, and to build a national leadership group outside conventional politics to design and implement emergency decarbonisation of the Australian economy. This would adopt all available safe solutions using sound, existential risk-management practices.
While Disaster Alley is primarily concerned with the Asia-Pacific region, the issues are clearly global. Following closely on Disaster Alley, on September 6, 2017, Breakthrough published What Lies Beneath: The Scientific Understatement of Climate Risks [Breakthrough 2017b],which further emphasized the chronic underestimation of climate risk in mainstream science, and the risks posed by poor risk analysis itself, which is a chronic condition in much of mainstream climate science.
It is now clear that climate change is an existential risk to human civilisation: that is, an adverse outcome that would either annihilate intelligent life or permanently and drastically curtail its potential. Temperature rises that are now in prospect, even after the Paris Agreement, are in the range of 3–5°C. The Paris Agreement voluntary emission reduction commitments, if implemented, would result in the planet warming by 3°C, without taking into account “long-term” carbon-cycle feedbacks. With a higher climate sensitivity figure of 4.5°C, for example, which would account for such feedbacks, the Paris path would lead to around 5°C of warming, according to a MIT study. A study by Schroder Investment Management published in June 2017 found — after taking into account indicators across a wide range of the political, financial, energy and regulatory sectors — the average temperature increase implied across all sectors was 4.1°C.
Warming of 4°C or more could reduce the global human population by 80% or 90%, and the World Bank reports “there is no certainty that adaptation to a 4°C world is possible.” A study by two US national security think tanks concluded that 3°C of warming and a 0.5 metre sea-level rise would likely lead to “outright chaos”. A recent study by the European Commission’s Joint Research Centre found that if global temperatures rise 4°C, then extreme heatwaves with “apparent temperatures” peaking at over 55°C (131o F) will begin to regularly affect many densely populated parts of the world. At 55°C or so, much activity in the modern industrial world would have to stop. (“Apparent temperatures” refers to the Heat Index, which quantifies the combined effect of heat and humidity to provide people with a means of avoiding dangerous conditions.) [p. 9]
Finally, on July 9, 2017, a climate reality bludgeon reached the American (and global) public with the publication of “Uninhabitable Earth” in New York Magazine.[16] More widely read by far than any of the other references here, author David Wallace-Wells interviewed numerous climate scientists about bad- and worst-case scenarios.
It is, I promise, worse than you think. If your anxiety about global warming is dominated by fears of sea-level rise, you are barely scratching the surface of what terrors are possible, even within the lifetime of a teenager today. And yet the swelling seas — and the cities they will drown — have so dominated the picture of global warming, and so overwhelmed our capacity for climate panic, that they have occluded our perception of other threats, many much closer at hand. Rising oceans are bad, in fact very bad; but fleeing the coastline will not be enough.
Indeed, absent a significant adjustment to how billions of humans conduct their lives, parts of the Earth will likely become close to uninhabitable, and other parts horrifically inhospitable, as soon as the end of this century.
Wallace-Wells digs wider and deeper, and while many climate scientists objected to at least some of his inferences and logic, overall he’s on solid ground. It is, after all, the ground that these scientists helped prepare – and all in all, they did their jobs well.
To this sad litany we would only add that the most terrible, excruciating lethal strokes that we face from climate catastrophe are more likely the whimper than the bang. While sea-level rise, hurricanes and fires are dramatic, the primary cause of death for billions of people will be no food and no water.[17]
This study compared three corn-soybean cropping systems: (1) conventional 2-yr rotation with chemical inputs, and residues returned to soil; (2) a longer (than 2 years), organic rotation with grass/legume hayed and returned to soil in manure; and (3) a longer (than 2 years) organic rotation with grass/legume turned back into the soil directly. Even though the conventional system returned more total residue to the soil, carbon sequestration was significantly lower for the conventional system than for the two organic, legume-based systems. Authors suggest that this is due to greater temporal plant diversity from the longer rotations, and higher quality residue (greater N:C) in the two legume-based organic systems. Furthermore, CO2 emissions were lower in the legume-based organic systems due to 50% lower energy use.
Even though the conventional system returned more total residue to the soil, carbon sequestration was significantly lower for the conventional system than for the two organic, legume-based systems. Authors suggest that this is due to greater temporal plant diversity from the longer rotations, and higher quality residue (greater N:C) in the two legume-based organic systems. [Drinkwater]
A sequestration rate of 2.363t C/ha/yr was demonstrated where compost made of dairy manure and leaves was applied to fields in a three year rotation of corn-vegetable-small grain, with leguminous cover crops. The same rotation treated with chemical fertilizer instead of compost resulted in a net loss of -0.317t C/ha/yr.
This meta-analysis estimates total carbon sequestration potential in the US from various agroforestry practices to be 530 TgC/year (530 million metric tons), equivalent to about 1/3 of annual US carbon emissions from fossil fuel combustion. Based on their literature review, the authors estimate per-hectare sequestration rates (based on aboveground and belowground carbon accumulation) for each practice as follows: 6.1t C/ha/yr (silvopastoral), 3.4t C/ha/yr (alleycropping), 6.4t C/ha/yr (windbreaks), 2.6t C/ha/yr (riparian buffer).
An analysis of hundreds of soil samples collected from organic and conventional farms around the US shows higher average percentages both of total SOM and of humic substances – a measure of carbon sequestration – for organic farm soils compared to conventional farm soils. The mean percent humification (humic substances divided by total SOM) for organic soils is 57.3%, compared to 45.6% for conventional soils.
This review examines “the state of knowledge for the stocks of, inputs to, and outputs from SOM around the world” [Jackson 2017: 422], with a view toward developing better understanding of processes that stabilize SOM. It explains the biological processes involved in carbon cycling and storage, finding that “root inputs are approximately five times more likely than an equivalent mass of aboveground litter to be stabilized as SOM” [Jackson 2017: 420]. Litter input can either increase or decrease SOM, despite the assumption in most carbon models that there is a linear relationship between litter input and transformation of carbon into more stable forms. This finding suggests that perennials and other deep-rooting plants have an important role to play with respect to carbon sequestration. As the author puts it:
Managing carbon inputs and relative allocation, for instance, through selection for deep roots or for greater belowground allocation in crops (Kell 2011), has been suggested as a way to increase SOM formation and stabilization in such systems (Bolinder et al. 2007, Eclesia et al. 2016). However, plant breeding has traditionally selected for aboveground yields alone; therefore, potential trade-offs between yield and root production must be carefully evaluated (DeHaan et al. 2005). New tools for monitoring root systems and in situ SOM in the field are needed (Molon et al. 2017) [Jackson 2017: 422]. . . .
The importance of root inputs for SOM formation is likely attributable to both their chemical composition and, almost certainly, their presence in the soil; upon death, they immediately interact with soil minerals, microbes, and aggregates. Roots tend to be characterized more by aliphatic compounds that are readily sorbed to mineral surfaces, and their composition (and that of root exudates) can increase microbial carbon use efficiency (CUE), defined as the ratio of microbial growth to carbon uptake, more than litter can. High CUE promotes microbial growth and carbon stabilization in mineral-associated soil pools, and low CUE favors biomass respiration (Manzonietal.2012a) [Jackson 2017: 423]. . . .
Soils hold the largest biogeochemically active terrestrial carbon pool on Earth and are critical for stabilizing atmospheric CO2 concentrations. Nonetheless, global pressures on soils continue from changes in land management, including the need for increasing bioenergy and food production [Jackson 2017: 420].
. . . plant breeding has traditionally selected for aboveground yields alone; therefore, potential trade-offs between yield and root production must be carefully evaluated[Jackson 2017].
Abstract: Human occupation is usually associated with degraded landscapes but 13,000 years of repeated occupation by British Columbia’s coastal First Nations has had the opposite effect, enhancing temperate rainforest productivity. This is particularly the case over the last 6,000 years when intensified intertidal shellfish usage resulted in the accumulation of substantial shell middens. We show that soils at habitation sites are higher in calcium and phosphorous. Both of these are limiting factors in coastal temperate rainforests. Western red cedar (Thuja plicata) trees growing on the middens were found to be taller, have higher wood calcium, greater radial growth and exhibit less top die-back. Coastal British Columbia is the first known example of long-term intertidal resource use enhancing forest productivity and we expect this pattern to occur at archaeological sites along coastlines globally [Trant 2016: 1].
Although focused on forests and not farmland, this study shows that, as in the Amazon, where indigenous people created SOM-rich terra preta soil (akin to biochar-enhanced soil), human populations can increase soil quality and ecosystem productivity beyond what the potential would have been absent human activity.
This is an interesting point with respect to global potential for soil carbon sequestration. Scientists often refer to an equilibrium point, up to which soils can regain carbon previously lost through exploitive human activity. Equilibrium is generally seen as being the point at which new SOC levels are equivalent to or somewhat less than what they were prior to human exploitation of the soil, and never greater than the original amount. While Trant et al. [2016] have found evidence of calcium and phosphorus (not carbon) enrichment due to human activity, their findings raise questions about the extent to which intentionally building soils through all the methods we know to maximize carbon storage could increase various soils’ presumed equilibrium points.
Human occupation is usually associated with degraded landscapes but 13,000 years of repeated occupation by British Columbia’s coastal First Nations has had the opposite effect, enhancing temperate rainforest productivity [Trant 2016: 1].
Edited by innovator and entrepreneur Paul Hawken, Drawdown is a remarkable and comprehensive work presenting eighty well-vetted solutions and twenty promising “coming attractions” to remove carbon from the atmosphere and restore planetary health. Hawken engaged numerous scientists, modellers, advisers, artists and writers, resulting in a beautifully illustrated and comprehensive exploration of possibilities for reversing global warming.
The impact of the book as a whole is as important as each solution: Drawdown presents a universe of actions that go far beyond what we can imagine if we consider only emissions reductions and alternative energy. It leads to an entirely different climate conversation from the one we’re used to, and offers many threads of hope.
Drawdown has something for everyone, covering sectors of Energy, Food, Buildings and Cities, Land Use, Transport, and Materials. Near the top of the list is Women and Girls, whose education has dramatic effects on population and is one of the most important climate positive steps we can take. Of course technology offerings abound, but they are amply balanced by discussions of biology and social change, often sorely missing in debates on global warming. Of particular interest in this Compendium are biological strategies; we’ll mention just three of them here.
Agroforestry in Burkina Faso
After terrible droughts in the 1980s resulted in a 20% reduction in rainfall and millions of deaths by starvation, farmer Yacouba Sawadogo enhanced a traditional practice of digging rain-capturing pits by adding manure. There were seeds in the manure and as a result trees began to grow, holding soils together with roots, protecting plantings from wind gusts that before had required frequent re-sowing, and opening channels that moved water into the soils and raised water tables. This foray into agro-forestry spread across the rural countryside to widespread beneficial effect.
Of great significance is that the expertise, invention and community organizing were native and local, required no foreign aid or expensive soil inputs, and in terms of money cost nothing. This is what sustainability can look like. [Hawken 2017: 118-120]
Pasture Cropping
In 1979, after a devastating fire destroyed his two-thousand acre farm in Australia, Colin Seis began to question why crops and animals couldn’t be profitably raised on the same land, effectively doubling output. Persisting through a difficult transition, Seis saw water retention improvements, decreased input costs, a virtual end to insect infestation, and measures of soil fertility and carbon content go up along with profits. Today, pasture cropping is practiced on over two thousand Australian farms and is spreading throughout the world. [Hawken 2017: 175]
Intensive Silvopasture
Silvopasture, the most common form of agroforestry, is the practice of combining trees and woody shrubs with pasture grasses. The result is healthier plant and animal growth, including sequestering a respectable one to four tons of carbon per acre. It is currently practiced on over one billion acres worldwide.
For remarkable next steps enter the intensive part of silvopasture, starting with a quickly growing, edible leguminous shrub, Leucaena leucocephala in Australia and Latin America (different species of shrubs are suitable in different ecosystems). Water retention improves, biomass increases, species biodiversity doubles, animal stocking rates almost triple, ambient temperatures decrease by 14 to 23 degrees F in the tropics, meat production increases by a factor of 4 to 10, and perhaps most strikingly, soil carbon sequestration rates have exceeded 10 tons per acre (conventional agriculture can claim 1 ton of carbon per acre or less, or even net carbon loss to the atmosphere). [Hawken 2017: 181]
This is one of the most comprehensive mainstream studies to date of a broad spectrum of natural climate solutions by thirty-two co-authors and supported by The Nature Conservancy. The report examines “20 conservation, restoration, and/or improved land management actions that increase carbon storage and/or avoid greenhouse gas emissions across global forests, wetlands, grasslands, and agricultural lands.” The authors “find that the maximum potential of NCS [Natural Climate Solutions] —when constrained by food security, fiber security, and biodiversity conservation—is 23.8 petagrams[11] of CO2 equivalent . . . This is ≥30% higher than prior estimates, which did not include the full range of options and safeguards considered here.” [Griscom 2017: 11645]. The study seeks to assess both the potential emissions from land use as well as the carbon-sequestration potential.
The study posits a target of <2o C as the conventionally agreed-upon safe limit:
Warming will likely be held to below 2 °C if natural pathways are implemented at cost-effective levels . . . and if we avoid increases in fossil fuel emissions for 10 y and then drive them down to 7% of current levels by 2050 and then to zero by 2095 [p. 11647]
The authors state that their estimates are intentionally conservative because (1) they do not include potential benefits of payments for high-money-value ecosystem services in stimulating NCS efforts; (2) they exclude various management practices where data were not “sufficiently robust for global extrapolation,” e.g., no-till, adaptive multi-paddock grazing, etc.; and (3) significant additional investment would be required to keep warming at 1.5o C. [Griscom 2017: 11648]
Detail is provided on contributions of specific mitigation pathways, such as forests, wetlands, grasslands, etc., and on challenges as well. For example, “Despite the large potential of NCS, land-based sequestration efforts receive only about 2.5% of climate mitigation dollars.” [Griscom 2017: 11648] This observation is consistent with our observations of limited available resources for the most basic NCS education. Other challenges include deforestation for farming and animal husbandry, losing high carbon sequestration benefits of wetlands due to reclamation, and impacts of climate feedbacks such as fire, drought, temperature increases, etc.
We applaud Griscom et al. for an excellent and comprehensive analysis and review of many of the factors in natural climate solutions. We do, however, believe that (1) the potential of nature’s solutions is far greater than Griscom et al. estimate, and (2) that the temperature limits (1.5o – 2o C) are too high and too dangerous – considering that natural processes are already changing, drastically and for the worse, with an average global temperature increase of barely 1o C (see Appendix A: Urgency of the Biodiversity and Climate Crisis).
The differences between the perspectives of Griscom et al. and those adopted in this Compendium are paradigmatic. Griscom et al. acknowledges that their estimates are conservative, looks at a set of studies that tends toward the mainstream and is primarily based on established and widespread practice. This is perfectly reasonable in the process of what Thomas Kuhn calls “normal science” (see Compendium Vol. 1 No. 1 for an extensive discussion of Kuhn’s landmark work). Unfortunately the process of normal science for accepting new thinking and discoveries usually takes decades, and we are currently in the throes of an extinction, and an emergency with respect to biodiversity, and climate change. Therefore we have to accelerate our response. Accordingly, Bio4Climate searches for studies that tend to examine positive variants, i.e., examples of what is possible beyond current conceptual boundaries. We emphasize goals to strive for, even if the data are not yet “sufficiently robust for global extrapolation.” The robustness of such data will increase with more intentional focus.
An interesting side effect of the paradigm difference is that numerous sources that we cite, many from the scientific literature, don’t appear in NCS references (for example, Richard Teague [Teague et al. 2016], Gabe Brown [Brown 2016], Tom Goreau [Goreau 2015], Rebecca Ryals and Whendee Silver [Ryals and Silver 2013], David Johnson [Johnson 2017], Paul and Elizabeth Kaiser [Kaiser 2017], Terry McCosker [McCosker 2000], Carol Evans and Jon Griggs [Evans et al., 2015], to name just a few). Nor are there discussions of permaculture or agroforestry, two of the more promising areas of research and practice in land management that lead to climate-positive results.
Unfortunately the process of normal science for accepting new thinking and discoveries usually takes decades, and we are currently in the throes of an extinction, and an emergency with respect to biodiversity, and climate change. Therefore we have to accelerate our response. Accordingly, Bio4Climate searches for studies that tend to examine positive variants, i.e., examples of what is possible beyond current conceptual boundaries. We emphasize goals to strive for, even if the data are not yet “sufficiently robust for global extrapolation.” The robustness of such data increases with intentional focus.
This is one of the most comprehensive mainstream studies to date of a broad spectrum of natural climate solutions by thirty-two co-authors and supported by The Nature Conservancy. The report examines “20 conservation, restoration, and/or improved land management actions that increase carbon storage and/or avoid greenhouse gas emissions across global forests, wetlands, grasslands, and agricultural lands.” The authors “find that the maximum potential of NCS [Natural Climate Solutions] —when constrained by food security, fiber security, and biodiversity conservation—is 23.8 petagrams[11] of CO2 equivalent . . . This is ≥30% higher than prior estimates, which did not include the full range of options and safeguards considered here.” [Griscom 2017: 11645]. The study seeks to assess both the potential emissions from land use as well as the carbon-sequestration potential.
The study posits a target of <2o C as the conventionally agreed-upon safe limit:
Warming will likely be held to below 2 °C if natural pathways are implemented at cost-effective levels . . . and if we avoid increases in fossil fuel emissions for 10 y and then drive them down to 7% of current levels by 2050 and then to zero by 2095 [p. 11647]
The authors state that their estimates are intentionally conservative because (1) they do not include potential benefits of payments for high-money-value ecosystem services in stimulating NCS efforts; (2) they exclude various management practices where data were not “sufficiently robust for global extrapolation,” e.g., no-till, adaptive multi-paddock grazing, etc.; and (3) significant additional investment would be required to keep warming at 1.5o C. [Griscom 2017: 11648]
Detail is provided on contributions of specific mitigation pathways, such as forests, wetlands, grasslands, etc., and on challenges as well. For example, “Despite the large potential of NCS, land-based sequestration efforts receive only about 2.5% of climate mitigation dollars.” [Griscom 2017: 11648] This observation is consistent with our observations of limited available resources for the most basic NCS education. Other challenges include deforestation for farming and animal husbandry, losing high carbon sequestration benefits of wetlands due to reclamation, and impacts of climate feedbacks such as fire, drought, temperature increases, etc.
We applaud Griscom et al. for an excellent and comprehensive analysis and review of many of the factors in natural climate solutions. We do, however, believe that (1) the potential of nature’s solutions is far greater than Griscom et al. estimate, and (2) that the temperature limits (1.5o – 2o C) are too high and too dangerous – considering that natural processes are already changing, drastically and for the worse, with an average global temperature increase of barely 1o C (see Appendix A: Urgency of the Biodiversity and Climate Crisis).
The differences between the perspectives of Griscom et al. and those adopted in this Compendium are paradigmatic. Griscom et al. acknowledges that their estimates are conservative, looks at a set of studies that tends toward the mainstream and is primarily based on established and widespread practice. This is perfectly reasonable in the process of what Thomas Kuhn calls “normal science” (see Compendium Vol. 1 No. 1 for an extensive discussion of Kuhn’s landmark work). Unfortunately the process of normal science for accepting new thinking and discoveries usually takes decades, and we are currently in the throes of an extinction, and an emergency with respect to biodiversity, and climate change. Therefore we have to accelerate our response. Accordingly, Bio4Climate searches for studies that tend to examine positive variants, i.e., examples of what is possible beyond current conceptual boundaries. We emphasize goals to strive for, even if the data are not yet “sufficiently robust for global extrapolation.” The robustness of such data will increase with more intentional focus.
An interesting side effect of the paradigm difference is that numerous sources that we cite, many from the scientific literature, don’t appear in NCS references (for example, Richard Teague [Teague et al. 2016], Gabe Brown [Brown 2016], Tom Goreau [Goreau 2015], Rebecca Ryals and Whendee Silver [Ryals and Silver 2013], David Johnson [Johnson 2017], Paul and Elizabeth Kaiser [Kaiser 2017], Terry McCosker [McCosker 2000], Carol Evans and Jon Griggs [Evans et al., 2015], to name just a few). Nor are there discussions of permaculture or agroforestry, two of the more promising areas of research and practice in land management that lead to climate-positive results.
Unfortunately the process of normal science for accepting new thinking and discoveries usually takes decades, and we are currently in the throes of an extinction, and an emergency with respect to biodiversity, and climate change. Therefore we have to accelerate our response. Accordingly, Bio4Climate searches for studies that tend to examine positive variants, i.e., examples of what is possible beyond current conceptual boundaries. We emphasize goals to strive for, even if the data are not yet “sufficiently robust for global extrapolation.” The robustness of such data increases with intentional focus.
Drawdown, Hawken, ed. 2017
Edited by innovator and entrepreneur Paul Hawken, Drawdown is a remarkable and comprehensive work presenting eighty well-vetted solutions and twenty promising “coming attractions” to remove carbon from the atmosphere and restore planetary health. Hawken engaged numerous scientists, modellers, advisers, artists and writers, resulting in a beautifully illustrated and comprehensive exploration of possibilities for reversing global warming.
The impact of the book as a whole is as important as each solution: Drawdown presents a universe of actions that go far beyond what we can imagine if we consider only emissions reductions and alternative energy. It leads to an entirely different climate conversation from the one we’re used to, and offers many threads of hope.
Drawdown has something for everyone, covering sectors of Energy, Food, Buildings and Cities, Land Use, Transport, and Materials. Near the top of the list is Women and Girls, whose education has dramatic effects on population and is one of the most important climate positive steps we can take. Of course technology offerings abound, but they are amply balanced by discussions of biology and social change, often sorely missing in debates on global warming. Of particular interest in this Compendium are biological strategies; we’ll mention just three of them here.
Agroforestry in Burkina Faso
After terrible droughts in the 1980s resulted in a 20% reduction in rainfall and millions of deaths by starvation, farmer Yacouba Sawadogo enhanced a traditional practice of digging rain-capturing pits by adding manure. There were seeds in the manure and as a result trees began to grow, holding soils together with roots, protecting plantings from wind gusts that before had required frequent re-sowing, and opening channels that moved water into the soils and raised water tables. This foray into agro-forestry spread across the rural countryside to widespread beneficial effect.
Of great significance is that the expertise, invention and community organizing were native and local, required no foreign aid or expensive soil inputs, and in terms of money cost nothing. This is what sustainability can look like. [Hawken 2017: 118-120]
Pasture Cropping
In 1979, after a devastating fire destroyed his two-thousand acre farm in Australia, Colin Seis began to question why crops and animals couldn’t be profitably raised on the same land, effectively doubling output. Persisting through a difficult transition, Seis saw water retention improvements, decreased input costs, a virtual end to insect infestation, and measures of soil fertility and carbon content go up along with profits. Today, pasture cropping is practiced on over two thousand Australian farms and is spreading throughout the world. [Hawken 2017: 175]
Intensive Silvopasture
Silvopasture, the most common form of agroforestry, is the practice of combining trees and woody shrubs with pasture grasses. The result is healthier plant and animal growth, including sequestering a respectable one to four tons of carbon per acre. It is currently practiced on over one billion acres worldwide.
For remarkable next steps enter the intensive part of silvopasture, starting with a quickly growing, edible leguminous shrub, Leucaena leucocephala in Australia and Latin America (different species of shrubs are suitable in different ecosystems). Water retention improves, biomass increases, species biodiversity doubles, animal stocking rates almost triple, ambient temperatures decrease by 14 to 23 degrees F in the tropics, meat production increases by a factor of 4 to 10, and perhaps most strikingly, soil carbon sequestration rates have exceeded 10 tons per acre (conventional agriculture can claim 1 ton of carbon per acre or less, or even net carbon loss to the atmosphere). [Hawken 2017: 181]
Intertidal resource use over millennia enhances forest productivity, Trant 2016
Abstract: Human occupation is usually associated with degraded landscapes but 13,000 years of repeated occupation by British Columbia’s coastal First Nations has had the opposite effect, enhancing temperate rainforest productivity. This is particularly the case over the last 6,000 years when intensified intertidal shellfish usage resulted in the accumulation of substantial shell middens. We show that soils at habitation sites are higher in calcium and phosphorous. Both of these are limiting factors in coastal temperate rainforests. Western red cedar (Thuja plicata) trees growing on the middens were found to be taller, have higher wood calcium, greater radial growth and exhibit less top die-back. Coastal British Columbia is the first known example of long-term intertidal resource use enhancing forest productivity and we expect this pattern to occur at archaeological sites along coastlines globally [Trant 2016: 1].
Although focused on forests and not farmland, this study shows that, as in the Amazon, where indigenous people created SOM-rich terra preta soil (akin to biochar-enhanced soil), human populations can increase soil quality and ecosystem productivity beyond what the potential would have been absent human activity.
This is an interesting point with respect to global potential for soil carbon sequestration. Scientists often refer to an equilibrium point, up to which soils can regain carbon previously lost through exploitive human activity. Equilibrium is generally seen as being the point at which new SOC levels are equivalent to or somewhat less than what they were prior to human exploitation of the soil, and never greater than the original amount. While Trant et al. [2016] have found evidence of calcium and phosphorus (not carbon) enrichment due to human activity, their findings raise questions about the extent to which intentionally building soils through all the methods we know to maximize carbon storage could increase various soils’ presumed equilibrium points.
Human occupation is usually associated with degraded landscapes but 13,000 years of repeated occupation by British Columbia’s coastal First Nations has had the opposite effect, enhancing temperate rainforest productivity [Trant 2016: 1].
The ecology of soil carbon: pools, vulnerabilities, and biotic and abiotic controls, Jackson 2017
This review examines “the state of knowledge for the stocks of, inputs to, and outputs from SOM around the world” [Jackson 2017: 422], with a view toward developing better understanding of processes that stabilize SOM. It explains the biological processes involved in carbon cycling and storage, finding that “root inputs are approximately five times more likely than an equivalent mass of aboveground litter to be stabilized as SOM” [Jackson 2017: 420]. Litter input can either increase or decrease SOM, despite the assumption in most carbon models that there is a linear relationship between litter input and transformation of carbon into more stable forms. This finding suggests that perennials and other deep-rooting plants have an important role to play with respect to carbon sequestration. As the author puts it:
Managing carbon inputs and relative allocation, for instance, through selection for deep roots or for greater belowground allocation in crops (Kell 2011), has been suggested as a way to increase SOM formation and stabilization in such systems (Bolinder et al. 2007, Eclesia et al. 2016). However, plant breeding has traditionally selected for aboveground yields alone; therefore, potential trade-offs between yield and root production must be carefully evaluated (DeHaan et al. 2005). New tools for monitoring root systems and in situ SOM in the field are needed (Molon et al. 2017) [Jackson 2017: 422]. . . .
The importance of root inputs for SOM formation is likely attributable to both their chemical composition and, almost certainly, their presence in the soil; upon death, they immediately interact with soil minerals, microbes, and aggregates. Roots tend to be characterized more by aliphatic compounds that are readily sorbed to mineral surfaces, and their composition (and that of root exudates) can increase microbial carbon use efficiency (CUE), defined as the ratio of microbial growth to carbon uptake, more than litter can. High CUE promotes microbial growth and carbon stabilization in mineral-associated soil pools, and low CUE favors biomass respiration (Manzonietal.2012a) [Jackson 2017: 423]. . . .
Soils hold the largest biogeochemically active terrestrial carbon pool on Earth and are critical for stabilizing atmospheric CO2 concentrations. Nonetheless, global pressures on soils continue from changes in land management, including the need for increasing bioenergy and food production [Jackson 2017: 420].
. . . plant breeding has traditionally selected for aboveground yields alone; therefore, potential trade-offs between yield and root production must be carefully evaluated[Jackson 2017].
National comparison of the total and sequestered organic matter contents of conventional and organic farm soils, Ghabbour 2017
An analysis of hundreds of soil samples collected from organic and conventional farms around the US shows higher average percentages both of total SOM and of humic substances – a measure of carbon sequestration – for organic farm soils compared to conventional farm soils. The mean percent humification (humic substances divided by total SOM) for organic soils is 57.3%, compared to 45.6% for conventional soils.
Agroforestry strategies to sequester carbon in temperate North America, Udawatta & Jose 2012
This meta-analysis estimates total carbon sequestration potential in the US from various agroforestry practices to be 530 TgC/year (530 million metric tons), equivalent to about 1/3 of annual US carbon emissions from fossil fuel combustion. Based on their literature review, the authors estimate per-hectare sequestration rates (based on aboveground and belowground carbon accumulation) for each practice as follows: 6.1t C/ha/yr (silvopastoral), 3.4t C/ha/yr (alleycropping), 6.4t C/ha/yr (windbreaks), 2.6t C/ha/yr (riparian buffer).
Compost, manure and synthetic fertilizer influences crop yields, soil properties, nitrate leaching and crop nutrient content, Hepperly 2009
A sequestration rate of 2.363t C/ha/yr was demonstrated where compost made of dairy manure and leaves was applied to fields in a three year rotation of corn-vegetable-small grain, with leguminous cover crops. The same rotation treated with chemical fertilizer instead of compost resulted in a net loss of -0.317t C/ha/yr.
Legume-based cropping systems have reduced carbon and nitrogen losses, Drinkwater 1998
This study compared three corn-soybean cropping systems: (1) conventional 2-yr rotation with chemical inputs, and residues returned to soil; (2) a longer (than 2 years), organic rotation with grass/legume hayed and returned to soil in manure; and (3) a longer (than 2 years) organic rotation with grass/legume turned back into the soil directly. Even though the conventional system returned more total residue to the soil, carbon sequestration was significantly lower for the conventional system than for the two organic, legume-based systems. Authors suggest that this is due to greater temporal plant diversity from the longer rotations, and higher quality residue (greater N:C) in the two legume-based organic systems. Furthermore, CO2 emissions were lower in the legume-based organic systems due to 50% lower energy use.
Even though the conventional system returned more total residue to the soil, carbon sequestration was significantly lower for the conventional system than for the two organic, legume-based systems. Authors suggest that this is due to greater temporal plant diversity from the longer rotations, and higher quality residue (greater N:C) in the two legume-based organic systems. [Drinkwater]
Agricultural production must produce enough food for almost 10 billion people by 2050 [FAO 2017],[10] and yet a third of all land is degraded [FAO 2015] and nearly all agricultural land has lost significant amounts of SOC (Soil Organic Carbon). So we have a puzzle to solve: how to produce more from less, and in the face of a more chaotic climate system. Between 1960 and 1990, the increased use of synthetic fertilizers, pesticides, irrigation, and modern seed varieties nearly doubled world cereal yield [FAO 1996]. Because of this apparent success, it’s not unreasonable to think the answer to the puzzle involves the same combination.
However, we now face a convergence of extremely dangerous crises – global warming, cascading species extinctions, antibiotic resistance, and ubiquitous chemical and nutrient pollution – all of which are aggravated by the current fossil-fuel-intensive industrial model of agriculture. Indeed, the current food system accounts for nearly 30% of total greenhouse gas emissions [Vermeulen 2012]. Furthermore, nearly half of harvested crops are lost because it is thrown away before being eaten or due to overconsumption (food consumption in excess of nutritional requirements) [Alexander 2017]. In spite of this, 11% of the world’s people are still hungry [FAO 2017].
Yet, for each agricultural problem, there is a known solution. For instance, agriculture has the potential to be a carbon sink. Many scientists have found that implementing various conservation practices can sequester up to 1 ton of carbon per hectare per year (1t C/ha/yr), or an estimated ~20% global emissions offset if such practices were broadly implemented [Smith 2008, Lal 2016]). Yet others have measured sequestration rates upwards of 8t C/ha/yr. While climate and soil type play a role in the variation among sequestration rates, farming practices are a major factor.
Studies with lower sequestration rates tend to isolate just one or two soil-building practices. For example, Minasny et al. [2017] compiled sequestration rates from around the world to assess the viability of the France-led “4 per 1000” initiative (seeking to offset the annual increase in atmospheric CO2 by increasing soil carbon by 0.4% per year). The authors estimate that an annual rate of 0.2-0.5t C/ha/yr “is possible after adoption of best management practices on arable land such as reduced tillage in combination with leguminous cover crops” [Minasny 2017: 61]. However, most of some 40 studies of best management practices assessed only one or two practices, often minimally improved, such as “reduced use of summer fallow,” “rice-rice with NPK,” “inorganic fertilizer,” and “pasture” (without mention of how the pasture was managed) [Minasny 2017: 64]. In other words, the “improved” practices here include even the use of synthetic fertilizer, which can generate more crop biomass and thus more residue, but has also been shown to diminish soil organic carbon [Khan 2007].
By contrast, researchers in New Mexico [Johnson 2017] recorded an annual carbon sequestration rate of 10.7t C/ha/yr from fungal-dominant compost in a 4.5-year trial, and they estimate a potential rate of 19.2t C/ha/yr. Chief investigator David Johnson found that increased plant growth is closely correlated with the fungal to bacterial ratio of the soil. Similarly, Machmuller et al. [2015] measured carbon sequestration rates in the southeastern United States of 8t C/ha/yr following conversion of row crop agriculture to management-intensive grazing, leading to an approximately 75% increase in soil carbon within six years.
The studies showing higher sequestration rates reveal what many farmers already know: that it takes not just one, but multiple regenerative practices, to really build soil organic matter (SOM). California Farmers Paul and Elizabeth Kaiser, for instance, use 5-10 times more compost than average, never till, rotate fields with an extremely diverse mix of vegetable varieties, surround their crops with native trees, shrubs and flowers and have thus built up a thick topsoil containing 10% SOM [Oppenheimer 2015; Kaiser 2017]. North Dakota Farmer Gabe Brown began practicing no-till in 1994. Since then, he has added cover crops (a diverse mix of 70 species), complex crop rotations, orchards, livestock grazing (including cattle, sheep, pork and chicken), vegetable production, and bees. By limiting soil disturbance and favoring biodiversity, Brown reports SOM increased from 1.7% in 1993 to as high as 11% in 2013. Over the same period, rainfall infiltration has increased from ½ inch per hour to more than 15 inches per hour [Brown 2016].
Looking at the big picture, researchers at the Rodale Institute [2014] estimate that if all cropland were converted to a regenerative model, it would sequester 40% of annual CO2 emissions. Adding regeneratively managed pastures to the picture would add another 71%, effectively exceeding the world’s yearly carbon dioxide emissions. Teague et al [2016] came up with similar results, estimating that regenerative conservation cropping and adaptive multi-paddock grazing can turn North American agricultural soils from a carbon source in conventional agriculture into a carbon sink at rate of ~3t C/ha/yr. Key factors include the use of no-till, diversified crop rotation, cover crops, organic soil amendments and reducing use of nitrogen (N) fertilizer.
The billion-dollar question, though, remains: can regenerative agriculture feed the world’s ever-expanding population? There is considerable evidence that it can, and in fact the opposite question (can the current model of industrial agriculture feed the world?) deserves at least as much scrutiny, given that so far the answer has been no.
In 2009, a multi-stakeholder team of hundreds of people from every region of the world released the International Assessment of Agricultural Knowledge Science and Technology for Development [IAASTD 2009], which provides a framework for a new global approach to agriculture. It poses the question: how can agricultural knowledge, science and technology (AKST) “be used to reduce hunger and poverty, improve rural livelihoods, and facilitate equitable … sustainable development” [IAASTD 2009: 3] in a global context of mounting social inequity, poverty, human migration, biodiversity loss, and climate change, among other concerns.
Their answer hinges on the concept of multifunctionality of agriculture, or “the challenge … to simultaneously meet development and sustainability goals while increasing agricultural production” [IAASTD 2009: 4]. It calls for a “fundamental shift” in AKST that recognizes “farming communities, farm households, and farmers as producers and managers of ecosystems” [IAASTD 2009: 4], and values both scientific research and traditional and local knowledge. With a focus on multifunctionality,
AKST can contribute to radically improving food security and enhancing the social and economic performance of agricultural systems as a basis for sustainable rural and community livelihoods and wider economic development. It can help to rehabilitate degraded land, reduce environmental and health risks associated with food production and consumption and sustainably increase production [IAASTD 2009: 5].
Thinking of agriculture as multifunctional means valuing agricultural land not only in terms of its capacity for maximum output, but also for its vital role in providing wildlife habitat, sequestering carbon, absorbing and storing rainfall, recycling nutrients, providing for nutritionally balanced diets, and providing the means for an adequate livelihood in farming. And there is abundant evidence that not only do regenerative methods provide these multiple services, they can also be as productive as fossil-fuel-intensive methods, and even more so in times of drought. A 22-year Pennsylvania study [Pimentel 2005] comparing the productivity of conventional versus organic systems showed that while corn yields were comparable overall, during five dry years of the study the organic systems were 28% to 34% more productive than their conventional counterparts.
Thinking of agriculture as multifunctional means valuing agricultural land not only in terms of its capacity for maximum output, but also for its vital role in providing wildlife habitat, sequestering carbon, absorbing and storing rainfall, recycling nutrients, providing for nutritionally balanced diets, and providing the means for an adequate livelihood in farming.
Similarly, a nine-year Iowa study [Liebman 2013] comparing corn and soybean yields in 2-year, 3-year, and 4-year rotations resulted in higher yields from the more diverse 3-yr and 4-yr rotation systems than for the conventional 2-yr system, despite substantial reductions in the use of synthetic N fertilizer, herbicides, and fossil-fuel energy in the longer rotations. The longer rotations also incorporated cover crops and manure fertilizer at planting time, as opposed to the 2-year rotation, which incorporated only synthetic fertilizer, and involved no cover cropping.
These results run counter to other studies that have reinforced the belief that industrial agriculture is necessarily more productive. The authors of the Rodale report offer an explanation for this apparent contradiction:
Meta-analyses of refereed publications show that, on average, organic yields are often lower than conventional. But the yield gap is prevalent when practices used in organic mimic conventional, that is, when the letter of organic standards is followed using an input mentality akin to conventional chemical-intensive agriculture [Rodale 2014: 15].
In other words, a more fundamental commitment to regenerative methods is necessary to bring productivity up to par with that of conventional/industrial methods. Removing the inputs that undergird the success of industrial agriculture will cause the system to falter unless the nutrients and pest resistance provided by those inputs are replaced by that which is proffered through healthy, living soil ecosystems. Indeed, the transition period from industrial system to regenerative system is typically characterized by a drop in yield until the previously damaged soil has come back to life.
Healthy, carbon-rich soil is a powerful engine for plant growth, thanks in large part to the presence of billions of microorganisms working in concert with plants. This includes bacteria living in the roots of leguminous plants that fix plant-available nitrogen, other bacteria defending plants against disease, hyphae-forming fungi attaching tothe end of roots and effectively extending those roots deeper into the soil to retrieve micro-nutrients. There are fungi that mine otherwise unavailable soil phosphorus and deliver it to plants, and fungi that help to build the structure of the soil by binding clumps of soil together in aggregates. This aggregation, in turn, facilitates soil aeration and water-holding capacity, while also holding soil organic carbon in place.
It is ironic that in the process of delivering synthetic inputs to feed plants and prevent disease, industrial methods destroy the microorganisms and soil structure that would otherwise serve these purposes. However, for soil to perform these functions in the context of agricultural production requires thoughtful management aiming to protect soil and recycle nutrients. Harvesting a crop removes nutrients from a system, which then need to be replaced. In a wild ecosystem, nutrients consumed are generally replaced by plant litter and animal waste. Nutrients are also made available to plants in healthy, undisturbed soils, by microbial action on mineral particles and rocks. Regenerative agricultural systems mimic this nutrient cycling through compost and manure application, cover cropping, and no-till (which protects the soil, allowing microorganisms to flourish). Moreover, regenerative agriculture mimics wild systems by striving for biodiversity, which improves the stability and productivity of ecosystems.
Also challenging is the fact that a majority of researchers demonstrating climate mitigation through better agricultural conservation practices (Minasny 2017, Zomer 2017, Grimson 2017, Lal 2016, Smith 2008, West & Post 2002, for example) stop short of considering carbon sequestration from a full suite of conservation practices representing a more fundamental commitment to regenerative agriculture. Making only minimal conservation improvements while maintaining the same industrial system tends to result not only in minimal sequestration outcomes, but also lower yields. As noted above, this in-between approach to agriculture risks sacrificing the strengths tendered by a full suite of practices, whether from the industrial or the ecological model.
Murmurings from every corner of the globe reveal that many already acknowledge the imperative, especially for wealthy countries, to radically change our way of living, including the design of our agricultural systems. The vision for societal change varies from cutting fossil fuel emissions to zero within a decade; to building a commons-oriented de-growth economy that values life and sharing over the hoarding of material wealth; to transitioning to a community-based, equitable, agro-ecological food-system.
Specifically responding to the question of how to feed more people with less, like a riddle from the Sphinx, requires thinking outside the box and acknowledging our imperative to radically change ourselves. It’s not enough simply to improve the current industrial agricultural model if, at best, that merely slows progress toward climate tipping points (when the positive feedback loops accelerate global warming beyond our control). Tweaking the current system with small improvements toward inadequate goals is what Margaret Klein Salamon calls “climate gradualism.” For all its political practicality, this approach is like putting a band-aid on a severed limb, and is irrational from the perspective of preserving human civilization in the face of climate breakdown.
In our technophilic society, it may come as a surprise that building biologically active, carbon-rich soil is the answer both to the climate crisis and to the question of how to feed more with less. While protecting and rehabilitating wild ecosystems are also essential, rebuilding the soil in agricultural lands through a regenerative, multifunctional approach is arguably the key to protecting human civilization.
In our technophilic society, it may come as a surprise that building biologically active, carbon-rich soil is the answer both to the climate crisis and to the question of how to feed more with less.
The articles that follow are a small sampling of many recent studies examining various aspects of agricultural ecosystems and food production in the face of climate breakdown. Each provides a glimpse into the range and depth of nature-based tools at our fingertips to transform agriculture into a wellspring of planetary resilience.
Compilation of agriculture articles
Natural climate solutions, Griscom 2017
This is one of the most comprehensive mainstream studies to date of a broad spectrum of natural climate solutions by thirty-two co-authors and supported by The Nature Conservancy. The report examines “20 conservation, restoration, and/or improved land management actions that increase carbon storage and/or avoid greenhouse gas emissions across global forests, wetlands, grasslands, and agricultural lands.” The authors “find that the maximum potential of NCS [Natural Climate Solutions] —when constrained by food security, fiber security, and biodiversity conservation—is 23.8 petagrams[11] of CO2 equivalent . . . This is ≥30% higher than prior estimates, which did not include the full range of options and safeguards considered here.” [Griscom 2017: 11645]. The study seeks to assess both the potential emissions from land use as well as the carbon-sequestration potential.
The study posits a target of <2o C as the conventionally agreed-upon safe limit:
Warming will likely be held to below 2 °C if natural pathways are implemented at cost-effective levels . . . and if we avoid increases in fossil fuel emissions for 10 y and then drive them down to 7% of current levels by 2050 and then to zero by 2095 [p. 11647]
The authors state that their estimates are intentionally conservative because (1) they do not include potential benefits of payments for high-money-value ecosystem services in stimulating NCS efforts; (2) they exclude various management practices where data were not “sufficiently robust for global extrapolation,” e.g., no-till, adaptive multi-paddock grazing, etc.; and (3) significant additional investment would be required to keep warming at 1.5o C. [Griscom 2017: 11648]
Detail is provided on contributions of specific mitigation pathways, such as forests, wetlands, grasslands, etc., and on challenges as well. For example, “Despite the large potential of NCS, land-based sequestration efforts receive only about 2.5% of climate mitigation dollars.” [Griscom 2017: 11648] This observation is consistent with our observations of limited available resources for the most basic NCS education. Other challenges include deforestation for farming and animal husbandry, losing high carbon sequestration benefits of wetlands due to reclamation, and impacts of climate feedbacks such as fire, drought, temperature increases, etc.
We applaud Griscom et al. for an excellent and comprehensive analysis and review of many of the factors in natural climate solutions. We do, however, believe that (1) the potential of nature’s solutions is far greater than Griscom et al. estimate, and (2) that the temperature limits (1.5o – 2o C) are too high and too dangerous – considering that natural processes are already changing, drastically and for the worse, with an average global temperature increase of barely 1o C (see Appendix A: Urgency of the Biodiversity and Climate Crisis).
The differences between the perspectives of Griscom et al. and those adopted in this Compendium are paradigmatic. Griscom et al. acknowledges that their estimates are conservative, looks at a set of studies that tends toward the mainstream and is primarily based on established and widespread practice. This is perfectly reasonable in the process of what Thomas Kuhn calls “normal science” (see Compendium Vol. 1 No. 1 for an extensive discussion of Kuhn’s landmark work). Unfortunately the process of normal science for accepting new thinking and discoveries usually takes decades, and we are currently in the throes of an extinction, and an emergency with respect to biodiversity, and climate change. Therefore we have to accelerate our response. Accordingly, Bio4Climate searches for studies that tend to examine positive variants, i.e., examples of what is possible beyond current conceptual boundaries. We emphasize goals to strive for, even if the data are not yet “sufficiently robust for global extrapolation.” The robustness of such data will increase with more intentional focus.
An interesting side effect of the paradigm difference is that numerous sources that we cite, many from the scientific literature, don’t appear in NCS references (for example, Richard Teague [Teague et al. 2016], Gabe Brown [Brown 2016], Tom Goreau [Goreau 2015], Rebecca Ryals and Whendee Silver [Ryals and Silver 2013], David Johnson [Johnson 2017], Paul and Elizabeth Kaiser [Kaiser 2017], Terry McCosker [McCosker 2000], Carol Evans and Jon Griggs [Evans et al., 2015], to name just a few). Nor are there discussions of permaculture or agroforestry, two of the more promising areas of research and practice in land management that lead to climate-positive results.
Unfortunately the process of normal science for accepting new thinking and discoveries usually takes decades, and we are currently in the throes of an extinction, and an emergency with respect to biodiversity, and climate change. Therefore we have to accelerate our response. Accordingly, Bio4Climate searches for studies that tend to examine positive variants, i.e., examples of what is possible beyond current conceptual boundaries. We emphasize goals to strive for, even if the data are not yet “sufficiently robust for global extrapolation.” The robustness of such data increases with intentional focus.
Drawdown, Hawken, ed. 2017
Edited by innovator and entrepreneur Paul Hawken, Drawdown is a remarkable and comprehensive work presenting eighty well-vetted solutions and twenty promising “coming attractions” to remove carbon from the atmosphere and restore planetary health. Hawken engaged numerous scientists, modellers, advisers, artists and writers, resulting in a beautifully illustrated and comprehensive exploration of possibilities for reversing global warming.
The impact of the book as a whole is as important as each solution: Drawdown presents a universe of actions that go far beyond what we can imagine if we consider only emissions reductions and alternative energy. It leads to an entirely different climate conversation from the one we’re used to, and offers many threads of hope.
Drawdown has something for everyone, covering sectors of Energy, Food, Buildings and Cities, Land Use, Transport, and Materials. Near the top of the list is Women and Girls, whose education has dramatic effects on population and is one of the most important climate positive steps we can take. Of course technology offerings abound, but they are amply balanced by discussions of biology and social change, often sorely missing in debates on global warming. Of particular interest in this Compendium are biological strategies; we’ll mention just three of them here.
Agroforestry in Burkina Faso
After terrible droughts in the 1980s resulted in a 20% reduction in rainfall and millions of deaths by starvation, farmer Yacouba Sawadogo enhanced a traditional practice of digging rain-capturing pits by adding manure. There were seeds in the manure and as a result trees began to grow, holding soils together with roots, protecting plantings from wind gusts that before had required frequent re-sowing, and opening channels that moved water into the soils and raised water tables. This foray into agro-forestry spread across the rural countryside to widespread beneficial effect.
Of great significance is that the expertise, invention and community organizing were native and local, required no foreign aid or expensive soil inputs, and in terms of money cost nothing. This is what sustainability can look like. [Hawken 2017: 118-120]
Pasture Cropping
In 1979, after a devastating fire destroyed his two-thousand acre farm in Australia, Colin Seis began to question why crops and animals couldn’t be profitably raised on the same land, effectively doubling output. Persisting through a difficult transition, Seis saw water retention improvements, decreased input costs, a virtual end to insect infestation, and measures of soil fertility and carbon content go up along with profits. Today, pasture cropping is practiced on over two thousand Australian farms and is spreading throughout the world. [Hawken 2017: 175]
Intensive Silvopasture
Silvopasture, the most common form of agroforestry, is the practice of combining trees and woody shrubs with pasture grasses. The result is healthier plant and animal growth, including sequestering a respectable one to four tons of carbon per acre. It is currently practiced on over one billion acres worldwide.
For remarkable next steps enter the intensive part of silvopasture, starting with a quickly growing, edible leguminous shrub, Leucaena leucocephala in Australia and Latin America (different species of shrubs are suitable in different ecosystems). Water retention improves, biomass increases, species biodiversity doubles, animal stocking rates almost triple, ambient temperatures decrease by 14 to 23 degrees F in the tropics, meat production increases by a factor of 4 to 10, and perhaps most strikingly, soil carbon sequestration rates have exceeded 10 tons per acre (conventional agriculture can claim 1 ton of carbon per acre or less, or even net carbon loss to the atmosphere). [Hawken 2017: 181]
Intertidal resource use over millennia enhances forest productivity, Trant 2016
Abstract: Human occupation is usually associated with degraded landscapes but 13,000 years of repeated occupation by British Columbia’s coastal First Nations has had the opposite effect, enhancing temperate rainforest productivity. This is particularly the case over the last 6,000 years when intensified intertidal shellfish usage resulted in the accumulation of substantial shell middens. We show that soils at habitation sites are higher in calcium and phosphorous. Both of these are limiting factors in coastal temperate rainforests. Western red cedar (Thuja plicata) trees growing on the middens were found to be taller, have higher wood calcium, greater radial growth and exhibit less top die-back. Coastal British Columbia is the first known example of long-term intertidal resource use enhancing forest productivity and we expect this pattern to occur at archaeological sites along coastlines globally [Trant 2016: 1].
Although focused on forests and not farmland, this study shows that, as in the Amazon, where indigenous people created SOM-rich terra preta soil (akin to biochar-enhanced soil), human populations can increase soil quality and ecosystem productivity beyond what the potential would have been absent human activity.
This is an interesting point with respect to global potential for soil carbon sequestration. Scientists often refer to an equilibrium point, up to which soils can regain carbon previously lost through exploitive human activity. Equilibrium is generally seen as being the point at which new SOC levels are equivalent to or somewhat less than what they were prior to human exploitation of the soil, and never greater than the original amount. While Trant et al. [2016] have found evidence of calcium and phosphorus (not carbon) enrichment due to human activity, their findings raise questions about the extent to which intentionally building soils through all the methods we know to maximize carbon storage could increase various soils’ presumed equilibrium points.
Human occupation is usually associated with degraded landscapes but 13,000 years of repeated occupation by British Columbia’s coastal First Nations has had the opposite effect, enhancing temperate rainforest productivity [Trant 2016: 1].
The ecology of soil carbon: pools, vulnerabilities, and biotic and abiotic controls, Jackson 2017
This review examines “the state of knowledge for the stocks of, inputs to, and outputs from SOM around the world” [Jackson 2017: 422], with a view toward developing better understanding of processes that stabilize SOM. It explains the biological processes involved in carbon cycling and storage, finding that “root inputs are approximately five times more likely than an equivalent mass of aboveground litter to be stabilized as SOM” [Jackson 2017: 420]. Litter input can either increase or decrease SOM, despite the assumption in most carbon models that there is a linear relationship between litter input and transformation of carbon into more stable forms. This finding suggests that perennials and other deep-rooting plants have an important role to play with respect to carbon sequestration. As the author puts it:
Managing carbon inputs and relative allocation, for instance, through selection for deep roots or for greater belowground allocation in crops (Kell 2011), has been suggested as a way to increase SOM formation and stabilization in such systems (Bolinder et al. 2007, Eclesia et al. 2016). However, plant breeding has traditionally selected for aboveground yields alone; therefore, potential trade-offs between yield and root production must be carefully evaluated (DeHaan et al. 2005). New tools for monitoring root systems and in situ SOM in the field are needed (Molon et al. 2017) [Jackson 2017: 422]. . . .
The importance of root inputs for SOM formation is likely attributable to both their chemical composition and, almost certainly, their presence in the soil; upon death, they immediately interact with soil minerals, microbes, and aggregates. Roots tend to be characterized more by aliphatic compounds that are readily sorbed to mineral surfaces, and their composition (and that of root exudates) can increase microbial carbon use efficiency (CUE), defined as the ratio of microbial growth to carbon uptake, more than litter can. High CUE promotes microbial growth and carbon stabilization in mineral-associated soil pools, and low CUE favors biomass respiration (Manzonietal.2012a) [Jackson 2017: 423]. . . .
Soils hold the largest biogeochemically active terrestrial carbon pool on Earth and are critical for stabilizing atmospheric CO2 concentrations. Nonetheless, global pressures on soils continue from changes in land management, including the need for increasing bioenergy and food production [Jackson 2017: 420].
. . . plant breeding has traditionally selected for aboveground yields alone; therefore, potential trade-offs between yield and root production must be carefully evaluated[Jackson 2017].
National comparison of the total and sequestered organic matter contents of conventional and organic farm soils, Ghabbour 2017
An analysis of hundreds of soil samples collected from organic and conventional farms around the US shows higher average percentages both of total SOM and of humic substances – a measure of carbon sequestration – for organic farm soils compared to conventional farm soils. The mean percent humification (humic substances divided by total SOM) for organic soils is 57.3%, compared to 45.6% for conventional soils.
Agroforestry strategies to sequester carbon in temperate North America, Udawatta & Jose 2012
This meta-analysis estimates total carbon sequestration potential in the US from various agroforestry practices to be 530 TgC/year (530 million metric tons), equivalent to about 1/3 of annual US carbon emissions from fossil fuel combustion. Based on their literature review, the authors estimate per-hectare sequestration rates (based on aboveground and belowground carbon accumulation) for each practice as follows: 6.1t C/ha/yr (silvopastoral), 3.4t C/ha/yr (alleycropping), 6.4t C/ha/yr (windbreaks), 2.6t C/ha/yr (riparian buffer).
Compost, manure and synthetic fertilizer influences crop yields, soil properties, nitrate leaching and crop nutrient content, Hepperly 2009
A sequestration rate of 2.363t C/ha/yr was demonstrated where compost made of dairy manure and leaves was applied to fields in a three year rotation of corn-vegetable-small grain, with leguminous cover crops. The same rotation treated with chemical fertilizer instead of compost resulted in a net loss of -0.317t C/ha/yr.
Legume-based cropping systems have reduced carbon and nitrogen losses, Drinkwater 1998
This study compared three corn-soybean cropping systems: (1) conventional 2-yr rotation with chemical inputs, and residues returned to soil; (2) a longer (than 2 years), organic rotation with grass/legume hayed and returned to soil in manure; and (3) a longer (than 2 years) organic rotation with grass/legume turned back into the soil directly. Even though the conventional system returned more total residue to the soil, carbon sequestration was significantly lower for the conventional system than for the two organic, legume-based systems. Authors suggest that this is due to greater temporal plant diversity from the longer rotations, and higher quality residue (greater N:C) in the two legume-based organic systems. Furthermore, CO2 emissions were lower in the legume-based organic systems due to 50% lower energy use.
Even though the conventional system returned more total residue to the soil, carbon sequestration was significantly lower for the conventional system than for the two organic, legume-based systems. Authors suggest that this is due to greater temporal plant diversity from the longer rotations, and higher quality residue (greater N:C) in the two legume-based organic systems. [Drinkwater]
Testing the effects of ants and termites on crop yield in an arid part of Australia, this study showed “that ants and termites increase wheat yield by 36% from increased soil water infiltration due to their tunnels and improved soil nitrogen” [Evans 2011: 1]. The authors conclude: “Our results suggest that ants and termites have similar functional roles to earthworms, and that they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates” [Evans 2011: 1].
Ants and termites have similar functional roles to earthworms, and . . . they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates [Evans 2011: 1].
Termites are particularly important in savannas of Africa, Australasia, and South America, and their nest structures (“mounds”) shape many environmental properties; analogous structures built by ants and burrowing mammals are similarly influential worldwide. Mound soils differ from surrounding “matrix” soils in physical and chemical composition, which enhances vegetation growth, creating “islands of fertility.” Moreover, mounds are frequently spatially over-dispersed owing to competition among neighboring colonies, which creates spotted vegetation patterns [Bonachela 2015: 652].
This study seeks to characterize landscape patterns created by termites in order to distinguish between that and other causes of spotted vegetation patterns that have been assumed to indicate imminent ecological collapse. “Rather, mound-field landscapes are more robust to aridity, suggesting that termites may help stabilize ecosystems under global change” [Bonachela 2015: 651].
“Dung beetles (Scarabaeidae: Scarabaeinae, Aphodiinae, Geotrupidae) are some of the most important invertebrate contributors to dung decomposition in both temperate and tropical agricultural grasslands. As such, they may help mitigate GHG [Greenhouse Gas] emissions and aid carbon sequestration through removing dung deposited on the pastures, increasing grass growth and fertilization” [Slade 2015: 1]. This Finland study analyzed the percent of GHGs removed by dung beetles at three levels: dung pat, pasture, and dairy/beef production life-cycle, finding reduced GHG emissions of 7%, 12%, and 0.05 to 0.13%, respectively. Dung beetles reduce methane emissions by aerating the dung pats, thereby preventing methane-producing anaerobic decomposition of the dung.
The reason dung beetles have a minimal effect in the full life-cycle analysis for Finland cattle is that the animals spend only a short portion of the year grazing in pasture, and thus emissions from dung on pasture is “dwarfed in comparison to other emissions of milk and meat production, such as methane emissions from enteric fermentation, nitrous oxide emissions from soils, and carbon dioxide emissions from energy use” [Slade 2015: 5]. However, “in regions where outdoor livestock grazing is more commonly used, the emissions from manure left on pasture will have a larger contribution to total agricultural emissions, with estimated fractions ranging from 11% in Asia up to 35% in Africa. Such patterns are combined with likely differences in dung beetle efficiency: In tropical regions, dung beetles can remove the majority of a fresh dung pat within the first few days after deposition – whereas in temperate conditions, a substantial fraction will remain throughout the grazing season” [Slade 2015: 5].
The authors recommend further research in tropical regions, predicting: “that effects at all levels from dung pats through pastures to the whole lifecycle of milk or beef production may be strongly accentuated at low latitudes” [Slade 2015: 5].
Baleen and sperm whales, known collectively as the great whales, include the largest animals in the history of life on Earth. With high metabolic demands and large populations, whales probably had a strong influence on marine ecosystems before the advent of industrial whaling: as consumers of fish and invertebrates; as prey to other large-bodied predators; as reservoirs and vertical and horizontal vectors for nutrients; and as detrital sources of energy and habitat in the deep sea. The decline in great whale numbers, estimated to be at least 66% and perhaps as high as 90%, has likely altered the structure and function of the oceans, but recovery is possible and in many cases is already underway. Future changes in the structure and function of the world’s oceans can be expected with the restoration of great whale populations.
Testing paired sites in four ecosystem types, this study finds that higher fungal activity in soil is associated with higher soil carbon content, and that disturbing the soil reduces fungal activity. The paper’s introduction explains why fungi have been found to store more carbon than do bacteria – for example, fungi can store up to 26 times more carbon from leaf litter than bacteria. This is because the chemical composition of fungal biomass is more complex and more resistant to degradation; also, fungi have higher carbon assimilation efficiencies than do bacteria, and thus store more of the carbon they metabolize.
A variety of intimate, symbiotic relationships exist between the roots of plants and the microorganisms in the soil. For instance, mycorrhizal fungi colonize the surface of plant roots, effectively extending them further through the soil to collect nutrients otherwise out of reach. These mycorrhizal branching structures, known as hyphae, emanating from plant roots also improve soil aggregation and hence improve water infiltration and aeration. In return, Mycorrhiza can demand up to 20-40% of photosynthetically derived carbon from their plant hosts. In the world of rhizospheric bacteria, Rhizobia[9] are well known for their key role in fixing atmospheric nitrogen for plant uptake. Yet there are, additionally, more than two dozen known genera of rhizobacteria that help plants grow, either directly by releasing growth stimulants (phytohormones) and enhancing mineral uptake, or indirectly by fighting off plant pathogens.
Fungus is a key component of healthy soil. It is known to “translocate compounds from nutrient-rich to nutrient-poor regions… facilitate the access of bacteria to suitable microhabitats for growth, enable efficient contaminant biodegradation, and increase the functional stability in systems exposed to osmotic stress” [Berthold 2016: 5]. This study shows that, in addition, mycelia facilitate bacterial evolution, thereby bolstering bacterial diversity and adaptability.
Abstract: Horizontal gene transfer (HGT) is a main mechanism of bacterial evolution endowing bacteria with new genetic traits. The transfer of mobile genetic elements such as plasmids (conjugation) requires the close proximity of cells. HGT between genetically distinct bacteria largely depends on cell movement in water films, which are typically discontinuous in natural systems like soil. Using laboratory microcosms, a bacterial reporter system and flow cytometry, we here investigated if and to which degree mycelial networks facilitate contact of and HGT between spatially separated bacteria. Our study shows that the network structures of mycelia promote bacterial HGT by providing continuous liquid films in which bacterial migration and contacts are favoured. This finding was confirmed by individual-based simulations, revealing that the tendency of migrating bacteria to concentrate in the liquid film around hyphae is a key factor for improved HGT along mycelial networks. Given their ubiquity, we propose that hyphae can act as a focal point for HGT and genetic adaptation in soil.
Fossil evidence shows that early land plants hosted fungal symbionts, which are likely to have facilitated phosphorus acquisition by plants and thus increased net primary production, perpetuating the transition to a cooler, oxygen-rich environment suitable for animal life. Mills’ study tests this hypothesis by integrating plant-fungal phosphorus acquisition into a biogeochemical model of the Paleozoic climate transition. The study finds “significant Earth system sensitivity to phosphorus uptake from mycorrhizal fungi” [Mills 2017: 7], and that “efficient phosphorus uptake at superambient CO2 results in enhanced carbon sequestration, which contributes to a reduction in CO2 and drives a rise in O2” [Mills 2017: 6].
Understanding drivers of an historic climate cooling is obviously relevant today given current atmospheric CO2 accumulation. This study points to the importance of plant-fungal symbioses and phosphorus cycling, and thus to the importance of building and protecting soil health to allow such symbioses to flourish.
The articles below offer a sampling of the myriad ecosystem roles played by species we may not think much about. For example, fungi, an exemplar ecosystem cooperator, buries carbon in the soil, sources otherwise unavailable nutrients like phosphorus for plant growth, and facilitates bacterial evolution. Great whales transport nutrients through the ocean for other species to consume. Dung beetles reduce methane emissions from manure, while also fertilizing grasses. Termites and ants promote vegetation growth in arid climates by creating tunnels that catch and hold rainwater, and by making nutrients available to plants.
Fossil evidence shows that early land plants hosted fungal symbionts, which are likely to have facilitated phosphorus acquisition by plants and thus increased net primary production, perpetuating the transition to a cooler, oxygen-rich environment suitable for animal life. Mills’ study tests this hypothesis by integrating plant-fungal phosphorus acquisition into a biogeochemical model of the Paleozoic climate transition. The study finds “significant Earth system sensitivity to phosphorus uptake from mycorrhizal fungi” [Mills 2017: 7], and that “efficient phosphorus uptake at superambient CO2 results in enhanced carbon sequestration, which contributes to a reduction in CO2 and drives a rise in O2” [Mills 2017: 6].
Understanding drivers of an historic climate cooling is obviously relevant today given current atmospheric CO2 accumulation. This study points to the importance of plant-fungal symbioses and phosphorus cycling, and thus to the importance of building and protecting soil health to allow such symbioses to flourish.
Mycelia as a focal point for horizontal gene transfer among soil bacteria, Berthold 2016
Fungus is a key component of healthy soil. It is known to “translocate compounds from nutrient-rich to nutrient-poor regions… facilitate the access of bacteria to suitable microhabitats for growth, enable efficient contaminant biodegradation, and increase the functional stability in systems exposed to osmotic stress” [Berthold 2016: 5]. This study shows that, in addition, mycelia facilitate bacterial evolution, thereby bolstering bacterial diversity and adaptability.
Abstract: Horizontal gene transfer (HGT) is a main mechanism of bacterial evolution endowing bacteria with new genetic traits. The transfer of mobile genetic elements such as plasmids (conjugation) requires the close proximity of cells. HGT between genetically distinct bacteria largely depends on cell movement in water films, which are typically discontinuous in natural systems like soil. Using laboratory microcosms, a bacterial reporter system and flow cytometry, we here investigated if and to which degree mycelial networks facilitate contact of and HGT between spatially separated bacteria. Our study shows that the network structures of mycelia promote bacterial HGT by providing continuous liquid films in which bacterial migration and contacts are favoured. This finding was confirmed by individual-based simulations, revealing that the tendency of migrating bacteria to concentrate in the liquid film around hyphae is a key factor for improved HGT along mycelial networks. Given their ubiquity, we propose that hyphae can act as a focal point for HGT and genetic adaptation in soil.
The rhizosphere - roots, soil and everything in between, McNear 2013
A variety of intimate, symbiotic relationships exist between the roots of plants and the microorganisms in the soil. For instance, mycorrhizal fungi colonize the surface of plant roots, effectively extending them further through the soil to collect nutrients otherwise out of reach. These mycorrhizal branching structures, known as hyphae, emanating from plant roots also improve soil aggregation and hence improve water infiltration and aeration. In return, Mycorrhiza can demand up to 20-40% of photosynthetically derived carbon from their plant hosts. In the world of rhizospheric bacteria, Rhizobia[9] are well known for their key role in fixing atmospheric nitrogen for plant uptake. Yet there are, additionally, more than two dozen known genera of rhizobacteria that help plants grow, either directly by releasing growth stimulants (phytohormones) and enhancing mineral uptake, or indirectly by fighting off plant pathogens.
Fungal to bacterial ratios in soils investigated for enhanced C-sequestration, Bailey 2002
Testing paired sites in four ecosystem types, this study finds that higher fungal activity in soil is associated with higher soil carbon content, and that disturbing the soil reduces fungal activity. The paper’s introduction explains why fungi have been found to store more carbon than do bacteria – for example, fungi can store up to 26 times more carbon from leaf litter than bacteria. This is because the chemical composition of fungal biomass is more complex and more resistant to degradation; also, fungi have higher carbon assimilation efficiencies than do bacteria, and thus store more of the carbon they metabolize.
Whales as marine ecosystem engineers, Roman 2014
Baleen and sperm whales, known collectively as the great whales, include the largest animals in the history of life on Earth. With high metabolic demands and large populations, whales probably had a strong influence on marine ecosystems before the advent of industrial whaling: as consumers of fish and invertebrates; as prey to other large-bodied predators; as reservoirs and vertical and horizontal vectors for nutrients; and as detrital sources of energy and habitat in the deep sea. The decline in great whale numbers, estimated to be at least 66% and perhaps as high as 90%, has likely altered the structure and function of the oceans, but recovery is possible and in many cases is already underway. Future changes in the structure and function of the world’s oceans can be expected with the restoration of great whale populations.
The role of dung beetles in reducing greenhouse gas emissions from cattle farming, Slade 2015
“Dung beetles (Scarabaeidae: Scarabaeinae, Aphodiinae, Geotrupidae) are some of the most important invertebrate contributors to dung decomposition in both temperate and tropical agricultural grasslands. As such, they may help mitigate GHG [Greenhouse Gas] emissions and aid carbon sequestration through removing dung deposited on the pastures, increasing grass growth and fertilization” [Slade 2015: 1]. This Finland study analyzed the percent of GHGs removed by dung beetles at three levels: dung pat, pasture, and dairy/beef production life-cycle, finding reduced GHG emissions of 7%, 12%, and 0.05 to 0.13%, respectively. Dung beetles reduce methane emissions by aerating the dung pats, thereby preventing methane-producing anaerobic decomposition of the dung.
The reason dung beetles have a minimal effect in the full life-cycle analysis for Finland cattle is that the animals spend only a short portion of the year grazing in pasture, and thus emissions from dung on pasture is “dwarfed in comparison to other emissions of milk and meat production, such as methane emissions from enteric fermentation, nitrous oxide emissions from soils, and carbon dioxide emissions from energy use” [Slade 2015: 5]. However, “in regions where outdoor livestock grazing is more commonly used, the emissions from manure left on pasture will have a larger contribution to total agricultural emissions, with estimated fractions ranging from 11% in Asia up to 35% in Africa. Such patterns are combined with likely differences in dung beetle efficiency: In tropical regions, dung beetles can remove the majority of a fresh dung pat within the first few days after deposition – whereas in temperate conditions, a substantial fraction will remain throughout the grazing season” [Slade 2015: 5].
The authors recommend further research in tropical regions, predicting: “that effects at all levels from dung pats through pastures to the whole lifecycle of milk or beef production may be strongly accentuated at low latitudes” [Slade 2015: 5].
Termite mounds can increase the robustness of dryland ecosystems to climatic change, Bonachela 2015
Termites are particularly important in savannas of Africa, Australasia, and South America, and their nest structures (“mounds”) shape many environmental properties; analogous structures built by ants and burrowing mammals are similarly influential worldwide. Mound soils differ from surrounding “matrix” soils in physical and chemical composition, which enhances vegetation growth, creating “islands of fertility.” Moreover, mounds are frequently spatially over-dispersed owing to competition among neighboring colonies, which creates spotted vegetation patterns [Bonachela 2015: 652].
This study seeks to characterize landscape patterns created by termites in order to distinguish between that and other causes of spotted vegetation patterns that have been assumed to indicate imminent ecological collapse. “Rather, mound-field landscapes are more robust to aridity, suggesting that termites may help stabilize ecosystems under global change” [Bonachela 2015: 651].
Ants and termites increase crop yield in a dry climate, Evans 2011
Testing the effects of ants and termites on crop yield in an arid part of Australia, this study showed “that ants and termites increase wheat yield by 36% from increased soil water infiltration due to their tunnels and improved soil nitrogen” [Evans 2011: 1]. The authors conclude: “Our results suggest that ants and termites have similar functional roles to earthworms, and that they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates” [Evans 2011: 1].
Ants and termites have similar functional roles to earthworms, and . . . they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates [Evans 2011: 1].
This study illustrates how ecosystem restoration enhances biodiversity and productivity. A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years, and without any human input after the original orange waste treatment of that site. The thick layer of orange peels suppressed existing non-native pasture grasses and added macro- and micronutrients to the soil, ultimately allowing for the natural (unmanaged) repopulating of the treated area from adjacent forest seedstock.
Our results provide nuance and detail to what was overwhelmingly obvious during informal surveys in 1999 and 2003: depositing orange waste on this degraded and abandoned pastureland greatly accelerated the return of tropical forest, as measured by lasting increases in soil nutrient availability, tree biomass, tree species richness, and canopy closure. The clear implication is that deposition of agricultural waste could serve as a tool for effective, low-cost tropical forest restoration, with a particularly important potential role at low-fertility sites [Treuer 2017: 6].
A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years [Treuer 2017].
This English grasslands study, comparing alternative strategies for increasing productivity, showed that “increasing plant species richness levels were more effective than the imposed levels of increasing management intensity” [Weigelt 2009: 1701]. The management intensification strategy included synthetic fertilization and mowing, while the biodiversity strategy increased species richness from 1 to 16 species. The authors conclude that:
For permanent grasslands, which cover one third of the utilised agricultural area in Europe (Smit et al., 2008), highly diverse communities composed of complementary species and N2-fixing legumes could provide an excellent agro-economic and ecological option for sustainable and highly productive grassland use [Weigelt 2009: 1704].
This study illustrates the importance of biodiversity for maintaining ecosystem stability. It tests the hypothesis that “other drivers of global environmental change may have biodiversity-mediated effects on ecosystem functioning – that changes in biodiversity resulting from anthropogenic drivers may be an intermediate cause of subsequent changes in ecosystem functioning” [Hautier 2015: 337]. Researchers found that “changes in plant diversity in response to anthropogenic drivers, including N, CO2, fire, herbivory[8], and water, were positively associated with changes in temporal stability of productivity,” and that “this positive association was independent of the nature of the driver” [Hautier 2015: 338]. In other words, the experimental interventions (N, CO2, fire, etc.) affected biodiversity, which in turn affected ecosystem stability; the interventions didn’t affect ecosystem stability directly, but only through changes in biodiversity as an intermediary.
For example, whether a 30% change in plant diversity … resulted from elevated N, CO2, or water or from herbivore exclusion, fire suppression, or direct manipulation of plant diversity, stability tended to decrease in parallel by 8%… This conclusion is supported by analyses showing that there was no remaining effect of anthropogenic drivers on changes in stability after biodiversity-mediated effects were taken into account [Hautier 2016: 338].
This meta-analysis finds that biodiversity across groups, especially between bacteria and fungi, contributes more to soil aggregation than species from just one group acting alone. For example, fungi specialize in binding macroaggregates, while bacteria can also bind microaggregates, and earthworms can “grind and remould ingested particles into new aggregates” [Lehmann 2017: 1]. There were no such effects from within-group biodiversity, however.
Soil biota potentially contribute to soil aggregation in a number of ways. For example, bacteria can exude biopolymers that act as binding agents for aggregates on the micrometre scale, fungal hyphae can entangle particles to hold them together (on the micrometre to millimetre scale) and geophagous animals, such as earthworms, grind and remould ingested particles into new aggregates and create biopores (on the millimetre to centimetre scale). Due to these various contributions of soil biota to soil aggregation, there is also a clear potential for complementarity among soil aggregation mechanisms, as has been shown in isolated studies [Lehmann 2017: 1].
These findings support the hypothesis that there is functional complementarity contributing to soil aggregation, and the results highlight that this functional complementarity mainly resides at the level of the HTC [Higher Taxonomic Category] . The presence of pronounced organismal interaction effects highlights the opportunity to use soil biota mixtures tailored for enhancing soil aggregation (for example, inoculation for use in restoration). This result also emphasizes the need to manage for overall high levels of soil biodiversity, especially across HTCs, in agroecosystems, which would facilitate the development of such interactions [Lehmann 2017: 4].
“Biodiversity has a major role in sustaining the productivity of Earth’s ecosystems” [Duffy 2017: 263]. This is the conclusion drawn from an analysis of 133 estimates reported in 67 field studies on the effects of species richness (number of species) on biomass production, isolating biodiversity as a variable from other factors that affect productivity (nutrient availability and climate). The results validate theoretical predictions and corroborate lab experiments showing that greater biodiversity leads to greater ecosystem production, while also refuting prevailing doubts about the significance, after accounting for other factors, of biodiversity’s effect on productivity.
Because of the long history of skepticism that species diversity affects productivity of natural ecosystems, the strength and consistency of results presented here were unanticipated. In every case we found the opposite of long-standing views expressed in the ecological literature. Ecosystems with high species richness commonly had higher biomass and productivity in observational field data from a wide range of taxa and ecosystems, including grassland plants, trees, lake phytoplankton and zooplankton, and marine fishes. Observed positive associations of biodiversity with production in nature were stronger when covariates were accounted for, stronger than biodiversity effects documented in controlled experiments, and comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers. Our results also corroborate findings of a recent synthesis of experimental data reporting that biodiversity effects are comparable in magnitude to major drivers of global change, and extend related conclusions based on observational data from forests and dryland plants to a broad range of ecosystems [Duffy 2017: 263].
Integration of this perspective [on the vital role of biodiversity] into global change policy is increasingly urgent as Earth faces widespread and potentially irreversible losses and invasions of species, which are already changing ecosystems [Duffy 2017: 263].
Observed positive associations of biodiversity with production in nature were … comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers [Duffy 2017: 263].
In a mixed forest-savanna landscape of tropical Guyana researchers found that mammal diversity is positively related to carbon concentration in the soil. The authors explain that this is due to increased feeding interaction associated with greater mammal diversity, and specify that animal abundance per se did not increase carbon content in the soil. “The lack of effect of both tree biomass and animal abundance on the response variables highlights the relevance of species richness” [Sobral 2017: 2].
“…mammal and tree richness increase the number of feeding interactions observed. The amount of organic remains (fruit and seed parts, non-fruit plant parts, faeces and animal parts) on the ground is predicted by the number of feeding interactions, and is positively related to carbon concentration in the soil. The organic remains that most affect soil carbon concentration were animal and fruit remains, which were themselves driven by carnivory and frugivory[7] interactions suggesting that both processing of fruits and direct biomass contributions by vertebrates and plants affect soil carbon concentration” [Sobral 2017: 3]
Mammal diversity influences the carbon cycle through trophic interactions in the Amazon, Sobral 2017
In a mixed forest-savanna landscape of tropical Guyana researchers found that mammal diversity is positively related to carbon concentration in the soil. The authors explain that this is due to increased feeding interaction associated with greater mammal diversity, and specify that animal abundance per se did not increase carbon content in the soil. “The lack of effect of both tree biomass and animal abundance on the response variables highlights the relevance of species richness” [Sobral 2017: 2].
“…mammal and tree richness increase the number of feeding interactions observed. The amount of organic remains (fruit and seed parts, non-fruit plant parts, faeces and animal parts) on the ground is predicted by the number of feeding interactions, and is positively related to carbon concentration in the soil. The organic remains that most affect soil carbon concentration were animal and fruit remains, which were themselves driven by carnivory and frugivory[7] interactions suggesting that both processing of fruits and direct biomass contributions by vertebrates and plants affect soil carbon concentration” [Sobral 2017: 3]
—
Biodiversity effects in the wild are common and as strong as key drivers of productivity, Duffy 2017
“Biodiversity has a major role in sustaining the productivity of Earth’s ecosystems” [Duffy 2017: 263]. This is the conclusion drawn from an analysis of 133 estimates reported in 67 field studies on the effects of species richness (number of species) on biomass production, isolating biodiversity as a variable from other factors that affect productivity (nutrient availability and climate). The results validate theoretical predictions and corroborate lab experiments showing that greater biodiversity leads to greater ecosystem production, while also refuting prevailing doubts about the significance, after accounting for other factors, of biodiversity’s effect on productivity.
Because of the long history of skepticism that species diversity affects productivity of natural ecosystems, the strength and consistency of results presented here were unanticipated. In every case we found the opposite of long-standing views expressed in the ecological literature. Ecosystems with high species richness commonly had higher biomass and productivity in observational field data from a wide range of taxa and ecosystems, including grassland plants, trees, lake phytoplankton and zooplankton, and marine fishes. Observed positive associations of biodiversity with production in nature were stronger when covariates were accounted for, stronger than biodiversity effects documented in controlled experiments, and comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers. Our results also corroborate findings of a recent synthesis of experimental data reporting that biodiversity effects are comparable in magnitude to major drivers of global change, and extend related conclusions based on observational data from forests and dryland plants to a broad range of ecosystems [Duffy 2017: 263].
Integration of this perspective [on the vital role of biodiversity] into global change policy is increasingly urgent as Earth faces widespread and potentially irreversible losses and invasions of species, which are already changing ecosystems [Duffy 2017: 263].
Observed positive associations of biodiversity with production in nature were … comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers [Duffy 2017: 263].
Soil biota contributions to soil aggregation, Lehmann 2017
This meta-analysis finds that biodiversity across groups, especially between bacteria and fungi, contributes more to soil aggregation than species from just one group acting alone. For example, fungi specialize in binding macroaggregates, while bacteria can also bind microaggregates, and earthworms can “grind and remould ingested particles into new aggregates” [Lehmann 2017: 1]. There were no such effects from within-group biodiversity, however.
Soil biota potentially contribute to soil aggregation in a number of ways. For example, bacteria can exude biopolymers that act as binding agents for aggregates on the micrometre scale, fungal hyphae can entangle particles to hold them together (on the micrometre to millimetre scale) and geophagous animals, such as earthworms, grind and remould ingested particles into new aggregates and create biopores (on the millimetre to centimetre scale). Due to these various contributions of soil biota to soil aggregation, there is also a clear potential for complementarity among soil aggregation mechanisms, as has been shown in isolated studies [Lehmann 2017: 1].
These findings support the hypothesis that there is functional complementarity contributing to soil aggregation, and the results highlight that this functional complementarity mainly resides at the level of the HTC [Higher Taxonomic Category] . The presence of pronounced organismal interaction effects highlights the opportunity to use soil biota mixtures tailored for enhancing soil aggregation (for example, inoculation for use in restoration). This result also emphasizes the need to manage for overall high levels of soil biodiversity, especially across HTCs, in agroecosystems, which would facilitate the development of such interactions [Lehmann 2017: 4].
Anthropogenic environmental changes affect ecosystem stability via biodiversity, Hautier 2015
This study illustrates the importance of biodiversity for maintaining ecosystem stability. It tests the hypothesis that “other drivers of global environmental change may have biodiversity-mediated effects on ecosystem functioning – that changes in biodiversity resulting from anthropogenic drivers may be an intermediate cause of subsequent changes in ecosystem functioning” [Hautier 2015: 337]. Researchers found that “changes in plant diversity in response to anthropogenic drivers, including N, CO2, fire, herbivory[8], and water, were positively associated with changes in temporal stability of productivity,” and that “this positive association was independent of the nature of the driver” [Hautier 2015: 338]. In other words, the experimental interventions (N, CO2, fire, etc.) affected biodiversity, which in turn affected ecosystem stability; the interventions didn’t affect ecosystem stability directly, but only through changes in biodiversity as an intermediary.
For example, whether a 30% change in plant diversity … resulted from elevated N, CO2, or water or from herbivore exclusion, fire suppression, or direct manipulation of plant diversity, stability tended to decrease in parallel by 8%… This conclusion is supported by analyses showing that there was no remaining effect of anthropogenic drivers on changes in stability after biodiversity-mediated effects were taken into account [Hautier 2016: 338].
Biodiversity for multifunctional grasslands: equal productivity in high-diversity low-input and low-diversity high-input systems, Weigelt 2009
This English grasslands study, comparing alternative strategies for increasing productivity, showed that “increasing plant species richness levels were more effective than the imposed levels of increasing management intensity” [Weigelt 2009: 1701]. The management intensification strategy included synthetic fertilization and mowing, while the biodiversity strategy increased species richness from 1 to 16 species. The authors conclude that:
For permanent grasslands, which cover one third of the utilised agricultural area in Europe (Smit et al., 2008), highly diverse communities composed of complementary species and N2-fixing legumes could provide an excellent agro-economic and ecological option for sustainable and highly productive grassland use [Weigelt 2009: 1704].
This study illustrates how ecosystem restoration enhances biodiversity and productivity. A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years, and without any human input after the original orange waste treatment of that site. The thick layer of orange peels suppressed existing non-native pasture grasses and added macro- and micronutrients to the soil, ultimately allowing for the natural (unmanaged) repopulating of the treated area from adjacent forest seedstock.
Our results provide nuance and detail to what was overwhelmingly obvious during informal surveys in 1999 and 2003: depositing orange waste on this degraded and abandoned pastureland greatly accelerated the return of tropical forest, as measured by lasting increases in soil nutrient availability, tree biomass, tree species richness, and canopy closure. The clear implication is that deposition of agricultural waste could serve as a tool for effective, low-cost tropical forest restoration, with a particularly important potential role at low-fertility sites [Treuer 2017: 6].
A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years [Treuer 2017].
Mammal diversity influences the carbon cycle through trophic interactions in the Amazon, Sobral 2017
In a mixed forest-savanna landscape of tropical Guyana researchers found that mammal diversity is positively related to carbon concentration in the soil. The authors explain that this is due to increased feeding interaction associated with greater mammal diversity, and specify that animal abundance per se did not increase carbon content in the soil. “The lack of effect of both tree biomass and animal abundance on the response variables highlights the relevance of species richness” [Sobral 2017: 2].
“…mammal and tree richness increase the number of feeding interactions observed. The amount of organic remains (fruit and seed parts, non-fruit plant parts, faeces and animal parts) on the ground is predicted by the number of feeding interactions, and is positively related to carbon concentration in the soil. The organic remains that most affect soil carbon concentration were animal and fruit remains, which were themselves driven by carnivory and frugivory[7] interactions suggesting that both processing of fruits and direct biomass contributions by vertebrates and plants affect soil carbon concentration” [Sobral 2017: 3]
—
Biodiversity effects in the wild are common and as strong as key drivers of productivity, Duffy 2017
“Biodiversity has a major role in sustaining the productivity of Earth’s ecosystems” [Duffy 2017: 263]. This is the conclusion drawn from an analysis of 133 estimates reported in 67 field studies on the effects of species richness (number of species) on biomass production, isolating biodiversity as a variable from other factors that affect productivity (nutrient availability and climate). The results validate theoretical predictions and corroborate lab experiments showing that greater biodiversity leads to greater ecosystem production, while also refuting prevailing doubts about the significance, after accounting for other factors, of biodiversity’s effect on productivity.
Because of the long history of skepticism that species diversity affects productivity of natural ecosystems, the strength and consistency of results presented here were unanticipated. In every case we found the opposite of long-standing views expressed in the ecological literature. Ecosystems with high species richness commonly had higher biomass and productivity in observational field data from a wide range of taxa and ecosystems, including grassland plants, trees, lake phytoplankton and zooplankton, and marine fishes. Observed positive associations of biodiversity with production in nature were stronger when covariates were accounted for, stronger than biodiversity effects documented in controlled experiments, and comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers. Our results also corroborate findings of a recent synthesis of experimental data reporting that biodiversity effects are comparable in magnitude to major drivers of global change, and extend related conclusions based on observational data from forests and dryland plants to a broad range of ecosystems [Duffy 2017: 263].
Integration of this perspective [on the vital role of biodiversity] into global change policy is increasingly urgent as Earth faces widespread and potentially irreversible losses and invasions of species, which are already changing ecosystems [Duffy 2017: 263].
Observed positive associations of biodiversity with production in nature were … comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers [Duffy 2017: 263].
Soil biota contributions to soil aggregation, Lehmann 2017
This meta-analysis finds that biodiversity across groups, especially between bacteria and fungi, contributes more to soil aggregation than species from just one group acting alone. For example, fungi specialize in binding macroaggregates, while bacteria can also bind microaggregates, and earthworms can “grind and remould ingested particles into new aggregates” [Lehmann 2017: 1]. There were no such effects from within-group biodiversity, however.
Soil biota potentially contribute to soil aggregation in a number of ways. For example, bacteria can exude biopolymers that act as binding agents for aggregates on the micrometre scale, fungal hyphae can entangle particles to hold them together (on the micrometre to millimetre scale) and geophagous animals, such as earthworms, grind and remould ingested particles into new aggregates and create biopores (on the millimetre to centimetre scale). Due to these various contributions of soil biota to soil aggregation, there is also a clear potential for complementarity among soil aggregation mechanisms, as has been shown in isolated studies [Lehmann 2017: 1].
These findings support the hypothesis that there is functional complementarity contributing to soil aggregation, and the results highlight that this functional complementarity mainly resides at the level of the HTC [Higher Taxonomic Category] . The presence of pronounced organismal interaction effects highlights the opportunity to use soil biota mixtures tailored for enhancing soil aggregation (for example, inoculation for use in restoration). This result also emphasizes the need to manage for overall high levels of soil biodiversity, especially across HTCs, in agroecosystems, which would facilitate the development of such interactions [Lehmann 2017: 4].
Anthropogenic environmental changes affect ecosystem stability via biodiversity, Hautier 2015
This study illustrates the importance of biodiversity for maintaining ecosystem stability. It tests the hypothesis that “other drivers of global environmental change may have biodiversity-mediated effects on ecosystem functioning – that changes in biodiversity resulting from anthropogenic drivers may be an intermediate cause of subsequent changes in ecosystem functioning” [Hautier 2015: 337]. Researchers found that “changes in plant diversity in response to anthropogenic drivers, including N, CO2, fire, herbivory[8], and water, were positively associated with changes in temporal stability of productivity,” and that “this positive association was independent of the nature of the driver” [Hautier 2015: 338]. In other words, the experimental interventions (N, CO2, fire, etc.) affected biodiversity, which in turn affected ecosystem stability; the interventions didn’t affect ecosystem stability directly, but only through changes in biodiversity as an intermediary.
For example, whether a 30% change in plant diversity … resulted from elevated N, CO2, or water or from herbivore exclusion, fire suppression, or direct manipulation of plant diversity, stability tended to decrease in parallel by 8%… This conclusion is supported by analyses showing that there was no remaining effect of anthropogenic drivers on changes in stability after biodiversity-mediated effects were taken into account [Hautier 2016: 338].
Biodiversity for multifunctional grasslands: equal productivity in high-diversity low-input and low-diversity high-input systems, Weigelt 2009
This English grasslands study, comparing alternative strategies for increasing productivity, showed that “increasing plant species richness levels were more effective than the imposed levels of increasing management intensity” [Weigelt 2009: 1701]. The management intensification strategy included synthetic fertilization and mowing, while the biodiversity strategy increased species richness from 1 to 16 species. The authors conclude that:
For permanent grasslands, which cover one third of the utilised agricultural area in Europe (Smit et al., 2008), highly diverse communities composed of complementary species and N2-fixing legumes could provide an excellent agro-economic and ecological option for sustainable and highly productive grassland use [Weigelt 2009: 1704].
This study illustrates how ecosystem restoration enhances biodiversity and productivity. A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years, and without any human input after the original orange waste treatment of that site. The thick layer of orange peels suppressed existing non-native pasture grasses and added macro- and micronutrients to the soil, ultimately allowing for the natural (unmanaged) repopulating of the treated area from adjacent forest seedstock.
Our results provide nuance and detail to what was overwhelmingly obvious during informal surveys in 1999 and 2003: depositing orange waste on this degraded and abandoned pastureland greatly accelerated the return of tropical forest, as measured by lasting increases in soil nutrient availability, tree biomass, tree species richness, and canopy closure. The clear implication is that deposition of agricultural waste could serve as a tool for effective, low-cost tropical forest restoration, with a particularly important potential role at low-fertility sites [Treuer 2017: 6].
A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years [Treuer 2017].
Remarkable roles of unremarked creatures
The articles below offer a sampling of the myriad ecosystem roles played by species we may not think much about. For example, fungi, an exemplar ecosystem cooperator, buries carbon in the soil, sources otherwise unavailable nutrients like phosphorus for plant growth, and facilitates bacterial evolution. Great whales transport nutrients through the ocean for other species to consume. Dung beetles reduce methane emissions from manure, while also fertilizing grasses. Termites and ants promote vegetation growth in arid climates by creating tunnels that catch and hold rainwater, and by making nutrients available to plants.
Fossil evidence shows that early land plants hosted fungal symbionts, which are likely to have facilitated phosphorus acquisition by plants and thus increased net primary production, perpetuating the transition to a cooler, oxygen-rich environment suitable for animal life. Mills’ study tests this hypothesis by integrating plant-fungal phosphorus acquisition into a biogeochemical model of the Paleozoic climate transition. The study finds “significant Earth system sensitivity to phosphorus uptake from mycorrhizal fungi” [Mills 2017: 7], and that “efficient phosphorus uptake at superambient CO2 results in enhanced carbon sequestration, which contributes to a reduction in CO2 and drives a rise in O2” [Mills 2017: 6].
Understanding drivers of an historic climate cooling is obviously relevant today given current atmospheric CO2 accumulation. This study points to the importance of plant-fungal symbioses and phosphorus cycling, and thus to the importance of building and protecting soil health to allow such symbioses to flourish.
Mycelia as a focal point for horizontal gene transfer among soil bacteria, Berthold 2016
Fungus is a key component of healthy soil. It is known to “translocate compounds from nutrient-rich to nutrient-poor regions… facilitate the access of bacteria to suitable microhabitats for growth, enable efficient contaminant biodegradation, and increase the functional stability in systems exposed to osmotic stress” [Berthold 2016: 5]. This study shows that, in addition, mycelia facilitate bacterial evolution, thereby bolstering bacterial diversity and adaptability.
Abstract: Horizontal gene transfer (HGT) is a main mechanism of bacterial evolution endowing bacteria with new genetic traits. The transfer of mobile genetic elements such as plasmids (conjugation) requires the close proximity of cells. HGT between genetically distinct bacteria largely depends on cell movement in water films, which are typically discontinuous in natural systems like soil. Using laboratory microcosms, a bacterial reporter system and flow cytometry, we here investigated if and to which degree mycelial networks facilitate contact of and HGT between spatially separated bacteria. Our study shows that the network structures of mycelia promote bacterial HGT by providing continuous liquid films in which bacterial migration and contacts are favoured. This finding was confirmed by individual-based simulations, revealing that the tendency of migrating bacteria to concentrate in the liquid film around hyphae is a key factor for improved HGT along mycelial networks. Given their ubiquity, we propose that hyphae can act as a focal point for HGT and genetic adaptation in soil.
The rhizosphere - roots, soil and everything in between, McNear 2013
A variety of intimate, symbiotic relationships exist between the roots of plants and the microorganisms in the soil. For instance, mycorrhizal fungi colonize the surface of plant roots, effectively extending them further through the soil to collect nutrients otherwise out of reach. These mycorrhizal branching structures, known as hyphae, emanating from plant roots also improve soil aggregation and hence improve water infiltration and aeration. In return, Mycorrhiza can demand up to 20-40% of photosynthetically derived carbon from their plant hosts. In the world of rhizospheric bacteria, Rhizobia[9] are well known for their key role in fixing atmospheric nitrogen for plant uptake. Yet there are, additionally, more than two dozen known genera of rhizobacteria that help plants grow, either directly by releasing growth stimulants (phytohormones) and enhancing mineral uptake, or indirectly by fighting off plant pathogens.
Fungal to bacterial ratios in soils investigated for enhanced C-sequestration, Bailey 2002
Testing paired sites in four ecosystem types, this study finds that higher fungal activity in soil is associated with higher soil carbon content, and that disturbing the soil reduces fungal activity. The paper’s introduction explains why fungi have been found to store more carbon than do bacteria – for example, fungi can store up to 26 times more carbon from leaf litter than bacteria. This is because the chemical composition of fungal biomass is more complex and more resistant to degradation; also, fungi have higher carbon assimilation efficiencies than do bacteria, and thus store more of the carbon they metabolize.
Whales as marine ecosystem engineers, Roman 2014
Baleen and sperm whales, known collectively as the great whales, include the largest animals in the history of life on Earth. With high metabolic demands and large populations, whales probably had a strong influence on marine ecosystems before the advent of industrial whaling: as consumers of fish and invertebrates; as prey to other large-bodied predators; as reservoirs and vertical and horizontal vectors for nutrients; and as detrital sources of energy and habitat in the deep sea. The decline in great whale numbers, estimated to be at least 66% and perhaps as high as 90%, has likely altered the structure and function of the oceans, but recovery is possible and in many cases is already underway. Future changes in the structure and function of the world’s oceans can be expected with the restoration of great whale populations.
The role of dung beetles in reducing greenhouse gas emissions from cattle farming, Slade 2015
“Dung beetles (Scarabaeidae: Scarabaeinae, Aphodiinae, Geotrupidae) are some of the most important invertebrate contributors to dung decomposition in both temperate and tropical agricultural grasslands. As such, they may help mitigate GHG [Greenhouse Gas] emissions and aid carbon sequestration through removing dung deposited on the pastures, increasing grass growth and fertilization” [Slade 2015: 1]. This Finland study analyzed the percent of GHGs removed by dung beetles at three levels: dung pat, pasture, and dairy/beef production life-cycle, finding reduced GHG emissions of 7%, 12%, and 0.05 to 0.13%, respectively. Dung beetles reduce methane emissions by aerating the dung pats, thereby preventing methane-producing anaerobic decomposition of the dung.
The reason dung beetles have a minimal effect in the full life-cycle analysis for Finland cattle is that the animals spend only a short portion of the year grazing in pasture, and thus emissions from dung on pasture is “dwarfed in comparison to other emissions of milk and meat production, such as methane emissions from enteric fermentation, nitrous oxide emissions from soils, and carbon dioxide emissions from energy use” [Slade 2015: 5]. However, “in regions where outdoor livestock grazing is more commonly used, the emissions from manure left on pasture will have a larger contribution to total agricultural emissions, with estimated fractions ranging from 11% in Asia up to 35% in Africa. Such patterns are combined with likely differences in dung beetle efficiency: In tropical regions, dung beetles can remove the majority of a fresh dung pat within the first few days after deposition – whereas in temperate conditions, a substantial fraction will remain throughout the grazing season” [Slade 2015: 5].
The authors recommend further research in tropical regions, predicting: “that effects at all levels from dung pats through pastures to the whole lifecycle of milk or beef production may be strongly accentuated at low latitudes” [Slade 2015: 5].
Termite mounds can increase the robustness of dryland ecosystems to climatic change, Bonachela 2015
Termites are particularly important in savannas of Africa, Australasia, and South America, and their nest structures (“mounds”) shape many environmental properties; analogous structures built by ants and burrowing mammals are similarly influential worldwide. Mound soils differ from surrounding “matrix” soils in physical and chemical composition, which enhances vegetation growth, creating “islands of fertility.” Moreover, mounds are frequently spatially over-dispersed owing to competition among neighboring colonies, which creates spotted vegetation patterns [Bonachela 2015: 652].
This study seeks to characterize landscape patterns created by termites in order to distinguish between that and other causes of spotted vegetation patterns that have been assumed to indicate imminent ecological collapse. “Rather, mound-field landscapes are more robust to aridity, suggesting that termites may help stabilize ecosystems under global change” [Bonachela 2015: 651].
Ants and termites increase crop yield in a dry climate, Evans 2011
Testing the effects of ants and termites on crop yield in an arid part of Australia, this study showed “that ants and termites increase wheat yield by 36% from increased soil water infiltration due to their tunnels and improved soil nitrogen” [Evans 2011: 1]. The authors conclude: “Our results suggest that ants and termites have similar functional roles to earthworms, and that they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates” [Evans 2011: 1].
Ants and termites have similar functional roles to earthworms, and . . . they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates [Evans 2011: 1].
Biodiversity refers to the outcome of 3.8 billion years of evolution since single-cellular life appeared on Earth. It is a concept embodied by an endless variety of life forms and strategies undertaken within the kingdoms of life. Biodiversity allows for a dynamic web of interactions, whereby countless organisms reliably supply one another with sufficient nutrients and shelter for survival. In technical terms, biodiversity is a measure of the total number of species in an area (species richness) weighted by the distribution of individuals across species (species evenness) [Barbour 1987: 162], and is commonly used simply to describe a particular ecological community.
Yet, because biodiversity is increasingly threatened, its relevance to the normal functioning of Earth’s systems is coming into sharper focus. Biodiversity is a primary source of the stability and productivity in ecosystems [Hautier 2015, Duffy 2017], and enhances soil carbon sequestration [Sobral 2017, Lehmann 2017, Udawatta 2011]. In the words of researcher Johan Rockström of the Stockholm Resilience Centre, “the composition of trees, plants, microbes in soils, phytoplankton in oceans, top predators in ecosystems…together constitute a fundamental core contributor to regulating the state of the planet.” Rockström continues: “Without biodiversity, no ecosystems. No ecosystems, no biomes. No biomes, no living regulator of all the cycles of carbon, nitrogen, oxygen, carbon dioxide and water” [Hance 2018].
An important component of biodiversity is “functional diversity,” which refers to “the range and value of those species and organismal traits that influence ecosystem functioning” [Tilman 2001: 109]. In other words, specific traits of various species, like the ability to photosynthesize, to decompose dead organic matter, or to control herbivore populations, affect the way an ecosystem operates.
The reintroduction of wolves in Yellowstone Park illustrates this: when wolves were reintroduced they reduced overgrazing by elks in the valleys, which allowed aspen, willow and cottonwood trees to grow back into forest, in turn stabilizing the banks of the river. The return of trees brought back birds and beavers. The beaver dams are believed to have brought back otters, muskrats, fish, ducks, reptiles and amphibians. The wolves also controlled coyote populations, allowing mice and rabbit populations to rebound and attract other predators, including ravens and bald eagles. This magnificent ecosystem-wide ripple effect from reintroducing the top predator is called a “trophic cascade” [Ripple 2011], and it was poignantly illustrated in a short video narrated by George Monbiot [Monbiot 2014].
In short, particular species and groups of species, including ones we may not like, know or care about, play vital roles. In the collection of articles below, we highlight the ecological importance of biodiversity in general, as well as varieties of particular species with remarkable roles to play in the perpetuation of a biologically productive and habitable world. Notably, fungi play a starring role.
Compilation of biodiversity articles
Biodiversity
Mammal diversity influences the carbon cycle through trophic interactions in the Amazon, Sobral 2017
In a mixed forest-savanna landscape of tropical Guyana researchers found that mammal diversity is positively related to carbon concentration in the soil. The authors explain that this is due to increased feeding interaction associated with greater mammal diversity, and specify that animal abundance per se did not increase carbon content in the soil. “The lack of effect of both tree biomass and animal abundance on the response variables highlights the relevance of species richness” [Sobral 2017: 2].
“…mammal and tree richness increase the number of feeding interactions observed. The amount of organic remains (fruit and seed parts, non-fruit plant parts, faeces and animal parts) on the ground is predicted by the number of feeding interactions, and is positively related to carbon concentration in the soil. The organic remains that most affect soil carbon concentration were animal and fruit remains, which were themselves driven by carnivory and frugivory[7] interactions suggesting that both processing of fruits and direct biomass contributions by vertebrates and plants affect soil carbon concentration” [Sobral 2017: 3]
—
Biodiversity effects in the wild are common and as strong as key drivers of productivity, Duffy 2017
“Biodiversity has a major role in sustaining the productivity of Earth’s ecosystems” [Duffy 2017: 263]. This is the conclusion drawn from an analysis of 133 estimates reported in 67 field studies on the effects of species richness (number of species) on biomass production, isolating biodiversity as a variable from other factors that affect productivity (nutrient availability and climate). The results validate theoretical predictions and corroborate lab experiments showing that greater biodiversity leads to greater ecosystem production, while also refuting prevailing doubts about the significance, after accounting for other factors, of biodiversity’s effect on productivity.
Because of the long history of skepticism that species diversity affects productivity of natural ecosystems, the strength and consistency of results presented here were unanticipated. In every case we found the opposite of long-standing views expressed in the ecological literature. Ecosystems with high species richness commonly had higher biomass and productivity in observational field data from a wide range of taxa and ecosystems, including grassland plants, trees, lake phytoplankton and zooplankton, and marine fishes. Observed positive associations of biodiversity with production in nature were stronger when covariates were accounted for, stronger than biodiversity effects documented in controlled experiments, and comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers. Our results also corroborate findings of a recent synthesis of experimental data reporting that biodiversity effects are comparable in magnitude to major drivers of global change, and extend related conclusions based on observational data from forests and dryland plants to a broad range of ecosystems [Duffy 2017: 263].
Integration of this perspective [on the vital role of biodiversity] into global change policy is increasingly urgent as Earth faces widespread and potentially irreversible losses and invasions of species, which are already changing ecosystems [Duffy 2017: 263].
Observed positive associations of biodiversity with production in nature were … comparable to or stronger than associations with climate and nutrient availability, which are arguably two of the strongest abiotic drivers of ecosystem structure and functioning, as well as major global change drivers [Duffy 2017: 263].
Soil biota contributions to soil aggregation, Lehmann 2017
This meta-analysis finds that biodiversity across groups, especially between bacteria and fungi, contributes more to soil aggregation than species from just one group acting alone. For example, fungi specialize in binding macroaggregates, while bacteria can also bind microaggregates, and earthworms can “grind and remould ingested particles into new aggregates” [Lehmann 2017: 1]. There were no such effects from within-group biodiversity, however.
Soil biota potentially contribute to soil aggregation in a number of ways. For example, bacteria can exude biopolymers that act as binding agents for aggregates on the micrometre scale, fungal hyphae can entangle particles to hold them together (on the micrometre to millimetre scale) and geophagous animals, such as earthworms, grind and remould ingested particles into new aggregates and create biopores (on the millimetre to centimetre scale). Due to these various contributions of soil biota to soil aggregation, there is also a clear potential for complementarity among soil aggregation mechanisms, as has been shown in isolated studies [Lehmann 2017: 1].
These findings support the hypothesis that there is functional complementarity contributing to soil aggregation, and the results highlight that this functional complementarity mainly resides at the level of the HTC [Higher Taxonomic Category] . The presence of pronounced organismal interaction effects highlights the opportunity to use soil biota mixtures tailored for enhancing soil aggregation (for example, inoculation for use in restoration). This result also emphasizes the need to manage for overall high levels of soil biodiversity, especially across HTCs, in agroecosystems, which would facilitate the development of such interactions [Lehmann 2017: 4].
Anthropogenic environmental changes affect ecosystem stability via biodiversity, Hautier 2015
This study illustrates the importance of biodiversity for maintaining ecosystem stability. It tests the hypothesis that “other drivers of global environmental change may have biodiversity-mediated effects on ecosystem functioning – that changes in biodiversity resulting from anthropogenic drivers may be an intermediate cause of subsequent changes in ecosystem functioning” [Hautier 2015: 337]. Researchers found that “changes in plant diversity in response to anthropogenic drivers, including N, CO2, fire, herbivory[8], and water, were positively associated with changes in temporal stability of productivity,” and that “this positive association was independent of the nature of the driver” [Hautier 2015: 338]. In other words, the experimental interventions (N, CO2, fire, etc.) affected biodiversity, which in turn affected ecosystem stability; the interventions didn’t affect ecosystem stability directly, but only through changes in biodiversity as an intermediary.
For example, whether a 30% change in plant diversity … resulted from elevated N, CO2, or water or from herbivore exclusion, fire suppression, or direct manipulation of plant diversity, stability tended to decrease in parallel by 8%… This conclusion is supported by analyses showing that there was no remaining effect of anthropogenic drivers on changes in stability after biodiversity-mediated effects were taken into account [Hautier 2016: 338].
Biodiversity for multifunctional grasslands: equal productivity in high-diversity low-input and low-diversity high-input systems, Weigelt 2009
This English grasslands study, comparing alternative strategies for increasing productivity, showed that “increasing plant species richness levels were more effective than the imposed levels of increasing management intensity” [Weigelt 2009: 1701]. The management intensification strategy included synthetic fertilization and mowing, while the biodiversity strategy increased species richness from 1 to 16 species. The authors conclude that:
For permanent grasslands, which cover one third of the utilised agricultural area in Europe (Smit et al., 2008), highly diverse communities composed of complementary species and N2-fixing legumes could provide an excellent agro-economic and ecological option for sustainable and highly productive grassland use [Weigelt 2009: 1704].
This study illustrates how ecosystem restoration enhances biodiversity and productivity. A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years, and without any human input after the original orange waste treatment of that site. The thick layer of orange peels suppressed existing non-native pasture grasses and added macro- and micronutrients to the soil, ultimately allowing for the natural (unmanaged) repopulating of the treated area from adjacent forest seedstock.
Our results provide nuance and detail to what was overwhelmingly obvious during informal surveys in 1999 and 2003: depositing orange waste on this degraded and abandoned pastureland greatly accelerated the return of tropical forest, as measured by lasting increases in soil nutrient availability, tree biomass, tree species richness, and canopy closure. The clear implication is that deposition of agricultural waste could serve as a tool for effective, low-cost tropical forest restoration, with a particularly important potential role at low-fertility sites [Treuer 2017: 6].
A one-time application in 1998 of 1,000 truckloads of agricultural waste (orange peels) to 3 ha of degraded pasture accelerated tropical forest regeneration in this Costa Rica study. The treatment led to a tripling in species richness (24 tree species from 20 families, compared to 8 tree species from 7 families in the control plot), and 176% increase in aboveground biomass after 16 years [Treuer 2017].
Remarkable roles of unremarked creatures
The articles below offer a sampling of the myriad ecosystem roles played by species we may not think much about. For example, fungi, an exemplar ecosystem cooperator, buries carbon in the soil, sources otherwise unavailable nutrients like phosphorus for plant growth, and facilitates bacterial evolution. Great whales transport nutrients through the ocean for other species to consume. Dung beetles reduce methane emissions from manure, while also fertilizing grasses. Termites and ants promote vegetation growth in arid climates by creating tunnels that catch and hold rainwater, and by making nutrients available to plants.
Fossil evidence shows that early land plants hosted fungal symbionts, which are likely to have facilitated phosphorus acquisition by plants and thus increased net primary production, perpetuating the transition to a cooler, oxygen-rich environment suitable for animal life. Mills’ study tests this hypothesis by integrating plant-fungal phosphorus acquisition into a biogeochemical model of the Paleozoic climate transition. The study finds “significant Earth system sensitivity to phosphorus uptake from mycorrhizal fungi” [Mills 2017: 7], and that “efficient phosphorus uptake at superambient CO2 results in enhanced carbon sequestration, which contributes to a reduction in CO2 and drives a rise in O2” [Mills 2017: 6].
Understanding drivers of an historic climate cooling is obviously relevant today given current atmospheric CO2 accumulation. This study points to the importance of plant-fungal symbioses and phosphorus cycling, and thus to the importance of building and protecting soil health to allow such symbioses to flourish.
Mycelia as a focal point for horizontal gene transfer among soil bacteria, Berthold 2016
Fungus is a key component of healthy soil. It is known to “translocate compounds from nutrient-rich to nutrient-poor regions… facilitate the access of bacteria to suitable microhabitats for growth, enable efficient contaminant biodegradation, and increase the functional stability in systems exposed to osmotic stress” [Berthold 2016: 5]. This study shows that, in addition, mycelia facilitate bacterial evolution, thereby bolstering bacterial diversity and adaptability.
Abstract: Horizontal gene transfer (HGT) is a main mechanism of bacterial evolution endowing bacteria with new genetic traits. The transfer of mobile genetic elements such as plasmids (conjugation) requires the close proximity of cells. HGT between genetically distinct bacteria largely depends on cell movement in water films, which are typically discontinuous in natural systems like soil. Using laboratory microcosms, a bacterial reporter system and flow cytometry, we here investigated if and to which degree mycelial networks facilitate contact of and HGT between spatially separated bacteria. Our study shows that the network structures of mycelia promote bacterial HGT by providing continuous liquid films in which bacterial migration and contacts are favoured. This finding was confirmed by individual-based simulations, revealing that the tendency of migrating bacteria to concentrate in the liquid film around hyphae is a key factor for improved HGT along mycelial networks. Given their ubiquity, we propose that hyphae can act as a focal point for HGT and genetic adaptation in soil.
The rhizosphere - roots, soil and everything in between, McNear 2013
A variety of intimate, symbiotic relationships exist between the roots of plants and the microorganisms in the soil. For instance, mycorrhizal fungi colonize the surface of plant roots, effectively extending them further through the soil to collect nutrients otherwise out of reach. These mycorrhizal branching structures, known as hyphae, emanating from plant roots also improve soil aggregation and hence improve water infiltration and aeration. In return, Mycorrhiza can demand up to 20-40% of photosynthetically derived carbon from their plant hosts. In the world of rhizospheric bacteria, Rhizobia[9] are well known for their key role in fixing atmospheric nitrogen for plant uptake. Yet there are, additionally, more than two dozen known genera of rhizobacteria that help plants grow, either directly by releasing growth stimulants (phytohormones) and enhancing mineral uptake, or indirectly by fighting off plant pathogens.
Fungal to bacterial ratios in soils investigated for enhanced C-sequestration, Bailey 2002
Testing paired sites in four ecosystem types, this study finds that higher fungal activity in soil is associated with higher soil carbon content, and that disturbing the soil reduces fungal activity. The paper’s introduction explains why fungi have been found to store more carbon than do bacteria – for example, fungi can store up to 26 times more carbon from leaf litter than bacteria. This is because the chemical composition of fungal biomass is more complex and more resistant to degradation; also, fungi have higher carbon assimilation efficiencies than do bacteria, and thus store more of the carbon they metabolize.
Whales as marine ecosystem engineers, Roman 2014
Baleen and sperm whales, known collectively as the great whales, include the largest animals in the history of life on Earth. With high metabolic demands and large populations, whales probably had a strong influence on marine ecosystems before the advent of industrial whaling: as consumers of fish and invertebrates; as prey to other large-bodied predators; as reservoirs and vertical and horizontal vectors for nutrients; and as detrital sources of energy and habitat in the deep sea. The decline in great whale numbers, estimated to be at least 66% and perhaps as high as 90%, has likely altered the structure and function of the oceans, but recovery is possible and in many cases is already underway. Future changes in the structure and function of the world’s oceans can be expected with the restoration of great whale populations.
The role of dung beetles in reducing greenhouse gas emissions from cattle farming, Slade 2015
“Dung beetles (Scarabaeidae: Scarabaeinae, Aphodiinae, Geotrupidae) are some of the most important invertebrate contributors to dung decomposition in both temperate and tropical agricultural grasslands. As such, they may help mitigate GHG [Greenhouse Gas] emissions and aid carbon sequestration through removing dung deposited on the pastures, increasing grass growth and fertilization” [Slade 2015: 1]. This Finland study analyzed the percent of GHGs removed by dung beetles at three levels: dung pat, pasture, and dairy/beef production life-cycle, finding reduced GHG emissions of 7%, 12%, and 0.05 to 0.13%, respectively. Dung beetles reduce methane emissions by aerating the dung pats, thereby preventing methane-producing anaerobic decomposition of the dung.
The reason dung beetles have a minimal effect in the full life-cycle analysis for Finland cattle is that the animals spend only a short portion of the year grazing in pasture, and thus emissions from dung on pasture is “dwarfed in comparison to other emissions of milk and meat production, such as methane emissions from enteric fermentation, nitrous oxide emissions from soils, and carbon dioxide emissions from energy use” [Slade 2015: 5]. However, “in regions where outdoor livestock grazing is more commonly used, the emissions from manure left on pasture will have a larger contribution to total agricultural emissions, with estimated fractions ranging from 11% in Asia up to 35% in Africa. Such patterns are combined with likely differences in dung beetle efficiency: In tropical regions, dung beetles can remove the majority of a fresh dung pat within the first few days after deposition – whereas in temperate conditions, a substantial fraction will remain throughout the grazing season” [Slade 2015: 5].
The authors recommend further research in tropical regions, predicting: “that effects at all levels from dung pats through pastures to the whole lifecycle of milk or beef production may be strongly accentuated at low latitudes” [Slade 2015: 5].
Termite mounds can increase the robustness of dryland ecosystems to climatic change, Bonachela 2015
Termites are particularly important in savannas of Africa, Australasia, and South America, and their nest structures (“mounds”) shape many environmental properties; analogous structures built by ants and burrowing mammals are similarly influential worldwide. Mound soils differ from surrounding “matrix” soils in physical and chemical composition, which enhances vegetation growth, creating “islands of fertility.” Moreover, mounds are frequently spatially over-dispersed owing to competition among neighboring colonies, which creates spotted vegetation patterns [Bonachela 2015: 652].
This study seeks to characterize landscape patterns created by termites in order to distinguish between that and other causes of spotted vegetation patterns that have been assumed to indicate imminent ecological collapse. “Rather, mound-field landscapes are more robust to aridity, suggesting that termites may help stabilize ecosystems under global change” [Bonachela 2015: 651].
Ants and termites increase crop yield in a dry climate, Evans 2011
Testing the effects of ants and termites on crop yield in an arid part of Australia, this study showed “that ants and termites increase wheat yield by 36% from increased soil water infiltration due to their tunnels and improved soil nitrogen” [Evans 2011: 1]. The authors conclude: “Our results suggest that ants and termites have similar functional roles to earthworms, and that they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates” [Evans 2011: 1].
Ants and termites have similar functional roles to earthworms, and . . . they may provide valuable ecosystem services in dryland agriculture, which may become increasingly important for agricultural sustainability in arid climates [Evans 2011: 1].
Geotherapy: Innovative Methods of Soil Fertility Restoration, Carbon Sequestration, and Reversing CO2 Increase. Edited by Thomas J. Goreau, Ronal W. Larsen and Joanna Campe [Goreau 2015]
The term “geotherapy” was coined by Richard Grantham, an evolutionary biologist and geneticist who, in his later years, turned his attention to the deteriorating state of Earth in the current era, the Anthropocene. He regarded the planet as ill, as a patient, in need of treatment.
The first geotherapy conference was held in 1991 in France, preceding the first international climate conference in Rio in 1992. It was underfunded and the papers presented could not be published. In the meanwhile the international community focused virtually all of its attention on climate as a greenhouse gas problem, and the powerful biological drivers of not only climate but most processes on Earth receded into the background.
This book is an invaluable contribution to redressing serious oversights on the part of mainstream climate science. For example, anthropogenic climate change began long before the industrial revolution, as leading soil scientist Rattan Lal states in his Preface to Geotherapy:
One of the consequences of the drastic anthropogenic perturbation of the biosphere is the depletion of the ecosystem and soil carbon pools. Rather than commencing with the onset of the industrial revolution around 1750, anthropocene began with the beginning of settled agriculture 10 to 12 millennia ago. Over this period, more carbon may have been emitted into the atmosphere from deforestation and land use conversion than from fossil fuel combustion until the end of the twentieth century. Thus, recarbonization of the biosphere in general and that of the soil carbon pool in particular is important to the maintenance and enhancements of ecosystem functions and services. [Goreau 2015: xv-xvi]
Geotherapy is an anthology of thirty-four scientific articles that sketch a roadmap to planetary health. Topics covered range from biochar to rock powders to waste nutrient recycling to remediation with plants to carbon farming (farming with soil health a primary concern), and more. The articles are well-illustrated, well-referenced, and accessible to a layperson generally familiar with scientific writing. It’s a guidebook with many shovel-ready approaches as well as theoretical explanations. The sense of both urgency and hope in bringing back living systems to the anthropocene landscape is palpable:
If soils are not restored, crops will fail even if rains do not; hunger will perpetuate even with emphasis on biotechnology and genetically modified crops; civil strife and political instability will plague the developing world even with sermons on human rights and democratic ideals; and humanity will suffer even with great scientific strides. Political stability and global peace are threatened because of soil degradation, food insecurity, and desperateness. The time to act is now. [Rattan Lal in Goreau 2015: xvi]
In this second issue of the Compendium of Scientific and Practical Findings Supporting Eco-Restoration to Address Global Warming by Biodiversity for a Livable Climate (“Bio4Climate”), we focus on the pivotal roles of biodiversity and regenerative agriculture in stabilizing ecosystems and the climate. We review a selection of a large and growing trove of research demonstrating the relevance of biodiversity and regenerative agriculture for an effective response to the climate crisis.
We also include important and valuable information that is validated through land-management and other practice that has not yet appeared in peer-reviewed literature, which tends to be conservative and biased towards mainstream assumptions.[3] We consider these practices “positive variants” which hold great promise in addressing current rapidly growing environmental crises, including global warming.
Biodiversity and regenerative agriculture represent fertile ground for finding solutions, and should therefore be in the forefront of public discourse. Yet despite mounting and compelling evidence of a link, the relationship between biodiversity and the climate is not necessarily intuitive, nor is the connection often made in mainstream news or in political negotiations. Even in the face of biodiversity collapse, we broadly fail to prioritize biodiversity, let alone consider it as a key factor in the search for solutions to the climate crisis.
With respect to agriculture, an interest in agriculture-based climate solutions is evident among scientists, and this interest appears to be reaching the political sphere as well. Yet there is an apparent reluctance to look outside the context of the current, export-oriented, input-intensive system of agriculture. Instead, there is an assumption that we can improve the conservation potential of agriculture while remaining within the current agricultural paradigm, under the pretext that high external inputsare needed to “feed the world.”
Indeed, agriculture is a linchpin issue for humanity. Our survival as a civilization depends on viable agriculture systems. However, input-intensive agriculture has given us false hopes about technology-aided yield potential, while at the same time diminishing the soil’s inherent ability to provide for plant health and nutrition in an era of increasingly harshclimatic conditions for crop and livestock production. Yet agricultural land, which covers some 40% of Earth’s land surface [Foley 2005], could be a source of planetary regeneration. Indeed, it appears to be ONLY through regenerative agriculture that we will be able to feed ourselves in the future, since high-inputagriculture is ultimately a far more fragile system. Industrial agriculture is more vulnerable to weather extremes, pest invasions, andhighly reliant on increasingly scarce and expensive external inputs.
The purpose of this Compendium, and of Bio4Climate’s approach overall, is to assemble and showcase solutions to global warming that are largely already known, all of them rootedin ecosystem restoration. Since studies demonstrating the power of biological processes, biodiversity, and intact ecosystems to restore balance to the climate system are dispersed across multiple disciplines that may often be unaware of one another’s work, we attempt to shed light on such relationships and consider the climate crisis in a complex systems framework.
Understanding the planet as a complex system, encompassing myriadliving and non-living subsystems, opens up our awareness to the interdependence among seemingly unrelated processes, and to the possibility of indirect and cascading effects and abrupt changes. It helps us to accept and appreciate the vast complexity of billions of simultaneous processes that cannot be fully controlled, and yet also to recognize the patterns that restore balance to the systems sustaining human life (such as how protected and revived soil accumulates carbon and water that would otherwise be in the atmosphere or ocean. See Appendix B for further discussion of a systems approach to climate change.)
At the same time, these ecosystem approaches will be successful only if they are actually undertaken and replicated systematically, the world over. Therefore, it behooves us to contemplate the urgency of the crisis before us. The positive feedbacks[4] in climate, as witnessed by the dramatic accelerations of weather crises and many environmental degradations such as extinctions (including the unprecedented disappearance of insects), plankton loss, and disrupted timing of lifecycles and species migrations, raise such alarm that even while focusing on promising solutions, we must fully acknowledge our current dire situation. (See Appendix A for further discussion of current urgencies.)
The articles featured in this Compendium reveal the power of ecosystem properties and processes, when protected from human hyper-exploitation, to restore life and health to human society and to many other organisms upon whose wellbeing we are entirely dependent. Specifically, the effect of biodiversity at various taxonomic levels on ecosystem productivity rivals that of abiotic factors [Weigelt 2009, Duffy 2017, Lehmann 2017, Sobral 2017]. Furthermore, often-overlooked groups of species play major roles in ecosystems. Notably, fungi are associated with high soil carbon content and productivity, and with phosphorus cycling [Bailey 2002, Johnson 2017, Berthold 2016, Mills 2017].
In fact, there is poor understanding of the phosphorus[5] cycle unless you include the work of the fungi. Mark McMenamin, who spoke at Bio4Climate’s Oceans Conference in 2016,[6] wonders why land has perhaps 100 times the biomass and triple the productivity compared to the oceans [McMenamin 2016]. His theory, “Hypersea” [McMenamin 1996] proposes that high productivity on land is a result of the ways biodiversity creates “upwellings,“ which bring up essential minerals and water, often from deep in rocky soils, to facilitate photosynthesis. Fungi retrieve micronutrients for plants in exchange for the energy provided by the glucose produced by green plants.
In the ocean, phosphorus is available if there are upwellings from the bottom by winds, seamounts or currents. Phosphorus, nitrogen and other minerals are rapidly consumed by algae and if they are not replenished regularly then algae growth stops. The ocean is largely an “aqueous desert” because movement of nutrients to the surface from the deeper ocean is relatively rare.
Thus, Hypersea is about upwelling of nutrients. High productivity on land is because of fungi – plant symbioses. But human chemical agriculture has interrupted this system. By adding enormous amounts of nitrogen, phosphorus, and potassium at great expense, we have temporarily increased crop yields. However, these inputs cause bacteria populations to multiply and consume the mycorrhizal fungi networks. Because of this, nutrient flow from deep in the soil slows or stops.
Yet, to the extent that humans are capable of disrupting phosphorus, nitrogen, carbon, water and other cycles that are at once driven by and sustaining of Earth’s biosphere, we are also capable of acting to repair these cycles as Trant [2016] and Treuer [2017] illustrate – ancient and modern peoples alike have improved ecosystem productivity by composting food wastes, for example.
The books Drawdown and Geotherapy similarly offer a wealth of specific, proven and practical steps for restabilizing the climate. “Humanity has the means at hand. Nothing new needs to be invented. The solutions are in place and in action. Our work is to accelerate the knowledge and growth of what is possible” [Hawken 2017]. Indeed, illuminating that picture of what is possible is the purpose of this Compendium.
Understanding the role of paradigms in scientific investigation is one of the keys to approaching the revolutionary view of climate as a problem of ecosystem dynamics as opposed to one simply of excessive greenhouse gases. The new paradigm doesn’t render the old paradigm irrelevant, but it reframes its significance and role in addressing the current climate crisis. It exposes to open examination what was heretofore an invisible phenomenon, and avails a universe of solutions to what is, from the perspective of the greenhouse gas hypothesis, an intractable and quite possibly utterly hopeless problem. Therefore, we will take a moment to review the paradigm process and apply it to our contending climate paradigms.
In 1962, Thomas Kuhn, a Harvard-trained physicist who became a historian and philosopher of science, published a controversial book, The Structure of Scientific Revolutions. Prior to Kuhn, the prevailing assumptions about the way science progressed were that knowledge was gradually accumulated by generations of investigators, with occasional quantum leaps by great scientists, but in an overall smooth and continuous albeit occasionally heroic process.
Kuhn broke new ground by re-examining and reframing the process of scientific investigation.
He brought the term “paradigm” into common usage, by which he meant a body of “universally recognized scientific achievements that for a time provide model problems and solutions to a community of practitioners” (p. viii, emphasis added). We will review Kuhn’s work briefly and apply his analysis when comparing the mainstream greenhouse gas climate paradigm and the newly evolving eco-restoration climate paradigm.
Kuhn maintained that scientific progress is episodic, characterized by long periods of “normal science,” which takes place in the context of a paradigm:
At least in the mature sciences, answers (or full substitutes for answers) to [many] questions . . . are firmly embedded in the educational initiation that prepares and licenses the student for professional practice. Because that education is both rigorous and rigid, these answers come to exert a deep hold on the scientific mind. [Kuhn 1962:5]
Normal science, the activity in which most scientists inevitably spend almost all their time, is predicated on the assumption that the scientific community knows what the world is like. Much of the success of the enterprise derives from the community’s willingness to defend that assumption, if necessary at considerable cost. Normal science, for example, often suppresses fundamental novelties because they are necessarily subversive of its basic commitments. Nevertheless, so long as those commitments retain an element of the arbitrary, the very nature of normal research ensures that novelty shall not be suppressed for very long. [Kuhn 1962:5]
Normal science is punctuated by the appearance of anomalies which cannot be explained by the paradigm’s generally accepted theories, nor tested by what the paradigm might consider reasonable hypotheses, nor resolved with current testing protocols or equipment.
When examining normal science . . . we shall want finally to describe that research as a strenuous and devoted attempt to force nature into the conceptual boxes supplied by professional education. [Kuhn 1962:5]
[W]hen [normal science repeatedly goes astray] – when, that is, the profession can no longer evade anomalies that subvert the existing tradition of scientific practice – then begin the extraordinary investigations that lead the profession at last to a new set of commitments, a new basis for the practice of science. The extraordinary episodes in which that shift of professional commitments occurs are the ones known in this essay as scientific revolutions. They are the tradition-shattering complements to the tradition-bound activity of normal science. [Kuhn 1962: 6]
and
Normal science consists in . . . an actualization achieved by extending the knowledge of those facts that the paradigm displays as particularly revealing, by increasing the extent of the match between those facts and the paradigm’s predictions, and by further articulation of the paradigm itself.
Few people who are not actually practitioners of a mature science realize how much mop-up work of this sort a paradigm leaves to be done or quite how fascinating such work can prove in the execution. And these points need to be understood. Mopping-up operations are what engage most scientists throughout their careers. They constitute what I am here calling normal science. Closely examined, whether historically or in the contemporary laboratory, that enterprise seems an attempt to force nature into the preformed and relatively inflexible box that the paradigm supplies. No part of the aim of normal science is to call forth new sorts of phenomena; indeed those that will not fit the box are often not seen at all. Nor do scientists normally aim to invent new theories, and they are often intolerant of those invented by others. Instead, normal-scientific research is directed to the articulation of those phenomena and theories that the paradigm already supplies. [Kuhn 1962:23-24, emphasis added]
and
Paradigms gain their status because they are more successful than their competitors in solving a few problems that the group of practitioners has come to recognize as acute. To be more successful is not, however, to be either completely successful with a single problem or notably successful with any large number. The success of a paradigm . . . is at the start largely a promise of success discoverable in selected and still incomplete examples. [Kuhn 1962:23, emphasis added]
Even today, over half a century after Structures was originally published, normal science seems immune to the possibilities of paradigm shifts – such thoughts often do not occur until forced, even though the process should be reasonably well known if not entirely understood or accepted. The prevailing opinion about paradigm shifts (if there is any opinion at all) appears to be, “It doesn’t apply to my paradigm.”
In general, a paradigm shift doesn’t only involve “objective” factors, it touches scientific practitioners at a deep emotional level as well, as any participant in or observer of academic dispute can testify:
Scientific fact and theory are not categorically separable, except perhaps within a
single tradition of normal-scientific practice. That is why the unexpected discovery is not simply factual in its import and why the scientist’s world is qualitatively transformed as well as quantitatively enriched by fundamental novelties of either fact or theory. [Kuhn 1962:7]
Therefore, the transition to a new paradigm is disruptive and challenging:
The transition from a paradigm in crisis to a new one from which a new tradition of normal science can emerge is far from a cumulative process, one achieved by an articulation or extension of the old paradigm. Rather it is a reconstruction of the field from new fundamentals, a reconstruction that changes some of the field’s most elementary theoretical generalizations as well as many of its paradigm methods and applications. During the transition period there will be a large but never complete overlap between the problems that can be solved by the old and by the new paradigm. But there will also be a decisive difference in the modes of solution. When the transition is complete, the profession will have changed its view of the field, its methods, and its goals [Kuhn 1962:84-85].
The case in point here is the comparison between old and new climate paradigms
Paradigm step
Old paradigm (greenhouse gases)
New paradigm (Eco-restoration)
Paradigm fundamentals
CO2 and equiv are greenhouse gas blankets and elevated levels cause global warming, primarily caused by burning fossil fuels
Destruction of billions of acres of land interferes with carbon and water cycles, along with oxidation of soils for over 10k years, puts gigatons of carbon into atmosphere
Weaknesses intrinsic to paradigm
Positive feedbacks underrepresented, overlooked, not calculated or estimated;
biology is characterized as passive victim of climate change
Complex, interdependent systems that are difficult to model and to quantify into policy
Strengths intrinsic to paradigm
Amenable to modeling; yields numeric targets that can be translated into policy
Comprehensive of all likely drivers and their theoretical interdependencies. Plausible upon examination of biogeologic history.
Primary investigators
Physical scientists almost exclusively from academia
Restoration ecologists and others from biological sciences; non-academic land managers
Tools
Emissions reductions via alternative energy and elimination of carbon emissions sources
Photosynthesis and regenerative land management
Costs
High
Low
Technology requirements
Extensive
Minimal
Locus of investigation
Centralized in academia – universities, scientific journals, formal test sites
Based first in local land management practice, then investigated by academia, landscape managers, local practitioners – farmers, ranchers, horticulturalists, permaculturists, indigenous cultures, etc.
Weight of evidence
Formal studies, isolated variables
Practical results, holistic assessment of land health, biodiversity, water and carbon cycling
Success criteria
Reduced emissions and atmospheric carbon burdens (target 350 ppm? lower?)
Increased biodiversity, improved water cycles, land resilience, cooling of local biospheres on a global scale, reduced floods and droughts, decline in atmospheric carbon burdens (target 280 ppm)
Duration of existence of paradigm
Roughly 200 years
Roughly 20 years with some roots going back considerably longer
The Perils of Peer Review
It may well behoove us all, including the scientists among us, to take a careful look at how science works today and how practices may improve. For example, while peer review can be a powerful tool, it is worth keeping in mind that we’re in a world of shifting paradigms where there are libraries full of peer-reviewed papers in scientific landfills, review processes notwithstanding. Peer review may be a useful tool, but it may also be a significant obstacle to scientific progress.
Beyond the routine aging and demise of most scientific papers, however, is the problem of a system that excludes information that conflicts with or is invisible to the dominant paradigm. That is a central issue that we face when introducing an entirely new view of climate – in the normal course of science, taking a generation or two to transition between paradigms is acceptable, even healthy; in the throes of a climate emergency, conventional peer review may be a serious obstacle to progress.
Peer reviewers get to review their peers because they are thinking along similar lines, and are likely – perhaps even required – to reject ideas outside mainstream boundaries of thought. Even the conventional authors of a recent USDA study had a difficult time getting their study published because its results were so unexpected [Ausmus 2014; Follett 2012]. And that doesn’t begin to touch any of the forces in the political and economic realms that impact peer-reviewed science, including what actually gets studied (and funded!) and what does not.
Presence or absence of peer review should therefore not serve as a standard for accepting the validity of any paper or report; only the evidence is the basis for such decisions, whether it is within “acceptable” range or far beyond it. The evidence must stand for itself, and the professionals who are readers should have the opportunity to make up their own minds. Peer review is a standard, but not a gold standard – it is one among many, and practitioners of scientific method have an obligation to evaluate the relevance of standards.
Linkov 2006. Whereas most tools of science have evolved over the past three hundred years, there is one that stubbornly shows its age: the scientific journal. The author “argue[s] that the primary reason that journals have not changed is that they are ‘faith based’: we believe in them, we dare not question them.” [Linkov 2006: 596]
Linkov suggests that the journal hasn’t transformed into a new model of publication because it has never applied the scientific method to itself.
Jefferson recently presented an outstanding review of peer review and could find only 19 studies on peer review that were scientifically sound. We could find only 14 articles examining the editorial board/editorial decision making. Thus, with over 50 million articles and 300 years of the traditional journal approaches, there has been only a handful of studies questioning or testing the journal process itself. We scientists keep using the process without question, but with no data to show that it is effective. There is thus no evidence-based approach to the science of research communications. Recent studies reveal that peer review often misses major methodological problems in articles. No wonder it has not changed or improved, as there are no data questioning the process. Hypothesis testing research and randomized trials could easily and cheaply be initiated to understand the ‘grand challenges’ of research communication, but sadly they have not.
Isn’t it strange that three features that are inherent to research communication have not been looked at scientifically? There are several possible reasons for this. The most likely is that we scientists have almost complete faith in the journal process as right and unassailable. We thus take a ‘faith based’ approach to research communications. Faith is defined as a firm belief in something for which there is no proof. Many of us might view questioning of the journal process as an attack on science itself. Clearly, the scientific journal process is not a part of the scientific method. We are taught early in our training about the importance of learning to write articles (e.g. IMRaD[23]), the power of peer review and a belief in the editorial system. We do not question the process, despite the fact that the essence of science is questioning. Questioning peer review is like questioning the Bible, Quran or Torah. One role of science is to help separate science from dogma, which we should now do with journals, and avoid a faith based approach. New approaches need to be taken – you cannot teach dogma new tricks! [Linkov 2006: 597]
It is the scientific method that is central to science, not the scientific journal. The scientific method should be central to other research communication processes, but it is not and has not been used to continuously improve how we communicate research. Because of this, we are forced into a conundrum—we cannot change the process if the process is based upon faith, not data.
Experiences of various fields, including industry, demonstrate there are other forms of quality control besides peer review that could potentially be utilized in the biomedical journals. These methodologies include 6-sigma, statistical quality control, and web based, consumer driven systems such as that employed by Amazon, eBay, and Slashdot. There are thousands of studies in business and sociology evaluating the decision making process that could be brought to bear to evaluate the decision process at the editorial level, but they have not been used. It would seem very simple to develop randomized trials to determine which system best improves the quality of publication. As Jefferson has pointed out, there are almost no data suggesting that the existing peer review systems work and none to suggest that they are better than any other system. . . .
Based upon the data, we cannot reject the hypothesis that scientific journals are faith based. [Linkov 2006: 598]
Smith 2006. Richard Smith was editor of the British Medical Journal for thirteen years, and writes incisively and wryly about the peer-review process. He states that peer review is “the method by which grants are allocated, papers published, academics promoted, and Nobel prizes won. Yet it is hard to define. It has until recently been unstudied. And its defects are easier to identify than its attributes. Yet it shows no sign of going away.” [Smith 2006: 178]
What is peer review?
[And] who is a peer? Somebody doing exactly the same kind of research (in which case he or she is probably a direct competitor)? Somebody in the same discipline? Somebody who is an expert on methodology? And what is review? Somebody saying “The paper looks all right to me”, which is sadly what peer review sometimes seems to be. Or somebody pouring all over the paper, asking for raw data, repeating analyses, checking all the references, and making detailed suggestions for improvement? Such a review is vanishingly rare. . . .
Robbie Fox, the great 20th century editor of the Lancet, who was no admirer of peer review, wondered whether anybody would notice if he were to swap the piles marked ‘publish’ and ‘reject’. He also joked that the Lancet had a system of throwing a pile of papers down the stairs and publishing those that reached the bottom. When I was editor of the BMJ I was challenged by two of the cleverest researchers in Britain to publish an issue of the journal comprised only of papers that had failed peer review and see if anybody noticed. I wrote back ‘How do you know I haven’t already done it?’
Smith goes on to question what peer review is for and whether it works (not very well, which is no surprise at this point). Its drawbacks are that it’s slow and expensive; it is inconsistent, betraying the myth of being objective and reliable; there is bias, particularly against studies with negative results; it may be abused by reviewers who are competitors; and ideas and text may be plagiarized. Improvements may be made by standardization of the process, blinding reviewers to the identity of authors, feedback to reviewers, training reviewers and other techniques. Nonetheless, the obstacles to shifting a 300-year-old industry are daunting. Smith concludes:
So peer review is a flawed process, full of easily identified defects with little evidence that it works. Nevertheless, it is likely to remain central to science and journals because there is no obvious alternative, and scientists and editors have a continuing belief in peer review. How odd that science should be rooted in belief.
The Advancement of Science: From Paradigms to Peer Review
Paradigms and How They Shift
Understanding the role of paradigms in scientific investigation is one of the keys to approaching the revolutionary view of climate as a problem of ecosystem dynamics as opposed to one simply of excessive greenhouse gases. The new paradigm doesn’t render the old paradigm irrelevant, but it reframes its significance and role in addressing the current climate crisis. It exposes to open examination what was heretofore an invisible phenomenon, and avails a universe of solutions to what is, from the perspective of the greenhouse gas hypothesis, an intractable and quite possibly utterly hopeless problem. Therefore, we will take a moment to review the paradigm process and apply it to our contending climate paradigms.
In 1962, Thomas Kuhn, a Harvard-trained physicist who became a historian and philosopher of science, published a controversial book, The Structure of Scientific Revolutions. Prior to Kuhn, the prevailing assumptions about the way science progressed were that knowledge was gradually accumulated by generations of investigators, with occasional quantum leaps by great scientists, but in an overall smooth and continuous albeit occasionally heroic process.
Kuhn broke new ground by re-examining and reframing the process of scientific investigation.
He brought the term “paradigm” into common usage, by which he meant a body of “universally recognized scientific achievements that for a time provide model problems and solutions to a community of practitioners” (p. viii, emphasis added). We will review Kuhn’s work briefly and apply his analysis when comparing the mainstream greenhouse gas climate paradigm and the newly evolving eco-restoration climate paradigm.
Kuhn maintained that scientific progress is episodic, characterized by long periods of “normal science,” which takes place in the context of a paradigm:
At least in the mature sciences, answers (or full substitutes for answers) to [many] questions . . . are firmly embedded in the educational initiation that prepares and licenses the student for professional practice. Because that education is both rigorous and rigid, these answers come to exert a deep hold on the scientific mind. [Kuhn 1962:5]
Normal science, the activity in which most scientists inevitably spend almost all their time, is predicated on the assumption that the scientific community knows what the world is like. Much of the success of the enterprise derives from the community’s willingness to defend that assumption, if necessary at considerable cost. Normal science, for example, often suppresses fundamental novelties because they are necessarily subversive of its basic commitments. Nevertheless, so long as those commitments retain an element of the arbitrary, the very nature of normal research ensures that novelty shall not be suppressed for very long. [Kuhn 1962:5]
Normal science is punctuated by the appearance of anomalies which cannot be explained by the paradigm’s generally accepted theories, nor tested by what the paradigm might consider reasonable hypotheses, nor resolved with current testing protocols or equipment.
When examining normal science . . . we shall want finally to describe that research as a strenuous and devoted attempt to force nature into the conceptual boxes supplied by professional education. [Kuhn 1962:5]
[W]hen [normal science repeatedly goes astray] – when, that is, the profession can no longer evade anomalies that subvert the existing tradition of scientific practice – then begin the extraordinary investigations that lead the profession at last to a new set of commitments, a new basis for the practice of science. The extraordinary episodes in which that shift of professional commitments occurs are the ones known in this essay as scientific revolutions. They are the tradition-shattering complements to the tradition-bound activity of normal science. [Kuhn 1962: 6]
and
Normal science consists in . . . an actualization achieved by extending the knowledge of those facts that the paradigm displays as particularly revealing, by increasing the extent of the match between those facts and the paradigm’s predictions, and by further articulation of the paradigm itself.
Few people who are not actually practitioners of a mature science realize how much mop-up work of this sort a paradigm leaves to be done or quite how fascinating such work can prove in the execution. And these points need to be understood. Mopping-up operations are what engage most scientists throughout their careers. They constitute what I am here calling normal science. Closely examined, whether historically or in the contemporary laboratory, that enterprise seems an attempt to force nature into the preformed and relatively inflexible box that the paradigm supplies. No part of the aim of normal science is to call forth new sorts of phenomena; indeed those that will not fit the box are often not seen at all. Nor do scientists normally aim to invent new theories, and they are often intolerant of those invented by others. Instead, normal-scientific research is directed to the articulation of those phenomena and theories that the paradigm already supplies. [Kuhn 1962:23-24, emphasis added]
and
Paradigms gain their status because they are more successful than their competitors in solving a few problems that the group of practitioners has come to recognize as acute. To be more successful is not, however, to be either completely successful with a single problem or notably successful with any large number. The success of a paradigm . . . is at the start largely a promise of success discoverable in selected and still incomplete examples. [Kuhn 1962:23, emphasis added]
Even today, over half a century after Structures was originally published, normal science seems immune to the possibilities of paradigm shifts – such thoughts often do not occur until forced, even though the process should be reasonably well known if not entirely understood or accepted. The prevailing opinion about paradigm shifts (if there is any opinion at all) appears to be, “It doesn’t apply to my paradigm.”
In general, a paradigm shift doesn’t only involve “objective” factors, it touches scientific practitioners at a deep emotional level as well, as any participant in or observer of academic dispute can testify:
Scientific fact and theory are not categorically separable, except perhaps within a
single tradition of normal-scientific practice. That is why the unexpected discovery is not simply factual in its import and why the scientist’s world is qualitatively transformed as well as quantitatively enriched by fundamental novelties of either fact or theory. [Kuhn 1962:7]
Therefore, the transition to a new paradigm is disruptive and challenging:
The transition from a paradigm in crisis to a new one from which a new tradition of normal science can emerge is far from a cumulative process, one achieved by an articulation or extension of the old paradigm. Rather it is a reconstruction of the field from new fundamentals, a reconstruction that changes some of the field’s most elementary theoretical generalizations as well as many of its paradigm methods and applications. During the transition period there will be a large but never complete overlap between the problems that can be solved by the old and by the new paradigm. But there will also be a decisive difference in the modes of solution. When the transition is complete, the profession will have changed its view of the field, its methods, and its goals [Kuhn 1962:84-85].
The case in point here is the comparison between old and new climate paradigms
Paradigm step
Old paradigm (greenhouse gases)
New paradigm (Eco-restoration)
Paradigm fundamentals
CO2 and equiv are greenhouse gas blankets and elevated levels cause global warming, primarily caused by burning fossil fuels
Destruction of billions of acres of land interferes with carbon and water cycles, along with oxidation of soils for over 10k years, puts gigatons of carbon into atmosphere
Weaknesses intrinsic to paradigm
Positive feedbacks underrepresented, overlooked, not calculated or estimated;
biology is characterized as passive victim of climate change
Complex, interdependent systems that are difficult to model and to quantify into policy
Strengths intrinsic to paradigm
Amenable to modeling; yields numeric targets that can be translated into policy
Comprehensive of all likely drivers and their theoretical interdependencies. Plausible upon examination of biogeologic history.
Primary investigators
Physical scientists almost exclusively from academia
Restoration ecologists and others from biological sciences; non-academic land managers
Tools
Emissions reductions via alternative energy and elimination of carbon emissions sources
Photosynthesis and regenerative land management
Costs
High
Low
Technology requirements
Extensive
Minimal
Locus of investigation
Centralized in academia – universities, scientific journals, formal test sites
Based first in local land management practice, then investigated by academia, landscape managers, local practitioners – farmers, ranchers, horticulturalists, permaculturists, indigenous cultures, etc.
Weight of evidence
Formal studies, isolated variables
Practical results, holistic assessment of land health, biodiversity, water and carbon cycling
Success criteria
Reduced emissions and atmospheric carbon burdens (target 350 ppm? lower?)
Increased biodiversity, improved water cycles, land resilience, cooling of local biospheres on a global scale, reduced floods and droughts, decline in atmospheric carbon burdens (target 280 ppm)
Duration of existence of paradigm
Roughly 200 years
Roughly 20 years with some roots going back considerably longer
The Perils of Peer Review
It may well behoove us all, including the scientists among us, to take a careful look at how science works today and how practices may improve. For example, while peer review can be a powerful tool, it is worth keeping in mind that we’re in a world of shifting paradigms where there are libraries full of peer-reviewed papers in scientific landfills, review processes notwithstanding. Peer review may be a useful tool, but it may also be a significant obstacle to scientific progress.
Beyond the routine aging and demise of most scientific papers, however, is the problem of a system that excludes information that conflicts with or is invisible to the dominant paradigm. That is a central issue that we face when introducing an entirely new view of climate – in the normal course of science, taking a generation or two to transition between paradigms is acceptable, even healthy; in the throes of a climate emergency, conventional peer review may be a serious obstacle to progress.
Peer reviewers get to review their peers because they are thinking along similar lines, and are likely – perhaps even required – to reject ideas outside mainstream boundaries of thought. Even the conventional authors of a recent USDA study had a difficult time getting their study published because its results were so unexpected [Ausmus 2014; Follett 2012]. And that doesn’t begin to touch any of the forces in the political and economic realms that impact peer-reviewed science, including what actually gets studied (and funded!) and what does not.
Presence or absence of peer review should therefore not serve as a standard for accepting the validity of any paper or report; only the evidence is the basis for such decisions, whether it is within “acceptable” range or far beyond it. The evidence must stand for itself, and the professionals who are readers should have the opportunity to make up their own minds. Peer review is a standard, but not a gold standard – it is one among many, and practitioners of scientific method have an obligation to evaluate the relevance of standards.
Linkov 2006. Whereas most tools of science have evolved over the past three hundred years, there is one that stubbornly shows its age: the scientific journal. The author “argue[s] that the primary reason that journals have not changed is that they are ‘faith based’: we believe in them, we dare not question them.” [Linkov 2006: 596]
Linkov suggests that the journal hasn’t transformed into a new model of publication because it has never applied the scientific method to itself.
Jefferson recently presented an outstanding review of peer review and could find only 19 studies on peer review that were scientifically sound. We could find only 14 articles examining the editorial board/editorial decision making. Thus, with over 50 million articles and 300 years of the traditional journal approaches, there has been only a handful of studies questioning or testing the journal process itself. We scientists keep using the process without question, but with no data to show that it is effective. There is thus no evidence-based approach to the science of research communications. Recent studies reveal that peer review often misses major methodological problems in articles. No wonder it has not changed or improved, as there are no data questioning the process. Hypothesis testing research and randomized trials could easily and cheaply be initiated to understand the ‘grand challenges’ of research communication, but sadly they have not.
Isn’t it strange that three features that are inherent to research communication have not been looked at scientifically? There are several possible reasons for this. The most likely is that we scientists have almost complete faith in the journal process as right and unassailable. We thus take a ‘faith based’ approach to research communications. Faith is defined as a firm belief in something for which there is no proof. Many of us might view questioning of the journal process as an attack on science itself. Clearly, the scientific journal process is not a part of the scientific method. We are taught early in our training about the importance of learning to write articles (e.g. IMRaD[23]), the power of peer review and a belief in the editorial system. We do not question the process, despite the fact that the essence of science is questioning. Questioning peer review is like questioning the Bible, Quran or Torah. One role of science is to help separate science from dogma, which we should now do with journals, and avoid a faith based approach. New approaches need to be taken – you cannot teach dogma new tricks! [Linkov 2006: 597]
It is the scientific method that is central to science, not the scientific journal. The scientific method should be central to other research communication processes, but it is not and has not been used to continuously improve how we communicate research. Because of this, we are forced into a conundrum—we cannot change the process if the process is based upon faith, not data.
Experiences of various fields, including industry, demonstrate there are other forms of quality control besides peer review that could potentially be utilized in the biomedical journals. These methodologies include 6-sigma, statistical quality control, and web based, consumer driven systems such as that employed by Amazon, eBay, and Slashdot. There are thousands of studies in business and sociology evaluating the decision making process that could be brought to bear to evaluate the decision process at the editorial level, but they have not been used. It would seem very simple to develop randomized trials to determine which system best improves the quality of publication. As Jefferson has pointed out, there are almost no data suggesting that the existing peer review systems work and none to suggest that they are better than any other system. . . .
Based upon the data, we cannot reject the hypothesis that scientific journals are faith based. [Linkov 2006: 598]
Smith 2006. Richard Smith was editor of the British Medical Journal for thirteen years, and writes incisively and wryly about the peer-review process. He states that peer review is “the method by which grants are allocated, papers published, academics promoted, and Nobel prizes won. Yet it is hard to define. It has until recently been unstudied. And its defects are easier to identify than its attributes. Yet it shows no sign of going away.” [Smith 2006: 178]
What is peer review?
[And] who is a peer? Somebody doing exactly the same kind of research (in which case he or she is probably a direct competitor)? Somebody in the same discipline? Somebody who is an expert on methodology? And what is review? Somebody saying “The paper looks all right to me”, which is sadly what peer review sometimes seems to be. Or somebody pouring all over the paper, asking for raw data, repeating analyses, checking all the references, and making detailed suggestions for improvement? Such a review is vanishingly rare. . . .
Robbie Fox, the great 20th century editor of the Lancet, who was no admirer of peer review, wondered whether anybody would notice if he were to swap the piles marked ‘publish’ and ‘reject’. He also joked that the Lancet had a system of throwing a pile of papers down the stairs and publishing those that reached the bottom. When I was editor of the BMJ I was challenged by two of the cleverest researchers in Britain to publish an issue of the journal comprised only of papers that had failed peer review and see if anybody noticed. I wrote back ‘How do you know I haven’t already done it?’
Smith goes on to question what peer review is for and whether it works (not very well, which is no surprise at this point). Its drawbacks are that it’s slow and expensive; it is inconsistent, betraying the myth of being objective and reliable; there is bias, particularly against studies with negative results; it may be abused by reviewers who are competitors; and ideas and text may be plagiarized. Improvements may be made by standardization of the process, blinding reviewers to the identity of authors, feedback to reviewers, training reviewers and other techniques. Nonetheless, the obstacles to shifting a 300-year-old industry are daunting. Smith concludes:
So peer review is a flawed process, full of easily identified defects with little evidence that it works. Nevertheless, it is likely to remain central to science and journals because there is no obvious alternative, and scientists and editors have a continuing belief in peer review. How odd that science should be rooted in belief.
Note: As mentioned in the Release notes, we have a small staff, and therefore have had to postpone some important material for the next release, scheduled for January 2018. This is particularly true of forests and we will include a more thorough examination of their importance in addressing climate moving forward. Nonetheless, we felt that the investigations here were innovative and interesting, and we wanted to make them available to our readers sooner rather than later.
Forests cover nearly 31% of Earth’s total land area [FAO 2016], and remain one of the major terrestrial ecosystems on the planet. Forests play a significant role in the global ecosystem through cooling, evapotranspiration, covering/shading/sheltering, providing fuel and fiber, aiding cloud formation, and creating wind. Because global forests and wooded lands store an estimated 485 Gt of carbon, forest conservation and afforestation are recognized in the United Nations Framework Convention on Climate Change (UNFCCC) as key strategies for climate change mitigation [UNFCCC 2017].
Despite this acknowledgment, “for the world as a whole, carbon stocks in forest biomass decreased by an estimated 0.22 Gt annually during the period 2011–2015. This was mainly because of a reduction in the global forest area” [UNFCCC 2017].Indeed, humanity has been in the business of clearing forests for thousands of years, and this continues today. However, rapid reductions in deforestation could abate further carbon emissions and thus extreme results of climate change. Moreover, reductions in deforestation and implementation of agroforestry practices together could restore biodiversity in damaged ecosystems, repair local and global water cycles, and, ultimately, help restore carbon levels to pre-industrial levels. Here we present several articles illustrating the impact of forests on global climate, as well as the potential for restorative afforestation and agroforestry practices to sequester large amounts of carbon.
Forest Article Summaries
Ellison 2017. This paper takes the innovative and paradigm-shifting position that carbon is not the primary consideration in climate; rather, it is water that should be a central focus in assessing climate processes and effects. It considers forests from a systems perspective.
Forest-driven water and energy cycles are poorly integrated into regional, national, continental and global decision-making on climate change adaptation, mitigation, land use and water management. This constrains humanity’s ability to protect our planet’s climate and life-sustaining functions. The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. The effects of tree cover on climate at local, regional and continental scales offer benefits that demand wider recognition. The forest- and tree-centered research insights we review and analyze provide a knowledge-base for improving plans, policies and actions. Our understanding of how trees and forests influence water, energy and carbon cycles has important implications, both for the structure of planning, management and governance institutions, as well as for how trees and forests might be used to improve sustainability, adaptation and mitigation efforts. [Ellison 2017: Abstract]
Ford 2017. Structural Complexity Enhancement (SCE) is part of a larger ecological concept: nature tends to complexity, providing its resiliency, flexibility and inventiveness. SCE in treatment of forests is a management approach that promotes development of late-successional structure, including elevated levels of coarse woody debris. It adds variety to tree ages (favoring older trees), and variations in available sunlight and habitat.
Large trees, previously assumed to slow in both productivity and growth rate (Weiner and Thomas 2001, Meinzer et al. 2011), function as long-term carbon sinks (Carey et al. 2001). These findings further support the significance of structural retention as a co-benefit to forest carbon storage. Adaptive silvicultural practices promoting multiple co-benefits, for instance, by integrating carbon with production of harvestable commodities, can contribute to efforts to dampen the intensity of future climate change while maintaining resilient ecosystems (Millar et al. 2007). Prescriptions that enhance in situ forest biomass and thus carbon storage offer one such alternative (Ducey et al. 2013). U.S. forests currently offset approximately 16% of the nation’s anthropogenic CO2 emissions, but this has the potential to decline as a result of land-use conversion and lack of management (EPA 2012, Joyce et al. 2014). While passive or low-intensity management options have been found to yield the greatest carbon storage benefit, assuming no inclusion of substitution effects (Nunery and Keeton 2010) or elevated disturbance risks (Hurteau et al. 2016), we suggest the consideration of SCE to enhance carbon storage. Multiple studies have explored co-benefits provided by management for or retention of elements of stand structural complexity, including residual large living and dead trees, horizontal variability, and downed CWM (Angers et al. 2005, Schwartz et al. 2005, Dyer et al. 2010, Gronewold et al. 2012, Chen et al. 2015). Silvicultural treatments can effectively integrate both carbon and late-successional biodiversity objectives through SCE based on this study and previous research (e.g., Dove and Keeton 2015). Remaining cognizant of the potential for old-growth compositional and structural baselines to shift over time and space with global change—climate impacts on forest growth and disturbance regimes, altered species ranges, and the effects of invasive species—will be important for adaptive management for late-successional functions such as carbon storage. [Ford 2017: 16]
Healing Harvest Forest Foundation.
The spot compaction of animal feet is far less damaging to the forest soil and tree roots than the continuous track created by a wheel or track driven machine. Small sized tracts of timber can not be harvested with conventional methods that require higher capitalization and expensive moving cost. The economic pressure in conventional forest harvesting operations influences most loggers to feel that they must cut all the trees to make their work cost effective. This restricts the silvicultural prescriptions available for the management of the forest….Our method of selecting individual trees on a “worst first” basis and limiting removal to no more than 30% retains the forested condition and is indeed improvement forestry…. The holes created in the forest canopy are substantial enough for “shade intolerant” species to regenerate naturally from seedlings of the superior specimens that are left in a healthy “good growing” condition. We believe that basically the repair of the forest from previous “high grading” is best accomplished through several successive “low grading” harvests.[Healing Harvest 1999]
Makarieva 2007. The authors examine ecological and geophysical principles to explain how land far inland away from the ocean can remain moist, given that gravity continuously pulls surface and groundwater into the ocean over time.
All freshwater on land originates in the ocean from which it has evaporated, is carried on air flux, and precipitates over the land. Coastal regions benefit from this cycle by their proximity to the ocean, yet in the absence of natural forests in coastal regions precipitation weakens as distance from the ocean increases, leaving inland areas arid. The authors propose the concept of a biotic pump to explain how large continents can be sufficiently moist deep into the interior and abundant with rivers and lakes.
Air and moisture are pulled horizontally by evapotranspiration from coastal forests. When water vapor from plants condenses, it creates a partial vacuum which pulls water evaporating from the ocean into the continental interior where it rains in forest. By contrast, deserts are unable to pull ocean evaporation to them because they lack any evaporative force.
Such ongoing deforestation, and crucially coastal deforestation on a large scale, threatens to cut off rain to the interiors of Earth’s continents thereby creating new deserts. The Amazonian rainforest is the prime example. Deforestation of the eastern coast of South America has led to changes in the rainforest that is resulting in drying and desertification of the interior, with unprecedented fires and loss of rivers. Historically, Australia’s interior became a desert around the time the first humans arrived on the continent, and the authors speculate that early coastal deforestation was the cause. On the other hand, restoring natural coastal forests can also restore inland water cycles and reverse desertification.
This article illustrates the importance of biological relationships that are ecologically complex and poorly understood. It highlights the significance of the precautionary principle in assessing what we don’t know (and what we don’t know that we don’t know) when altering ecological processes, and taking preventive action in the face of uncertainty.
Grasslands have been estimated to cover approximately 40% of global land surface area, approximately 5.25 bn ha (~13 bn ac ), except for Greenland and Antarctica [Suttie 2005; White 2000:12]. Their deep soils are rich repositories of nutrients, especially carbon and water. Many grasslands are anthropogenic, i.e., resulting from various land management techniques to maintain land for grazing and crop production by humans. Virgin grasslands are increasingly rare, possibly leading to significant underestimations of their potential positive contribution to productivity, and to carbon and water storage. Grasslands are important repositories of biodiversity, and have significant impacts on weather and climate. Here we review research and articles that indicate soil carbon storage potentials of roughly 13 gigatons per year (the equivalent of 6.5 ppm) if global grasslands were managed regeneratively.
Overview
While we have separate sections for Grasslands, Croplands and Soils, there is inevitable overlap. Many croplands are modified grasslands, and both are, of course, based in soils. Yet there are enough differences in each area of study to merit separate sections, keeping in mind that systemic behaviors and interactions are broadly applicable.
Since the onset of agriculture over 10,000 years ago with land management techniques that expose soil to air, estimates of up to 537 gigatons of soil carbon have been oxidized to carbon dioxide and other greenhouse gases [Buringh 1984: 91]. Even so, soils (>2,300 Gt) currently hold almost as much carbon as plants (550 Gt), atmosphere (800 Gt) and ocean surface waters (1,000 Gt) combined [NASA 2011], and almost surely retain the potential to store enough atmospheric carbon to return to pre-industrial levels.
Typical soil studies examine the first 30 cm (1 ft) of soil depth, but more recent investigations indicate that major soil carbon storage takes place deeper than that, often in a more stable form [Liebig 2008, Follett 2012, Harper 2013]. A USDA paper found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr) [Follett 2012]. A study of switchgrass for bioenergy found rates of SOC increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft) [Liebig 2008]. On an intensively grazed former row-crop agriculture land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions [Machmuller 2015]. In addition, the ultimate methane emissions may have been markedly less than measured since the report did not consider methane breakdown into CO2 from methanotrophic bacteria and atmospheric hydroxyl radical oxidation, with a significant reduction of methane’s ultimate greenhouse gas impacts.
These reports demonstrate the potential for massive amounts of soil carbon storage, significant cooling of the biosphere, and dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Grassland Evolution
Grasslands have long been a rich repository of carbon, both stable and labile. The co-evolution of grasslands with grazing ruminants has contributed to dramatic global cooling over the past 50 million years as a result of significant photosynthetic carbon drawdown into grassland soils [Retallack 2013]. Thus, grasslands are more than a consequence of geophysical changes, they are
. . . a biological force in their own right (Retallack 1998), in some ways comparable to the human rise to dominance of planetary resources (Vitousek et al. 1997). Grasslands have long been considered products of the coevolution of grasses and grazers (Kovalevsky 1873). Few plants other than grasses can withstand the high-crowned, enamel-edged teeth and hard hooves of antelope and horses. Yet these same animals are best suited to the abrasive gritty opal phytoliths and dust of flat, open grasslands. Grasses recover readily from fire and nurture large herbivores such as elephants: both fire and elephants promote grassland at the expense of wood land (Retallack 1997b; Jacobs et al. 1999). Grasses suppress insect and fungal attack with secondary metabolites such as cyclic hydroxamic acids (Frey et al. 1997). Grasses create Mollisols, unique soils with fine crumb clods rich in organic matter (Retallack 1997b). (Retallack 2001:407) [Emphasis added.]
Occupying such vast areas of planetary land surface, grasslands have a major influence on the global climate:
CO2 and CH4 (which rapidly oxidizes to CO2 ) are important greenhouse gases, and mechanisms for burial of their C may result in climatic cooling (Berner 1999). The most important long-term C sink from grasslands is their supply by erosion to sedimentary basins of crumb peds, which are unusually rich in organic matter intimately admixed with clay (Pawluk and Bal 1985). Tropical forests, in contrast, yield highly oxidized spherical micropeds with virtually no organic content (Retallack 1991a).” (Retallack 2001:414)
While there are other soils that are greater carbon sinks, such as peat bogs, wetlands and coastal habitats (e.g., mangroves, seagrasses), for volume and depth of carbon storage on vast areas of land, grasslands have enormous potential:
Grasses themselves are C sinks, especially considering their mass of roots and rhizomes underground. . . . [Numerous soil investigations] indicate that past estimates of organic C in tropical grassland soils have been low, in part because soils were not analyzed to sufficient depths. . . . Grassland and woodland soils may have comparable amounts of organic C in the surface 15cm. Beyond that depth, organic C values drop off dramatically in woodland soils but remain high in grassland soils to a meter or more. The fine structure and fertility of grassland soils is in large part due to this large C reservoir. (Retallack 2001:415)
Conventionally, it is estimated that approximately 40% of global land surface area is grasslands (52.5 million square kilometers, or ~5.25 billion hectares, or ~13 billion acres [Suttie 2005]), except for Greenland and Antarctica [White 2000:12; see Figure 1, below]. This is likely a significant underestimation of soil surface area and volume, since grasslands are not uniformly flat, with topographical variations adding carbon, water, etc. to areas that are typically calculated on the basis of a two-dimensional map projection [Blakemore 2016: fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regeneration, carbon capture and water storage than is conventionally assumed (see the section, “Do We Have More Soil for Carbon Storage than We Thought?”).
Natural grasslands are typically areas of low and seasonal rainfall. Unlike temperate environments with year-round rainfall, semi-arid and arid grasslands are dependent on grazing animals as a keystone species. The habits of grassland plants are as dependent on grazing animals as the animals are dependent on the plants that grasslands provide as food.
Figure 1: Global Extent of Grassland (White 2000:12).
Grazing animals are ruminants and as long as they are grazed in herds that move frequently, as they do in natural habitats in the company of predators, ruminant species are often interchangeable insofar as grassland health is concerned. Bison and antelope, for example, may be the wild ruminants that roamed the prairies and savannahs, but domestic cattle, when properly managed (human herders are the equivalent of wild predators), serve the same ecosystem functions.
Pioneering Work Of Allan Savory
Allan Savory, a wildlife biologist from Zimbabwe, began studying desertification in the 1950s. He pioneered an approach that he has termed Holistic Planned Grazing (HPG) for regenerative management of grasslands. He noted that there are essential differences between temperate grasslands, which he termed “non-brittle” environments, and arid and semi-arid grasslands, which he referred to as “brittle.” These distinctions are critical in understanding how different habitats require different management approaches.
Non-brittle environments, because of year-round rainfall, are relatively forgiving of mismanagement that destroys soil biota and exposes soil to sunlight, air and the elements. Recovery from soil degradation can be be rapid. Brittle environments, to the contrary, are fragile and easily desiccated, and when poorly managed, either from overgrazing or undergrazing, may take decades or centuries to recover or even ultimately turn to desert.
Brittle environments particularly need ruminant evolutionary partners. Their hooves are designed to open the soils to air and water, and their digestive systems deposit a feast for soil organisms. The ruminant gut is a moist refuge during the dry season for soil microbes which are essential to the health of the land. Grasses need to be bitten lest they shade out their own new growth.
What Savory discovered is that the same land may either flourish or die depending on how it is grazed. When ruminants are kept in check by predatorsthey graze an area in tight herds for protection and then move to the next patch of fresh grasses and other plants, providing up to two years of recovery and regrowth time for the recently grazed pasture. On the other hand, when they are provided the safety of fencing and left to graze large areas at will, they return to their favorite plants and overgraze those areas, eventually compacting the soil, preventing water infiltration and proper aeration, killing the plants, and leading to desertification.
The difference in land health is dramatic.[19] The pictures below illustrate:
Mexico
Arizona
Zimbabwe
Fig. 1. These pictures are of neighboring properties in Mexico, Arizona and Zimbabwe. In each area they were taken on the same day, have similar soils, and the same precipitation. The pictures on the right are examples of properly managed livestock through Holistic Planned Grazing to restore grasslands. On the left we see examples of improperly managed livestock as well as exclusion from grazing (“resting the land”) [Savory Institute 2015:12]
Savory’s work, after decades of successful application on ranches in Africa, Asia, Australia and North and South America, garnered global attention (and controversy) after his TED Talk in 2013 [Savory 2013].
The primary point is that If well-managed, grassland soils can not only sequester annual greenhouse gas emissions but can alsobegin to draw down legacy atmospheric carbon as well. In addition, they provide human and other predator food, converting grasses inedible to non-ruminant mammals to meat.
Grasslands As Ecosystems
It is useful to understand how grasslands work as intact ecosystems, thereby providing a solid theoretical basis for observations of grasslands as vast carbon and water sinks.What appears above-ground is only a hint of grassland ecosystem dynamics. Soils are the planet’s most complex and least understood terrestrial ecosystem, yet soils are where most of the action takes place on grasslands as well. While all soils are built on the foundation of minerals provided by weathering of rock, the soils on grasslands are primarily biological soils. The kingdoms of life are the active agents in soil creation and it is the interactions among life forms that create the rich and productive grassland soils (see Soils section).
The basis for all life is the microbial kingdom. These smallest of cells, with their complex biochemistry, morphology and behavior, are active players in creating stable soil molecules, storing abundant carbon and water. In addition to microbes, soil ecosystems are built from exchanges among fungi, insects, worms, green plants, birds and small and large mammals. It is this set of rich interactions that creates the biodiverse, abundant and resilient environment of global grasslands.
For the sake of illustration, let us start the discussion with the ruminant gut during a dry season. Microbes survive in a warm, moist environment while constantly being cycled into the soil through digestion and elimination. The short-term hoof disturbance with minimal compaction while animals are constantly moving opens the soils to available moisture from precipitation, urination and condensation. During the rainy season the water is more effectively absorbed into opened soils, nourishing plants, raising the water table and eventually even leading to perennial streams and ponds. Even limited rainfall goes a long way in spongy soils that are covered with grasses and other plants to keep the ground cool and moist [Byck 2014: 8’38”]
Methane
Methane, a relatively short-lived but powerful greenhouse gas, is often raised as a serious concern with beef production. This is surely true when animals are left to roam freely and overgraze, and then moved to concentrated animal feeding operations with large manure lagoons. However, it is important to consider the whole of ecosystem functions in assessing methane emissions {Savory Institute 2015]. This includes the conversion of the methane molecule into carbon dioxide by bacteria (metanotrophs) that live in healthy soils and literally eat energy-rich methane, and the oxidation of methane by hydroxyl radicals present in the lower atmosphere. It may include other ecosystem processes, such as the effects of earthworms increasing methanotrophic bacterial activity in landfills and pastures [Héry 2008; Kernecker 2014]. The result is a virtuous cycle where plants can then take up that carbon dioxide through photosynthesis and send some of the carbon back underground through their root systems. Throughout their life cycle under conventional industrial management, cattle are rarely if ever exposed to such healthy, biodiverse soils.
Historical methane data indicates that in the United States, for example, pre-settlement wild ruminants generated approximately 86% of the methane of current farmed ruminants (Hristov 2012:1371). Yet even with vast numbers of ruminants on grasslands across the planet, atmospheric methane remained constant until the global dependence on widespread use of fossil fuels and its effects on agriculture and animal husbandry began to grow rapidly in the 18th century (Fig. 2).
Thus, results are very different with animals grazed in a manner that mimics nature. Rowntree et al. describe the importance of accounting for the beneficial ecosystem services that well-managed grazing systems can provide.
. . . LCA’s [Life Cycle Assessments] often consider soil C to be in dynamic equilibrium. However, empirical data suggest otherwise (e.g. Machmuller et al., 2015; Teague et al., 2011). Recent studies such as Ripple et al. (2014) and Eshel et al. (2014) have reported the emissions from ruminants in food production without accounting for the beneficial ecosystem services that well-managed grazing systems can provide. In our study, we used 3 tC/ha/yr (1.2 tC/ac/yr) as a potential C sequestration figure, which is relatively high (Conant et al., 2001) but viable based on existing studies (Teague et al., 2011; Delgado et al., 2011; Machmuller et al., 2015; Teague et al., 2016). Importantly, the results presented here suggest that with appropriately managed grazing, a grass-finished beef model can not only contribute to food provisioning but also be ecologically regenerative as well. [Rowntree 2016:36]
This excerpt illustrates a paradigm shift in action, wherein investigators within the dominant paradigm are constrained from evaluating the possibilities offered by “beneficial ecosystem services” because it doesn’t occur to them to consider them. Such biological processes are invisible due to limiting assumptions of the paradigm.
Research into systems implications of holistically managed grass-finished beef is growing. It is only recently that mainstream researchers are beginning to understand that the biological function of animals in an ecosystem is as dependent on the ecosystem as it is on the biology of the animal.[20] Studying animals in isolation or as part of a synthetic system such as industrial agriculture often leads to incorrect conclusions.
Fig. 2. Historical Concentrations of Greenhouse Gases [IPPC 2007]
Review of some studies of grasslands
Soil creation (pedogenesis) is conventionally defined as the weathering of rock; it may take 3,000 years or longer to create a foot of soil through geological processes. Soil created through biological activity, on the other hand, happens orders of magnitude faster, up to several inches per year.
Australian soil scientist Christine Jones notes that
The rates of soil formation provided in the scientific literature usually refer to the weathering of parent material and the differentiation of soil profiles. These are extremely slow processes, sometimes taking thousands of years. Topsoil formation is different and can occur rapidly under appropriate conditions. . . .
The late P.A. Yeomans, developer of the Keyline system of land management, recognised that the sustainability of the whole farm was dependent on living, vibrant topsoil. The formation of new topsoil using Keyline principles, at rates not previously considered possible, was due to the use of a tillage implement designed to increase soil oxygen and moisture levels, combined with a rest/recovery form of grazing and pasture slashing, to prune grass roots and feed soil biota, especially endemic earthworms. Yeomans was able to produce 10 cm of friable black soil within three years, on what was previously bare weathered red shale on his North Richmond farm (Hill 2002).
Bennett (1939) calculated a rate of topsoil formation of just over 11 t/ha/yr (4.4 t/ac/yr) for soils in which organic material was intermixed into surface layers. In situations where plant root mass is high, rates of topsoil formation of 15-20 t/ha/yr (6-8 t/ha/yr) have been indicated (Brady 1984). Healthy groundcover, high root biomass and high levels of associated microbial activity, are fundamental to the success of any technique for building new topsoil.
If the land management is appropriate, evidence of new topsoil formation can be seen within 12 months, with quite dramatic effects often observed within three years. Many people have built new topsoil in their vegetable or flower gardens. Some have started to build new topsoil on their farms. If you have not seen new soil being formed, make a point of doing so. (Jones 2003:19-20)
Healthy biodiverse grasslands with abundant animal populations provide favorable circumstances for biological soil accumulation and carbon sequestration, including opening soils to air and water, fertilizing soil life and stimulating growth of grasses.
Using current best land management practices, recent research has begun to confirm the importance of studying soil organic carbon accumulation on grasslands. In a 9-year study of bioenergy crops, investigators found that switchgrass and maize stored 50% of their soil organic carbon (SOC) below 30 cm (1 foot), up to 4 times more than used in models in use at that time (Follett 2012:866):
Most of the research on SOC in agricultural production systems focused on C in the 0 to 30 cm depth [22–27]. A few studies in which soil sampling has been conducted at greater depths indicate that production agriculture affects soil C deeper in the soil profile [28,29]. (Follett 2012:867)
Concerns about soil depth measurement are not new [Liebig 2008]; depth of soil measurement in estimating soil carbon storage potential is a significant issue. Conventional soil science, which largely addresses agricultural soils managed in industrial agricultural contexts, typically measures soil carbon down to around 30 – 40 cm (12 – 16 inches). Yet roots of native prairie plants may reach 5 times that depth (see Fig. 1), storing carbon in stable molecules for centuries and millennia as long as the soils are undisturbed and not exposed to light or desiccation. [See Soils section.]
Methodological issues for assessment of SOC have thus been problematic, and have likely led to serious overall underestimation of soil sequestration capacity. This is particularly relevant because these soils will not be able play their appropriate critical role in addressing climate until mainstream science and policy recognize and promote the potential of best practices in land management in all ecosystems. Harper & Tibbett found up to five times more soil carbon in Australian soils at depths greater than 1 meter (~3 feet) than is conventionally estimated:
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths (Table 1), it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5 m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm −2 or 21 % of the total store to bedrock. [Harper 2013: 645]
We discuss the dynamics of water cycling and forests elsewhere, but it is worth noting a recent paper suggesting that a more holistic view of ecosystem dynamics is in order. A paradigm shift prioritizing water over carbon as the driving climate force more accurately and effectively guides climate recovery strategies and offers more tactical and regenerative options [Ellison 2017; s.a. Schmidt 2017]. Water is more tangible to most people than carbon, and water recovery is more visible and rapid, offering hope and encouragement in a generally grim scenario. In addition there are numerous other benefits to people and landscapes with improved water management.
THE IMPORTANCE OF ANECDOTAL EVIDENCE
While anecdotal evidence is often disdained in academic science, in many scientific pursuits it forms the foundation of future study. This is especially true in such predominantly observational pursuits such as naturalist biology, ecology, rangeland science and agronomy, where study of the visible vagaries of the natural world inspires questions that may reach far beyond what a happenstance occurrence would imply. It is the weight of such observations that leads to formal protocols, hypotheses and theories to explore the details of a field. Isolating variables can be a very helpful tool; unfortunately, modern science practice has generally lost sight of systems contexts, and that system behavior can be very different from the behavior of any of its isolated parts.
Therefore, it is essential to embrace both analytical and holistic evidence in order to build a full understanding of how environmental systems work as wholes. Together these complementary approaches provide a more comprehensive picture of the systems in question, as well as much clearer guidance for how to proceed in current global ecosystem urgencies.
A small selection of a growing literature of informative anecdotal reports of grassland eco-restoration is included among formal studies below. They reflect the extensive experience of farmers, ranchers and other land managers, and demonstrate the potential positive effects of regenerative land management and eco-restoration on climate and the biosphere as a whole. See for example Stigge 2016, Oppenheimer 2015, Byck 2014, Brown n.d., Brown 2016.
Grassland Article Summaries
Byck 2014. This 12-minute video relates the experiences of three ranchers who manage cattle and land according to regenerative land management principles. They discuss their transition to Holistic Planned Grazing, where for two of them, in areas of ~15 inches of rainfall, their alternative had been bankruptcy. The video vividly illustrates the benefits of cover-cropping and organics, the improvement in lifestyle and economics, and the dramatic improvement in biodiversity and water management. [Byck 2014]
Follett 2012. A USDA study found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr). Ausmus reports that
. . . a 9-year project that evaluated the effects of nitrogen fertilizer and harvest treatments on soil organic carbon sequestration in switchgrass and no-till maize crops managed for biofeedstock production [found that] more than 50 percent of the soil carbon was found between 1 and 5 feet below the soil surface. The average annual increase of soil organic carbon throughout the first 5 feet of subsoil also exceeded 0.9 tons per acre per year [Ausmus 2014: 4-5].
Of interest were the difficulties the authors faced in getting the study through peer review and published since their results were so unexpected, as reported in Ausmus 2014.[21] It originally appeared in Bioenergy Research in 2012. [Follett 2012].
Note too that the Follett study was performed on already degraded soils, “Perennial grasses could be used as bioenergy crops on about 20 million ha (ha = 10,000 m2 or 2.5 acres) of marginal or idle cropland in the USA alone [18]” (Follett 2012:867). [Emphasis added.] A well-supported inference is that healthy, biodiverse soils will yield even better results.
Degraded soils may be less effective carbon sinks than virgin soils even though they have lost most of their carbon because the soil life that creates long-lasting stable carbon molecules is damaged or destroyed by synthetic inputs, tilling and other forms of mismanagement. Nonetheless, results were dramatic:
In the first 9 years of a long-term C sequestration study in eastern Nebraska, USA, switchgrass and maize with best management practices had average annual increases in SOC per hectare that exceed 2 tC/yr for the 0 to 150 cmsoil depth. For both switchgrass and maize, over 50 % of the increase in SOC was below the 30 cm depth. SOC sequestration by switchgrass was twofold to fourfold greater than that used in models to date which also assumed no SOC sequestration by maize. (Follett 2012:866) . . . .
Our results clearly show that significant amounts of C were sequestered deep in the soil profile by switchgrass grown and managed as a biomass energy crop and maize grown continuously in a no-tillage production system for the cultivar Trailblazer array of N fertility and harvest treatments for a 9-year period. For almost all other C sequestration reports used in bioenergy models, studies designed for other purposes were adapted to obtain soil C sequestration estimates, initial soil samples were not available, and control samples were from adjacent fields or non-treatment areas. Our results are supported by similar results reported by Liebig et al. [28] for four switchgrass fields managed with uniform N rates and harvest treatments for 5 years in the USA western Corn Belt. They are supported by the recent work of Varvel and Wilhelm [29] for maize in which significant increases in soil C occurred in soil layers up to 150 cm in depth in maize no-till plots as compared to tilled plots. The soil C that is sequestered deeper than 30 cm is expected to be more stable over time since it is below the tillage zone. Even in the top 30 cm of soil, sequestered C may be stable for extended periods in no-till production systems as reported previously by Follett et al. [33]. (Follett 2012:873)
Harper 2013. This study suggests that the standard sampling depth of 30cm vastly underestimates the global store of soil organic carbon, and thus, presumably, the potential of future soil carbon storage that could result from eco-restoration efforts. “Hypothesizing that SOC retained in soils below the top half metre (in highly weathered deep profiles) would account for the major proportion of SOC in the landscape” [Harper 2013: 642], this southwestern Australia study took deep soil samples in 38 spots across 5 locations. They found that 79% of total carbon store to bedrock occurred below a half meter depth, and 41% occurred below 5 meters depth. “There are two possible sources for the deep carbon; that produced in situ by roots or dissolved carbon that has moved downward from nearer the surface” [Harper 2013: 645]. The sample sites were recently reforested or were under agriculture, and previously had been “covered in a range of xerophytic plants, with root systems that extended to depths of 40 m, such as reported for a Eucalyptus marginata forest” [Harper 2013: 642]. The authors note that more research is needed to understand how deep SOC is affected by land-used changes and climate change.
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths, it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm−2 or 21 % of the total store to bedrock. If this is adjusted to 0.3 m depth, using an exponential function based on the samples in the surface metre, the value decreases to 5.6 kgCm−2. For the individual sites this ranged from 3.6 to 8.0 kgCm−2, or 14 –37 % of the total store. …the surface 5 m contained 16.3±1.38 kgCm−2 or 59 % of the total store to bedrock, with this proportion varying from 47 to 77% across the five sampling locations. The amount of carbon stored in the soils can also be contrasted with the biomass carbon storage of 11.0–16.0kg Cm−2 expected at equilibrium following reforestation for these sites (Harper et al. 2007) and likely previously removed from the sites by deforestation in advance of agriculture [p.645].
Liebig 2008. A study of switchgrass for bioenergy found rates of SOC (Soil Organic Carbon) increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft). “In this study, switchgrass significantly affected change in SOC. . . Across sites, SOC increased significantly at 0–30 cm (1 ft) and 0–120 cm (4 ft), with accrual rates of 1.1 and 2.9 tC/ha (0.44 and 1.16 tC/ac), respectively.” [Liebig 2008:215] This indicates the chronic underestimation of soil carbon capacity in the many studies which by convention only measure SOC down to 30 cm (1 ft).
Machmuller 2015. On intensively grazed former row-crop agricultural land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions. The study “sought to determine how fast and how much soil C accumulates following conversion of row crop agriculture to management-intensive grazed pastures in the southeastern United States. . . . The highest rates of belowground C accumulation occur when land is converted to grassland ecosystems” [Machmuller 2015: 2]. These intensively grazed managed systems led to an approximately 75% increase in soil carbon within six years,
[a] high C accumulation rate [that] stems from year round intensive forage/grazing management techniques on sandy soils with an initially low soil C content due to past conventional-till row crop agriculture. . . . These forage-management techniques are precisely those suggested to increase SOM in pasture systems and when they are applied to soils with degraded SOC content, such as soils in the southeastern United States, rapid C accumulation ensues. . . .
On the basis of a whole farm C-cycle analysis, C accumulation appears to offset methane emissions during the rapid soil C accumulation phase . . . As the C accumulation rate declines these farms will become net C-emitting—similar to all dairy production—because of ruminant methane emissions. However, the substantial soil-quality benefits of higher organic matter remain and will likely increase the sustainability of dairy production using management-intensive grazing [Machmuller 2015:3].
The eventual methane emissions may be markedly less than suggested, however, since the report did not consider methane breakdown from methanotrophic bacteria and atmospheric hydroxyl radical oxidation.
The authors conclude
that pasture-based intensively grazed dairy systems may provide a near-term solution for agricultural lands that have experienced soil-C loss from previous management practices. Emerging land uses, such as management-intensive grazing, offer profitable and sustainable solutions to our needs for pairing food production with soil restoration and C sequestration. [Machmuller 2015: 2-3]
McCosker 2000. A discussion of the introduction of what the author calls “cell grazing,” framed as a paradigm shift in Thomas Kuhn’s terms [Kuhn 1962] over the years 1990-1999. McCosker reviews the dichotomy between researchers and producers, and travels to see actual results in the U.S., Zimbabwe, Namibia, and South Africa. He states, “Only after seeing the outcomes time and again in all possible environments was I finally convinced that the principles could not be faulted.” This kind of paradigm shift has been experienced repeatedly as practitioners must develop the courage to challenge prevailing assumptions in order to take the requisite transitional steps.
Oppenheimer 2015. Farmers Paul and Elizabeth Kaiser own eight acres in Sebastopol in Northern California and they farm three of them, developing a model that other farmers are beginning to use.
[Kaiser] farms a mere eight acres, and harvests fewer than three of them. Nonetheless, his methods are at the forefront of a farming movement that is so new (at least in the U.S.), and so built for a climate-changed world of diminishing rains, that it opens up gargantuan possibilities. One might call this methodology sustainability on steroids, because it can generate substantial profits. Last year, Kaiser’s Sonoma County farm grossed more than $100,000 an acre, which is 10 times the average per-acre income of comparable California farms. This includes Sonoma’s legendary vineyards, which have been overtaking farmland for decades, largely because wine grapes have become much more lucrative these days than food, at least the way most farmers grow it.
Kaiser manages all of this without plowing an inch of his ground, without doing any weeding, and without using any sprays—either chemical or organic. And while most farmers, even on model organic farms, constantly tinker with various fertilizer cocktails, Kaiser concentrates on just one: a pile of rotten food and plants, commonly known as compost, and lots of it. Kaiser then adds this compost to a rare blend of farming practices, both old and new, all aimed at returning dirt to the richest, most fertile seedbed possible.
They use permaculture, agroforestry and other intensive techniques, have built deep, healthy soils by keeping the ground covered and spongy to capture water and carbon, all of which provide solid protection from droughts and floods.
Retallack 2001, 2013. “Grassland expansion initiated increased organic C storage in soils, soil water retention, speed of nutrient exploitation, surface albedo, and C burial in sediments eroded from their soils. These changes had many consequences, including long-term global cooling.” [Retallack 2001:422] and “This climatic zone is not only the most widespread, but also the most fertile region of our planet.” [Retallack 2013:78] The paleohistory of grasslands provides the basis for considering the potential of grasslands as huge biological carbon sinks that may be realized again with regenerative land management
Rodale Institute 2014.
Rodale reports that regenerative grazing practiced on a global scale could sequester 71% of annual emissions of 14 Gt C/yr. Combined results from regenerative grazing and agriculture techniques could, if practiced globally, lead to a net reduction of atmospheric carbon dioxide of 1.7 gigatons per year, or 0.85 ppm/year. This represents a potential sequestration of approx. 3t C/ha/yr (1.2t C/ac/yr) on grasslands and croplands. [Rodale Institute 2014:9] This does not include the considerable contributions of non-agricultural lands, nor recent developments in intensive regenerative practices such as permaculture and biochar.
Moreover, Rodale’s side-by-side trial after 30+ years showed that, following the three-year transition period, organic yields match conventional yields, except in drought years, when organic yields surpass conventional yields. Furthermore, energy input and greenhouse gas emissions were lower in organic systems, and profits were higher.
Rowntree 2016. Examining ruminant methane and net carbon sequestration for grassfed beef in a systems context, Rowntree found net sequestration rates of up to 2.11 tons/ha/yr (0.84 tC/ac/yr) for non-irrigated, lightly stocked grazing.
Therefore, including soil carbon sequestration (SCS) potential brings the differences in grazing environments into focus, and significantly changes the outcome. When comparing two well-managed grazing strategies, grass-finished (MOB) and conventional (IRG), each strategy could be an overall carbon sink, but the MOB grazing would only need sequester half as much carbon (1 tC/ha/yr) as IRG grazing (2 tC/ha/yr) for a net zero greenhouse gas footprint. Methane emissions were similar in both grazing environments, but MOB grazing offered significant benefits in increased carbon sequestration. [Rowntree 2016:36]
It is unlikely that such SCS would take place in the absence of a healthy biodiverse ecosystem, one that is supported through MOB grazing. The result is higher net methane emissions under conventional grazing practices. The opposite occurs with properly managed grazing practices, where grasslands as a system actively build more soil carbon for years, leading to a net increase in soil carbon despite enteric methane production by ruminants.[22]
The recent call for improved management of grazing systems as part of an international climate change mitigation strategy is critical, particularly in light of many existing beef LCAs [Life Cycle Assessments] that have concluded that beef cattle produced in grazing systems are a particularly large sources of GHG emissions. To identify the best opportunities to reduce GHG emissions from beef production, a systems approach that considers the potential to increase soil C and reduce ecosystem-level GHG emissions is essential… [W]e generated an LCA that indicates highly-managed grass-finished beef systems in the Upper Midwestern United States can mitigate GHG emissions through SCS while contributing to food provisioning at stocking rates as high as 2.5 Animal Units (AU) per hectare. From this data, we conclude that well-managed grazing and grass-finishing systems in environmentally appropriate settings can positively contribute to reducing the carbon footprint of beef cattle, while lowering overall atmospheric CO2 concentrations. [Rowntree 2016:36]
Schwartz 2013. Cows Save the Planet was a landmark book that helped launch the regenerative agriculture and eco-restoration climate movements. The author gathers stories from practitioners around the world and paints a picture of broad possibilities for addressing global warming, floods, droughts, desertification, malnutrition and many other seemingly unrelated problems that have a single common cause: human mismanagement of lands across the planet. An excellent introduction for a general readership as well as for scientists unfamiliar with the potentials that nature provides.
Shinn n.d. Ridge Shinn is a rancher in Central Massachusetts (Big Picture Beef) who introduced Allan Savory’s methods to the state. For over ten years he has been investigating the holistic context for bringing healthy meat through the local supply chain, and its relationship to human health and global warming. He states,
Since the 1990’s, science has discovered important connections between rotational grazing, soil health, and healthy food. Big Picture Beef’s methods for raising 100% grass-fed cattle offer huge benefits for the environment and for society. The long term goal of the program is Northeast beef for Northeast markets, carbon sequestration, soil fertility and biodiversity, energy savings, and a revitalized rural economy.
Teague et al. 2016. In a review of the literature, the authors conclude that regenerative conservation cropping and adaptive multi-paddock grazing can turn agricultural soils from a carbon source in conventional agriculture into a carbon sink at rate of ~3 tC/ha/yr (~1.2 tC/ac/yr). Key factors include the use of no-till, cover crops, managed grazing, organic soil amendments and biotic fertilizer formulations. These practices can result in elimination of soil erosion and loss, the greatest agricultural contribution to global warming (1 Gt C/yr). Benefits may include “increased water infiltration, improved water catchment, greater biodiversity, increased ecosystem stability and resilience, and improved C sequestration.” [Teague 2016:158]
Conclusion
These reports demonstrate the promising potential for storage of massive amounts of soil carbon to address both the atmospheric and eco-destruction aspects of climate, along with dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Cultivated land covers 1.6 billion hectares globally [FAO 2011]. About 62% of cropland produces food directly for human consumption, while 35% is dedicated to producing animal feed, and 3% to biofuel feedstock, seed and other industrial products [Foley 2011: 338]. Agriculture is a major source of emissions of carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O), contributing 10-12% (including crop and livestock production) of total greenhouse gas emissions [Smith 2007]. Agricultural emissions are driven by the globally dominant industrial model, which favors monocultures and fossil-fuel intensive inputs, and results in soil organic carbon loss and overall soil degradation. However, rather than being a source of carbon emissions globally, agriculture can become a powerful carbon sink. This section looks at the carbon sequestration outcomes of farming practices, such as cover cropping, agroforestry and no-till, which are designed to minimize erosion and boost soil biodiversity, thus restoring soil ecosystems to health and resilience. While more research is needed on holistic approaches that combine multiple soil-building practices, such as permaculture and agroecology, recent research suggests that restorative agriculture could sequester “more than 40% of annual emissions (an estimated 21 Gt CO2 each year [5.7 Gt C/year])” [Rodale 2014], and likely far more, as indicated below.
Cultivation thus began an ongoing slow ignition of
Earth’s largest surficial reservoir of carbon [16]
Overview
The purpose of this compendium, once again, is to emphasize possibilities, the “positive deviants” which lead us to expand our conceptual limits. Only when we can conceive of exceptional and inspiring outcomes may we find the motivation to overcome obstacles to attain them. Fortunately the evidence that supports regenerative land management is rapidly growing, and there are indications that it may outpace climate disruption and provide us with the time and opportunity to address the many difficult circumstances resulting from widespread eco-destruction, including the poster child, global warming. In this section we address the challenges of croplands and their ability to capture atmospheric carbon and recover quickly from millennia of mistreatment.
Under careful human management it is possible for soil organic carbon to reach amounts greater even than under natural, pre-agricultural conditions. A classic example is the Terra Preta soils of the Amazon, “where intensive management and high levels of organic matter additions were practiced over many years, resulting in greatly enhanced soil C” [Paustian 1997: 231].
In spite of a long history of soil carbon loss and a body of scientific literature that views carbon-poor soils as “normal,” many examples of building high levels of soil carbon exist among today’s ecologically minded land managers. California Farmers Paul and Elizabeth Kaiser, for instance, use 5-10 times more compost than average, never till, rotate fields with an extremely diverse mix of vegetable varieties, surround their crops with native trees, shrubs and flowers and have thus built up a thick topsoil containing 10% SOM [Oppenheimer 2015; Kaiser 2017].
In most scientific studies, carbon sequestration rates for croplands measure below 1t C/ha/yr (0.4t C/ac/yr), despite some exceptions as highlighted below. Leading soil scientist Rattan Lal [2016] estimates the global sequestration potential for cropland soils to be 0.8 to 1.2 t C/ha/yr, or “as much as 62 t/ha over the next 50 to 75 years … with a total C sink capacity of ~88 Gt on 1,400 Mha” [Lal 2016: 20A]. That amounts to an average annual global sequestration rate between 1 and 2 Gt C/year, compared to annual carbon emissions from fuel combustion and land use conversion of more than 10Gt C/yr [Lal 2016]. Similarly, Smith [2008] estimates that, under improved management, agriculture could offset 20% of global emissions. Both authors note that conservation-oriented agricultural is a small, albeit crucial, piece of the whole climate mitigation puzzle.
Yet, for a couple of important reasons, these estimates likely greatly underestimate the potential of global croplands to absorb carbon. First, samples are commonly taken to a depth of 30cm or less [Torres-Sallan 2017; Minasny 2017]. This is the default sampling depth recommended in the 2006 IPCC Guidelines for National Greenhouse Gas Inventories, despite acknowledgement in these same guidelines that land use and management is likely to have a major impact on deeper soil layers [FAO 2017b].
Indeed, significant amounts of carbon sequestration occurs in deeper soil profiles – even beyond a 1 m (3 ft) depth [Follett 2012, Liebig 2008, Schmidt 2011: 51]. Harper et al. found that half to three-quarters of total SOC to bedrock was in the surface 5 m with the remainder below that depth. The authors speculate that deep carbon may have been deposited directly by deep-rooting plants. “Where deep soils coincide with deep rooting the biological deposition of carbon from roots (and their associated biota) is inevitable at depths at which SOC has rarely been measured” [Harper 2013: 642].
Second, many studies measure sequestration rates for just one or two soil-building techniques, isolating them from additional, potentially synergistic, practices. In fact, intact ecosystems are based on countless synergistic relationships among organisms and their environment. In other words, many studies measure minor tweaks to conventional, industrial cropping systems.
For example, Minasny et al. [2017] compiled sequestration rates from around the world to assess the viability of the France-led “4 per 1000” initiative (seeking to halt the annual increase in atmospheric CO2 by increasing soil carbon by 0.4% per year). The authors estimate “that an annual rate of 0.2-0.5t C/ha/yr is possible after adoption of best management practices such as reduced tillage in combination with leguminous cover crops.”However, most of some 40 studies of best management practices on arable land assessed only one or two of many – often minimally improved – practices, such as “reduced use of summer fallow,” “rice-rice with NPK,” “inorganic fertilizer,” and “pasture.”
Similarly, an often-cited study by West and Post [2002], compiling 276 paired treatments from 67 long-term experiments, analyzes the sequestration rates for either increased rotation complexity (0.2+/-0.12t C/ha/yr) or a change from conventional tillage to no-till (0.57+/-0.14t C/ha/yr). While both practices were used at some sites, the data were not analyzed according to whether a single practice or combined practices were used. However, the authors suggest that using both practices together can be additive:
Data used in this analysis was stratified separately with regard to a change in tillage or a change in crop rotation. In practice, these changes could occur simultaneously. It can be inferred from our results that if of a decrease in tillage and an enhancement in rotation complexity occur simultaneously, the short-term (15–20yr) increase in SOC will primarily be caused by the change in tillage and subsequent decrease in the rate of SOC decomposition, while the long-term (40–60yr) increase in SOC will be primarily caused by the rotation enhancement and residue input and composition [West & Post 2002: 1943].
If moving to a combination of two restorative practices can increase carbon sequestration somewhat above the use of a single improved practice, then what is possible when many restorative practices are stacked one upon another within an agroecosystem? Permaculture, a design framework with “two broad conceptual criteria: ecosystem mimicry and system optimization,” where multiple restorative practices are indeed combined, represents a counterexample to industrial farming. Yet, sequestration rates from such a system have rarely, if ever, been measured. In fact, very little scientific study of any kind has been conducted in permaculture systems, despite the concept having been developed by scientist Bill Mollison, and adopted to favorable results globally for over 30 years [Ferguson and Lovell 2013].
In light of the centrality of agriculture’s role in ecosystem restoration due to the vast surface area it occupies, we present some literature representing agriculture’s maximum potential contribution to climate change mitigation, and argue that the focus of future research do the same.
Specifically, future studies should consider the effects of greater ecological intensity, diversity and potentially additive and synergistic interactions that can exist among multiple soil-building practices, rather than continuing to pursue measurement of their individual effects, which yield minimal outcomes. Future research must also measure SOC changes to greater depths in the soil horizon in order to capture the full benefit of any given practices. Such changes would likely present both a more accurate and more promising real-world potential for the climate mitigation potential of agriculture.
For a relevant and effective comprehensive assessment of regenerative management practices, one that supports the profound shift necessary in conventional 21st century agriculture, a scientific paradigm shift is necessary so that economics and policy will follow.
For 10,000 years, humans have been clearing patches of forest and grassland to plant crops. While clearing land by burning it visibly turns organic carbon into smoky CO2, plowing and tilling releases soil organic carbon by breaking up soil aggregates that protect carbon. Exposed soil organic carbon is consumed by microbes, and converted to CO2 through respiration. Tilling soil also subjects it to erosion. “Since tillage-based farming began, most agricultural soils have lost 30% to 75% of their soil organic carbon (SOC), with industrial agriculture accelerating these.” [Teague 2016: 157]
Agriculture is a source not only of CO2 emissions, but also of greenhouse gas emissions of methane (CH4) and nitrous oxide (N2O). In cropland soils, CH4 is produced by anaerobic decomposition of organic matter, usually in waterlogged soils like rice paddies. However, soils can also be a methane sink due to the presence of methanotrophic bacteria, which oxidize methane. N2O is produced by microorganisms, which transform excess ammonia fertilizer into nitrate and then N2O. “Upon conversion to NO2– or NO3–, excessive fertilizer N becomes subject to denitrification and thereby contributes to terrestrial emissions of N2O, which have been found to increase with the rate of N fertilization” [Mulvaney 2009: 2296].
Industrial agriculture compensates for soil carbon loss by abandoning degraded land or using chemical inputs for the nutrients and pest resistance that an otherwise carbon-rich, biologically active soil provides. However, the farming methods that rebuild topsoil without relying on synthetic inputs, while also ameliorating the worst effects of drought, are the same ones that can make agriculture a major sink for atmospheric CO2. Such methods, which can be used together as a complementary suite of practices include: no-till; cover-cropping; agro-forestry; diverse crop rotations, including integrating livestock grazing; use of compost, manure, and biochar; and use of deeper-rooting plants and perennials.
Cropland Article Summaries
Cover crops
Cover crops protect the soil during a time of year when no cash crops are growing and the soil would otherwise be bare. “Cover crops, also named inter-crops or catch crops, are crops that replace bare fallow during winter period and are ploughed under as green manure before sowing of the next main crop.” [Poeplau 2015: 34] Cover crops can also be rolled and crimped or mowed, instead of plowed, in preparation for the main crop.
Using cover crops reduces erosion, nutrient leaching, and drought stress, and add carbon through continued plant cover and growth as well as increase biodiversity. Leguminous cover crops also fix nitrogen. Furthermore, “in contrast to other organic amendments, a large part of the C input from cover crop is added as roots, which was found to contribute more effectively to the relatively stable carbon pool than aboveground C-input” [Poeplau 2015: 38].
Vick 2016. This Montana study demonstrates that leaving farmland fallow “depletes carbon stocks and thereby soil quality” [Vick 2016: 129], thus illustrating the importance of keeping land continuously covered with living vegetation. “Fallow” is the stage of crop rotation where no crop is grown.
In this study, a CO2 emissions rate of 1.35 tC/ha/yr (0.54 tC/ac/yr) was measured from land left fallow during the 2014 summer growing season; an adjacent field planted in winter wheat (summer 2013) and spring wheat (summer 2014) was a net carbon sink, measuring carbon input from the atmosphere into the soil at ~2 tC/ha/yr (0.8 tC/ac/yr) and ~1 tC/ha/yr (0.4 tC/ac/yr), respectively. Other parts of this study show a dramatic effect on area cooling as well as increased moisture and rainfall. These results occur only from ending the practice of fallowing.
The researchers observed that a widespread decline of land left fallow in agricultural areas of the Canadian Prairie Provinces coincided with a summertime cooling trend since the 1970s. They noted that extreme temperature events now occur less frequently than in the recent past, maximum summer temperatures have decreased by ca. 2° C (3.6° F), relative humidity has increased by some 7% and summer precipitation has increased by an average of 10 mm/ decade across parts of the Canadian Prairie Provinces. A remarkable 6 W/m2 summer cooling has been observed compared to a ca. 2.5 W/m2 warming globally since the dawn of the Industrial Era.
Even in degraded croplands, relatively small changes can lead to significant differences in rainfall, soil carbon sequestration, and ambient temperature. A 2016 study in Montana demonstrated the effects of reversing the practice of fallowing of wheat fields in the upper midwest. Fallowing is “the practice of keeping a field out of production during the growing season” (Vick 2016:129):
Fallow is a common management practice in the dryland wheat-growing regions of the northern North American Great Plains to conserve water for subsequent crops (Lubowski et al., 2006). Fallow however also increases erosion (Wischmeier, 1959) and soil carbon loss (Cihacek and Ulmer, 1995), and fallow-small grain management strategies are not considered sustainable from the soil conservation perspective (Merrill et al., 1999). [Vick 2016:130].
As a result of farmers’ experience, fallowing has progressively decreased across many areas of the northern midwestern plains since the 1970s, providing an environment suitable for comparison study:
The area of fallow in the Prairie Provinces of Canada has decreased from over 15 Mha in the 1970s to under 2 million ha at the present (Fig. 1) as producers have realized that the water-savings benefit of fallow is outweighed by the economic losses of not planting (Dhuyvetter et al., 1996). The area under fallow in the United States has likewise decreased from 16 Mha to 6 Mha across the same time frame (Lubowski et al., 2006), largely in the northern Great Plains and other areas of the semiarid West . . . Despite the decreasing trend in fallow area across the North American northern Great Plains, fallow remains common in many regions including major land resource area
(MLRA) 52 in north-central Montana – the largest wheat-growing region in the state – where some 40% of agricultural lands may remain in fallow in any given year. In contrast, fallow has been reduced in northeastern Montana (MLRA 53) by hundreds of kha over the past decade (Long et al., 2014, 2013) as producers have adopted continuous cropping or alternate cropping practices. [Vick 2016:130].
The effects of this relatively simple change of practice led to some remarkable results:
The widespread decline of fallow in agricultural areas of the Canadian Prairie Provinces (Fig. 1) has coincided with a summertime cooling trend since the 1970s (Betts et al., 2013a, 2013b; Gameda et al., 2007; Mahmood et al., 2014). Extreme temperature events now occur less frequently than in the recent past, maximum summer temperatures have decreased by ca. 2০C, relative humidity has increased by some 7% (Betts et al., 2013b), and summer precipitation has increased by an average of 10 mm/decade across parts of the Canadian Prairie Provinces (Gameda et al., 2007). A remarkable 6 W/m2 summer cooling has been observed (Betts et al., 2013a); for reference, anthropogenic greenhouse gasses are responsible for a ca. 2.5 W/m2 warming globally since the dawn of the Industrial Era (IPCC, 2007). These climate benefits have only occurred during the growing season; fall, winter, and early spring temperatures have followed global trends (Betts et al., 2013b) . . . In other words, the observed regional climate cooling is broadly consistent with the effects of fallow avoidance on climate processes. [Vick 2016:130-131]
As dramatic as some of these changes are with only reduced fallowing, there are other land-management practices with significant impacts on water cycles, soil carbon, biodiversity and productivity. Such practices hold additional potential, and include cover-cropping and green mulches, pasture cropping, elimination of synthetic inputs which encourage renewed activity of important soil biota, especially worms – and perhaps most importantly on grasslands that co-evolved with animals, the reintroduction of animals themselves.
Finally, it is worth noting that there may be a significant underestimation of surface area and volume of soils in grasslands, as well as in other ecosystems, since natural topographies are not uniformly flat. Topographical variations would add volumes of soil carbon, water, etc. to prior estimates of areas that are typically calculated on the basis of two-dimensional map projections [Blakemore 2016: Fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regenerative management, carbon capture and water storage than is conventionally assumed. Such adjustments to soil volume calculations would positively affect carbon drawdown estimates in considering the potentials of eco-restoration in climate (see section, Do We Have More Soil for Carbon Storage than We Thought?).
Pimentel 2011. Arguing for cover crops as an effective way to reduce erosion and conserve nutrients in soil, Pimentel notes that “Growing cover crops on land before and after a primary crop nearly doubles the quantity of solar energy harvested in the agricultural system per hectare per year. This increased solar energy capture provides additional organic matter, which improves soil quality and productivity.” [Pimentel 2011: 41]
Crop rotation
Crop rotation diversification can enhance pest resistance, nitrogen input (when leguminous crops are added), soil penetration for better water infiltration (when deeper rooting plants are added), and residue input (when crops that produce more biomass are added). The effects on carbon sequestration from increases in crop rotation diversity vary depending on what crops are included. “Crop species can vary significantly in growth patterns, biomass production, water requirements, and decomposition rates, all of which affect net GHG emissions. Therefore, many rotations could be adapted with alternative species or varieties of annual crops to promote soil C sequestration—increasing root and residue biomass, increasing root exudates, or slowing decomposition—or otherwise reduce emissions” [Eagle 2012: 13].
Clearly, crop rotation is something of an umbrella term, describing a variety of practices, and even leaving space for practices that would not seem to offer much in the way of soil restoration. For example, as West and Post [2002] state,
. . . enhancement of rotation complexity refers to (i) a change from monoculture to continuous rotation cropping, (ii) a change from crop–fallow systems to continuous monoculture or rotation cropping, and (iii) an increase in the number of crops used in a rotation cropping system. In this analysis, continuous cropping is a cropping system without a fallow season, monoculture is a system with only one crop grown, and rotation cropping indicates two or more crops rotated over time on the same unit of land. [West & Post 2002: 1931]
Thus, even “continuous monoculture” can be considered as a crop rotation meant to increase carbon sequestration capacity. On the other hand, crop rotation can also involve great diversity, such as at Paul and Elizabeth Kaiser’s farm, where 3-7 crops/year rotate through vegetable beds, sometimes intercropped two crops at a time [Kaiser 2017].
Teague 2016. This study argues for greater use of no-till, cover crops, and crop rotation, including integrating livestock rotation into cropping systems.
Crop production can be managed to maintain permanent ground cover through the rotation of forage and row crop mixes, including cover crops, and legumes to increase soil fertility by fixing N. Grazing livestock can accelerate nutrient cycling through the consumption and decomposition of residual aboveground biomass.” [Teague 2016: 159]
The authors present a set of testable hypothetical scenarios suggesting the adoption of conservation cropping and adaptive management grazing (including grass-finishing cattle).
No-till
No-till (NT) allows farmers to plant without disturbing the soil, thus protecting it from water and wind erosion, leaving soil aggregates intact, and preventing a flush of oxygen from activating microbial breakdown of organic matter and releasing CO2. No-till can contribute to climate mitigation both by reducing emissions from the turnover of soil organic matter caused by tillage, and by sequestering carbon, especially in the surface layer [Mangalassery 2015].
Brown 2016. North Dakota Farmer Gabe Brown began practicing no-till in 1994. Since then, he has added cover crops (a diverse mixture of 70 species), complex crop rotations, orchards, livestock grazing (including cattle, sheep, pork and chicken), vegetable production, and bees. Through a long-term commitment to building the soil through no-till, keeping the ground always covered, and favoring as much biodiversity as possible (including a wide diversity of cash crops), Brown reports SOM has increased from 1.7% in 1993 to 11% in 2013. Furthermore, water infiltration has increased from ½ inch to more than 14 inches over the same time span.
Follett 2012. Measured to a depth of 150 cm (~5 ft), no-till continuous maize grown in eastern Nebraska, fertilized with 120 kg/ha of nitrogen and stover left on the field after grain harvest, sequestered 2.6 tC/ha/yr (1 tC/ac/yr). Notably, more than 50% of sequestered carbon was found below 30 cm (1 ft), illustrating that studies failing to sample below this depth (a common practice) risk greatly underestimating sequestration rates.
Organic vs. synthetic inputs
Organic farming uses “cultural, biological, and mechanical practices that support the cycling of on-farm resources, promote ecological balance, and conserve biodiversity” according to the USDA, which prohibits the use of most synthetic pesticides and fertilizers on certified organic farms. Organic farmers must find alternatives to synthetic inputs for managing pests and fertility. For example, vermi-composting is commonly used in organic farming. It is a natural and proven enhancement of the humification process that uses specific earthworms (e.g. Eudrilus eugeniae [Blakemore 2015]) to rapidly convert all organic “wastes.” Returning this vermicompost to soil renders synthetic fertilizers and pesticides unnecessary, as vermicompost often confers natural resistance to pests [Howard 1945, Balfour 1975] and it enhances resident earthworms [Blakemore 2000, 2016a; see also Earthworms section].
While the organic law provides a baseline for organic practices, the term “organic” encompasses a wide range of approaches to farming. For instance, some organic farmers may do little more than substitute naturally occurring inputs into an otherwise conventional, industrial operation, likely leaving the soil similarly depleted. Other organic farmers put into practice several of the methods mentioned in this section, aiming to truly build the functionality of the soil to resist pests and provide fertility. The studies included below highlight benefits from organic inputs and problems that come with using synthetic fertilizers with respect to soil carbon and biodiversity.
Johnson 2017. Using fungal-dominant compost in a 4.5-year trial at Leyendecker Field Research Site in New Mexico, researchers recorded an annual carbon sequestration rate of 10.7t C/ha/yr (4.8t C/ac/yr). Based on the observed trajectory of increasing productivity, they estimate a potential rate of 19.2t C/ha/yr (7.67t C/ac/yr). Chief investigator David Johnson found that increased plant growth is correlated most closely with the fungal to bacterial ratio. At a fungi:bacteria ratio of 0.04, only 3% of carbon flow went into plant biomass production, with the remainder of the carbon going into other functions, including nitrogen fixation, the soil, and respiration. At a fungi:bacteria ratio of 3.68, plant growth was more efficient with 56% of carbon flow going to biomass production.
Rodale 2014. Compiling data collected from around the world, Rodale Institute concluded that if all cropland were converted to their regenerative model[17], it would sequester 40% of annual CO2 emissions. Adding pastures to that model would add another 71%, effectively exceeding the world’s yearly carbon dioxide emissions.
On-farm soil carbon sequestration can potentially sequester all of our current annual global greenhouse gas emissions of roughly 52 gigatonnes of carbon dioxide equivalent (GtCO2e). Indeed, if sequestration rates attained by exemplar cases were achieved on crop and pastureland across the globe, regenerative agriculture could sequester more than our current annual carbon dioxide (CO2) emissions. Even if modest assumptions about soil’s carbon sequestration potential are made, regenerative agriculture can easily keep annual emissions within the desirable lower end of the 41-47 GtCO2e range by 2020, which is identified as necessary if we are to have a good chance of limiting warming to 1.5°C. [p.5]
Ryals and Silver 2013. This study examined the effects on plant growth and respiration from compost application on annual grassland in both coastal and valley sites in California. They found that a single application of compost during the three-year study resulted in a carbon sequestration rate of 1.45t C/ha/yr (0.58t C/ac/yr) and 0.54t C/ha/yr (0.22t C/ac/yr) at the valley grassland and coastal grassland, respectively. This enhanced net primary productivity was partially offset by CO2 emissions from increased respiration, but the compost did not affect CH4 or N2O fluxes. The authors conclude that:
Our results have important implications for rangeland management in the context of climate change mitigation. Urban and agricultural green waste is often an important source of greenhouse gas emissions (IPCC 2001). Here we show that an alternative fate for that material can significantly increase NPP and slow rates of ecosystem C losses at the field scale. This approach provides important co-benefits to landowners, such as the sustained increase in forage production measured here [Ryals & Silver 2013: 56].
While these results are low compared to some of the other studies noted here, this study illustrates positive use for green waste, and a potential tool that may contribute to climate-positive management.
Khan 2007. This five-decade study of nitrogen fertilization effects on SOC in Illinois shows that, despite progressively greater corn crop residue input during the second half of the 20th Century (increasing from 20,000 or 30,000 to 69,000 plants/ha since 1955), partly due to synthetic fertilizer use, SOC content did not increase, and in most cases declined. SOC declines were most pronounced in subsurface (16-46cm) of the soil horizon, compared to the surface layer (0-15cm). These results are despite crop residue being incorporated, rather than removed, in most plots since 1955, and in all plots since 1967.
These findings implicate fertilizer N in promoting the decomposition of crop residues and soil organic matter and are consistent with data from numerous cropping experiments involving synthetic N fertilization in the USA Corn Belt and elsewhere, although not with the interpretation usually provided. [Khan 2007: 1821]
Perennial systems, agroforestry, and permaculture
Unlike annual plants, perennials live for many years – thousands of years in some cases. Because of their deep (>2m, or 6 ft) and extensive root system, and longer growing seasons, perennials are likely to sequester carbon better than annual cropping systems [Glover 2007].
Agroforestry is the practice of integrating trees (a type of perennial) into a cropping system, including alley cropping, windbreaks, riparian buffers, silvopasture, and forest farming [Eagle 2012; Nair 2009]. Agronomic practices are notable for adding significant amounts of carbon to aboveground biomass, which is often measured separately from soil organic carbon sequestration [Nair 2009]. One of the strengths of agroforestry is its enhancement of an agroecosystem’s functional diversity:
The utilization of the environment by species includes three main components: space, resources, and time. Any species utilizing the same exact combination of these resources as another will be in direct competition which could lead to a reduction in C sequestration. However, if one species differs in utilization of even one of the components, for example light saturation of C3 vs. C4 plants,[18]C sequestration will be enhanced.” [Udawatta 2011: 19]
Toensmeier 2017. Compiling carbon sequestration rates from individual studies, reviews, and expert estimates, and organizing them into groups of annual versus perennial systems, woody versus herbaceous crops, and polyculture versus monoculture, Toensmeier observes that “the general trend is that systems that incorporate trees sequester more carbon.” The highest sequestration rate listed, 18 tC/ha/yr (7.2 tC/ac/yr) falls into the perennial woody polyculture group, and more than half of all sequestration rates listed under perennials are more than 6 tC/ha/year (2.4 tC/ac/yr), while most rates for annual cropping systems are less than 1t C/ha/yr (0.4 tC/ac/yr).
Lawton 2016. On 10 acres of the Arabian Desert in Wadi Rum, Jordan, Permaculture Designer Geoff Lawton built an organic, multi-species food forest on what had previously been bare desert ground. Using wastewater from a nearby irrigated farm to get started, he designed a microclimate that would prevent evaporation in every way possible. Key elements included: date palm trees for wind protection and shade; smaller fruit tree and trellised grapevines for additional shade; a succulent ground cover, which also catches nutrient-rich desert dust; a shaded swale for irrigation; and cut legume trees for mulch.
Lawton sought to “build organic matter within the system as quickly as possible with any living elements that will achieve those ends.” Once the soil came alive, it became productive. Lawton explains that strategic arrangement of the space is especially important in the desert. That’s why crops were grown in two rows in between three slightly wider rows of mixed fruit trees for protection. After four years, this orchard/farm was producing an abundance of fruits and vegetables, showing that it is possible to work with nature and avoid industrial inputs to achieve a productive landscape even in the harshest environment.
DuPont 2010. A Land Institute study measured the effect on soil properties and biota from perennial polyculture systems as compared to annual grain crop systems. Since the latter are typically intensively managed, “the effects of tillage and plant community composition are often confounded” [DuPont 2010: 25]. To control for management effects, this study compared the soil carbon and root biomass outcomes from no-tilled annual crops (rotation of soybean, sorghum and wheat) versus a perennial polyculture. Total root biomass in no-till annual plot measured at only 43% of that in a perennial grass plot in the top 1m of soil. Also, the authors found significantly higher levels of readily oxidizable carbon (ROC) and microbial biomass in the perennial plots compared to the annual crop plots. ROC measures soil carbon that is more available to soil microbes.
Small changes in ROC and other labile fractions of SOC may provide an early indication of soil degradation or improvement in response to management practices. Changes in active carbon pools can be two to four times greater than changes in total C after the initiation of new management practices and they are more highly correlated with other soil quality indicators including microbial respiration, aggregate stability and plant productivity [DuPont 2010: 28].
The authors conclude that “even in the absence of tillage and under best management practices, annual cropping can reduce soil carbon and impact soil biota and food webs important in nutrient cycling after just three years” [DuPont 2010: 25].
Soto-Pinto et al. 2009. In this southern Mexico study of land-use change in various agroforestry systems, the authors show that converting “traditional fallow” (secondary growth woods following cropping, averaging 23.4 years in age) to maize (with beans, squash and pepper) production results in 94% loss of living biomass carbon. However, transitioning to (a) “taungya” (maize, beans, squash and peppers intercropped between rows of timber and multipurpose trees), (b) shaded coffee systems, or (c) “improved fallow” (adding timber trees to traditional fallow plots) preserves living biomass carbon. This study points to the mounting relevance of agroforestry systems that can provide economic benefits to small-scale farmers, while avoiding carbon emissions from land use change from forest to agriculture and livestock production, which accounts for 35% of total emissions in Mexico, according to the authors.
Association for Temperate Agroforestry 2004:
Agroforestry practices are intentional combinations of trees with crops and/or livestock which involve intensive management of the interactions between the components as an integrated agroecosystem.
Intentional: Combinations of trees, crops and/or animals are intentionally designed and managed as a whole unit, rather than as individual elements which may occur in close proximity but are controlled separately.
Intensive: Agroforestry practices are intensively managed to maintain their productive and protective functions, and often involve annual operations such as cultivation, fertilization and irrigation.
Interactive: Agroforestry management seeks to actively manipulate the biological and physical interactions between the tree, crop and animal components. The goal is to enhance the production of more than one harvestable component at a time, while also providing conservation benefits such as non-point source water pollution control or wildlife habitat.
Integrated: The tree, crop and/or animal components are structurally and functionally combined into a single, integrated management unit. Integration may be horizontal or vertical, and above- or below-ground. Such integration utilizes more of the productive capacity of the land and helps to balance economic production with resource conservation.
Liebig 2008. Measured to a depth of 120 cm (~4 ft), switchgrass grown for bioenergy at 10 farms across the Great Plains in the United States sequestered 2.9 tC/ha/yr (1.16 tC/ac/yr). Of that, only 1.1 tC/ha/yr (0.44 tC/ac/yr) was found in the first 30 cm (1 ft) depth, with the remainder measured below 30 cm. The authors explain what makes switchgrass effective in carbon sequestration:
Increases in SOC [soil organic carbon] under switchgrass were likely caused by belowground C input from root biomass and rhizodeposition and decreased soil organic matter losses by erosion. Research conducted by ecologist John Weaver and his graduate students over 60 years ago provide ancillary support for increased SOC under switchgrass. Their detailed surveys of prairie grass roots indicated switchgrass to have the deepest root system of all grasses examined, with roots extending to a soil depth of 3m (~10 ft). This finding, coupled with observations that prairie grass roots regenerate by replacing dying roots with new, live roots indicates the potential for significant C input to the soil under switchgrass.
Montagnini & Nair 2004. Agroforestry systems are multifunctional with respect to carbon capture. Agroforestry can: increase the soil carbon content and fertility of cropland, while allowing for continued food production; create greater sequestration efficiency through diversity of vegetation; and allow for harvest of forest products, potentially keeping carbon sequestered in wood products for many years, and thereby also decreasing pressure on natural forests. And because of the mixed use of agroforestry systems:
[T]he amount of biomass and therefore carbon that is harvested and ‘exported’ from the system is relatively low in relation to the total productivity of the tree (as in the case of shaded perennial systems). Therefore, unlike in tree plantations and other monocultural systems, agroforestry seems to have a unique advantage in terms of C sequestration [Montanigni & Nair 2004: 285].
A few sequestration rates highlighted in this article include: A Costa Rica study of cacao grown under two different species of shade trees Erythrina (a leguminous tree) and Cordia (a timber tree), measured C sequestration in perennial plant biomass at an average of 4.28t C/ha/yr (1.7t C/ac/yr) for the cacao-Cordia system, and 3.08t C/ha/yr (1.2 tC/ac/yr) in the cacao-Erythrina system . In another study, tropical smallholder agroforestry was projected to sequester 1.5-3.5t C/ha/yr (0.6-1.4 tC/ac/yr).
Onim 1990. Tropical agroforestry was observed to increase SOC (soil organic carbon), at the 0-30 cm depth, to a maximum of 8.34 tC/ha/yr (3.38 tC/ac/yr)and minimum of 0.73 tC/ha/yr (0.30 tC/ac/yr).
Biochar
Biocharis organic matter that has been decomposed through pyrolysis (burning) under controlled, low-oxygen conditions, where it emits relatively little CO2. Biochar is then added to the soil for long-term carbon storage and/or enhancing availability of soil nutrients, oxygen and water to plants and microbes. Because charred biomass has been observed to persist in the soil for centuries or millennia, biochar is seen as a stable or recalcitrant form of carbon that that may prove to be a useful tool for reversing climate change. Not only is the biochar itself a stable form of carbon that can remain in soils long-term, but also it helps build healthy soil structure which increases plant growth and therefore photosynthetic capacity, resulting in carbon being removed from the atmosphere and stored in biomass or soils. [McLaughlin 2017; Taylor 2010; Paustian 2016; Weng 2017; Remediation Magazine 2017]
It is worth noting that depending on the pyrolysis technique, the resulting biochar may range in quality from poor to excellent. One hopes that as the industry matures, the understanding of the importance of biochar quality in assessing results will grow as well.
McLaughlin 2017. Hugh McLaughlin, Ph.D., P.E. is an expert on the properties and production of chars created by pyrolyzing biomass, and the subsequent conversion to activated carbons. He has published extensively on biochar and biomass-derived heat production. In this video he gives a short but comprehensive review of the qualities and use of biochar.
Paustian 2016. Biochar application to soils is considered in this article among several activities (such as compost application, cover cropping, residue retention, no-till, and others, as previously mentioned in this compendium) designed to increase soil C stocks by increasing organic matter inputs or reducing decomposition rates. Biochar acts as a soil amendment stimulating plant growth, thereby allowing for greater C storage through greater biomass production, while also embodying a generally stable form of buried carbon.
Biochar mineralizes 10–100 times more slowly than uncharred biomass. Thus a large fraction of added C … can be retained in the soil over several decades or longer, although residence times vary depending on the amendment type, nutrient content and soil conditions (such as moisture, temperature and texture).
However, because the organic matter originates from outside the ecosystem ‘boundary’, a broader life-cycle assessment approach is needed, that considers the GHG impacts of: (1) offsite biomass removal, transport, and processing; (2) alternative end uses of the biomass; (3) interactions with other soil GHG-producing processes; and (4) synergies between these soil amendments and the fixation and retention of in situ plant-derived C. In many cases, net life-cycle emissions will largely depend on whether the biomass used as a soil amendment would have otherwise been burnt (either for fuel, thereby offsetting fossil fuel use, or as waste disposal), added to a landfill, or left in place as living biomass or detritus [Paustian 2016: 50].
Remediation Magazine 2017. A popular report on Weng 2017, quoting the authors:
The project’s leader, DPI [Department of Primary Industries] researcher and SCU [Southern Cross University] adjunct professor Lukas Van Zwieten said the research threw up some unexpected results. “We immediately saw an increase in soil carbon from the biochar, as expected, but what we didn’t expect was that soil carbon content continued to increase. This research demonstrates the ongoing benefits of biochar in farming systems to improve pastures and grasslands and increase farmers’ production and profitability . . . the researchers found that biochar enhanced the below-ground recovery of new root-derived carbon by 20% – that is, more of the carbon photosynthesised by plants was retained in the biochar-amended soil. Biochar accelerated the formation of soil microaggregates via interactions between organic matter and soil minerals, thus stabilising the root-derived carbon. . . . The increased microbial activity and improved physical structure of the soil would also ultimately improve the effectiveness of fertiliser use, making the application of biochar particularly beneficial for high-end, intensive crop production”
“[T]he improved structure of the soil protected the naturally occurring carbon, as well as the carbon added”, said Southern Cross University’s associate professor Terry Rose, a co-author of the study. “Importantly, the biochar also slowed down the natural breakdown of native soil organic carbon by more than 5%.
Taylor 2010. An anthology of articles written by biochar pioneers. Covers biochar history, testing, production, challenges and uses. Suitable reading for general audiences as well as land management and industry professionals.
Weng 2017. Biochar can increase the stable C content of soil. However, studies on the longer-term role of plant–soil–biochar interactions and the consequent changes to native soil organic carbon (SOC) are lacking. . . . We found that biochar accelerates the formation of microaggregates via organo-mineral interactions, resulting in the stabilization and accumulation of SOC in a rhodic ferralsol (s.a. Remediation Magazine 2017).
Note: As mentioned in the Release notes, we have a small staff and therefore have had to postpone some important material to the next release, scheduled for January 2018. This will include a more thorough exploration of the importance of wetlands in addressing climate.
Wetlands only cover only a small proportion of the terrestrial surface area, with estimates ranging from 5-8% [Mitsch 2007]. Despite this, they store a disproportionate amount of Earth’s soil carbon, with estimates ranging from 20-30% [Lal 2008]. Soil carbon in wetlands can be as high as 40% [Vepraskas and Craft 2016; cf. Nahlik 2016: 2], compared to typical estimates for agricultural soils that range up to 2% [Lal 1995; see the Croplands section of this Compendium for a detailed discussion of agricultural soil carbon].High carbon storage in wetlands is the result of anoxic conditions in wetland soils that lead to slowed decomposition and a resulting increase in stored organic matter. Wetlands have significant potential to act as carbon sinks under the right circumstances. Potentially reducing the carbon sequestration potential for wetlands are the methane emissions that they produce. Notwithstanding the uncertainty of methane emissions, wetlands globally likely serve as a significant net carbon sink. With the complexity of processes that contribute to wetland carbon exchange and resulting function as GHG sinks or sources, management decisions may be a critical factor in determining the carbon sequestration outcome of wetland ecosystems. [See Nahlik 2016 for review of quantitative data]
Overview
The importance of wetlands cannot be overstated. They exist in all biomes ranging from the tropics to the tundra, and on every continent except Antarctica . Wetlands also tend to host much higher concentrations of biological diversity than other ecosystems. The fact that the oldest intergovernmental environmental convention, the Ramsar Convention of 1971, was focused explicitly on wetland conservation is a testament to this importance [Mitra 2005].
The comparison of carbon dioxide storage to methane emissions is a source of uncertainty in determining the carbon sequestration potential for wetlands. Given methane’s stronger radiative forcing as a greenhouse gas, the IPCC currently uses a ratio of 25:1 to indicate the methane to carbon global warming potential. Despite the high global warming potential presented by methane, it has a relatively short atmospheric “lifetime”, often reported in the range of 8-12 years. Although methanotrophic (methane metabolizing) bacteria are not typically calculated into the equations regarding the atmospheric lifetime of methane, bacteria that metabolize atmospheric methane (e.g., high-affinity oxidation methanotrophs) provide an additional mechanism hastening methane attenuation [Jardine 2009].
Several trends appear to be instructive in wetland carbon storage. Tropical and temperate wetlands generally hold more carbon than northern boreal peatlands [Mitsch 2012: 7-9]. Freshwater inland wetlands may hold more carbon than coastal saline wetlands, though this may be based primarily on their surface area extent. More carbon storage occurs at depths from 30 – 120 cm (1-4 ft), at least as reported by an analysis of U.S. conterminous wetlands in which 65% of the total carbon was deeper than 30 cm (1 ft) [Nahlik 2016: 2-3]. A correlation exists between increased anthropogenic disturbance and decreased carbon storage, although this may be an artefact of settlement patterns rather than a causal relationship [Nahlik 2016: 4]. One study reported high carbon sequestration and low methane emissions in constructed wetlands [Mitsch 2012]; this is an encouraging result considering the many wetlands created as mitigations to compensate for human development impacts.
Wetland Article Summaries
Apfelbaum 1993. Steve Apfelbaum of Applied Ecological Services, Inc. is a restoration ecologist with several decades of experience around the world. This brief paper, “The Role of Landscapes in Stormwater Management,” describes the historical condition of wetlands in the upper midwest, the degrading effects of agriculture and urbanization on water cycles, vegetation and the resultant pollution. Included are recommendations for restoration of healthy wetlands and methods for slowing the movement of water so that it may keep soils hydrated and feed local ecosystems. In addition, restoration of wetlands includes high-capacity carbon storage due to low-oxygen conditions in wetland soils, with subsequent low rates of oxidation and loss of carbon to the atmosphere.
This paper presents evidence that many existing streams did not have conspicuous
channels and were not identified during pre-settlement times (prior to 1830s in the Midwestern United States). Many currently identified first, second, and third-order streams were identified as vegetated swales, wetlands, wet prairies, and swamps in the original land survey records of the U.S. General Land Office.
The data presented show that significant increases in discharge for low, medium, and high flows have occurred since settlement. Stream channels have formed inadvertently or were created to drain land for development and agricultural land uses. Currently, discharges may be 200 to 400 times greater than historical levels, based on data from 1886 to the present for the Des Plaines River in Illinois, a 620-square-mile watershed. Historic data document how this river had no measurable discharge or very low flow conditions for over 60 percent of each year during the period from 1886 to 1904.
This study suggests that land-use changes in the previous upland/prairie watershed have resulted in a change from a diffuse and slow overland flow to increased runoff, concentrated flows, and significantly reduced lag time. Preliminary modeling suggests the following results: reduced infiltration, reduced evaporation and evapotranspiration, greatly increased runoff and hydraulic volatility, and increased sediment yields and instream water quality problems caused by destabilization of streambanks.
The opportunity to emulate historical stormwater behavior by integrating upland landscape features in urban developments and agricultural lands offers stormwater management options that are easier to maintain, less expensive over time, attractive, and possibly more efficient compared with many conventional stormwater management solutions and the use of biofiltration wetlands.
and
Diverse and productive prairies, wetlands, savannas, and other ecological systems occupied hundreds of millions of acres in presettlement North America. These ecological systems have been replaced by a vast acreage of tilled and developed lands. Land-use changes have modified the capability of the upland systems and small depressional wetlands in the uplands to retain water and assimilate nutrients and other materials that now flow from the land into aquatic systems, streams, and wetlands. The historical plant communities that were dominated by deep-rooted, long-lived, and productive species have been primarily replaced by annual species (corn, soybeans, wheat) or shallow rooted non-native species (bluegrass lawns, brome grass fields). The native vegetation was efficient at using water and nutrients, and consequently maintained very high levels of carbon fixation and primary productivity. Modern communities, in turn, are productive but primarily above ground, in contrast to the prairie ecosystem where perhaps 70 percent of the biomass was actually created below ground in highly developed root systems. These changes in the landscape and vegetation coupled with intentional stormwater management have changed the lag time for water to remain in uplands and consequently increased the rate and volume of water leaving the landscape.
Mitsch et al 2012. This study evaluated the carbon storage and methane exchange potential for seven wetlands based on field data collected over several years, and used field data collected at 14 other wetlands globally to model the carbon sequestration and methane emission potential out to 300 years. A total of 21 wetlands were examined. The modeling accounted for the anticipated half-life of methane oxidative degradation in the atmosphere. Results indicated that methane emissions become unimportant within the 300 year model simulation time range, with most wetlands making the shift to net carbon storage by year 100 of the model. The study supports the potential for wetlands as carbon sinks.
Nahlik and Fennessy 2016. The objective of this article was to quantify the carbon stocks present in wetlands of the conterminous United States. To do so, the authors examined empirical field data collected during the 2011 National Wetland Condition Assessment conducted by the U.S. Environmental Protection Agency, and used this to quantitatively extrapolate to larger scale carbon estimates. These estimates were developed at regional and national scales. Results were evaluated by region, wetland type, freshwater or tidal status, and level of anthropogenic disturbance. The article indicated that 11.52 gigatons of carbon are present in the U.S., much of which is in soils deeper than 30 cm (1 ft). Freshwater wetlands located inland held nearly ten-fold as much carbon as intertidal wetlands overall, although this is at least partly due to the much greater aerial extent of inland freshwater wetlands; tidal wetlands still had higher concentrations of carbon storage. The authors also indicate a possible relationship between anthropogenic disturbance and carbon stocks, wherein less disturbed sites store more carbon. Insufficient data was available to determine whether this was a causal effect or an artefact of some kind, such as human preference in settlement patterns. The authors conclude that, due to the substantial carbon stocks that wetlands represent and the potential for anthropogenic impacts, existing intact wetlands should be protected to avoid the risk of further contributing to climate change.
With the rise of civilizations, humans began having significant impacts on bodies of water and the water cycle. The early “hydraulic civilizations” appeared along major rivers (Nile, Tigris-Euphrates, Indus, Yellow River and others), changed watercourses and built canals for agriculture and transportation. As populations and cities expanded, demand for food led to soil depletion while the built environment created growing areas of impermeable surfaces. Disruption of water cycles has reached a peak since the industrial revolution, with large areas of land covered with impermeable surfaces, and rainwater and waste rapidly shuttled away from land into the oceans. [Kravčik 2007: 42 ff.]
Eco-restoration requires a shift in thinking about water management and fortunately there are many successful water restoration projects under way, along with a strong theoretical basis to guide them. Water management is the key to cooling the biosphere, regenerating photosynthesis and drawing down carbon on billions of acres. Included in this section is a brief discussion of wetlands, which will be expanded in the next Compendium release.
Several eco-restoration examples are included here, and many were explored at our 2015 Restoring Water Cycles to Reverse Global Warming Conference at Tufts University – all speaker videos are available online. https://bio4climate.org/program-tufts-2015/.
Overview
Healthy Soils Australia, Walter Jehne 2015. Walter Jehne is a microbiologist, soil and climate scientist who has spent the past several years teaching and promoting the use of nature’s tools to address destruction of land and water cycles, and educating on methods important in addressing global warming. The text below represents excerpts (condensed and edited) from the paper, “Restoring Regional Rainfalls: Background Brief for Outcomes, Australia Forum on Vegetation-Rainfall Relationships”:
Contrary to the dominant assumptions that global warming is due to elevated atmospheric carbon concentrations,
Systemic climate changes such as aridification began in the 1970s well before CO2 levels or its temperature effects increased abnormally.
Water-based processes govern most climate effects and over 95% of the earth’s heat dynamics for billions of years, including some 80% of the natural greenhouse effect.
These hydrological heat dynamics have been ignored or deemed “secondary feedbacks” to the CO2 greenhouse effect because water is assumed to be a dominant determinant of our climate, and humans could not possibly have altered these global cycles to cause the anthropogenic climate changes
The hydrological processes are highly complex and difficult to model, and were therefore excluded in IPCC (Intergovernmental Panel on Climate Change) assumptions and models, whereas the minor CO2 component of the greenhouse effect is more readily modelled, and provides a simple marketable explanation of its “cause.”
Because of these IPCC assumptions, policy and response options have largely ignored the dominant hydrological determinants on climate, the effects of land management, and impacts these changes may have on climate, water and bio-system stability.
Yet we have greatly altered the earth’s natural hydrology and thus heat dynamics by:
Clearing over 75% (6.3 billion hectares or 15.75 billion acres) of the earth’s primary forest, greatly altering the land’s albedo and heat reflectance as well as transpiration and latent heat fluxes that cooled vast regions.
Oxidizing and eroding organic matter from some 10 billion hectares of soils thereby reducing the ability of landscapes to infiltrate, retain and supply water to sustain cooling transpiration and latent heat fluxes and the drawdown of carbon from the air by plants via photosynthesis.
Exposing vast areas of such degraded, cropped and bare soils to erosion which has dispersed 3 billion tonnes of additional dust aerosols into the air where it nucleates warming humid hazes that retain heat in the biosphere.
Heating bare exposed soils to greatly increase their re-radiation of heat which massively increases greenhouse warming effects.
Increasing the absorption of solar radiation by humid haze micro-droplets [resulting in] global dimming (while in the liquid phase), as well as the absorption of re-radiated heat (while in the gaseous phase) to warm the lower atmosphere via the water vapour greenhouse effect.
Reducing regional rainfalls often by up to 30% due to the increase in persistent haze micro-droplets which are too small to coalesce into raindrops and precipitate by themselves.
Increasing surface humidity due to the persistent humid hazes, thus lowering evaporation rates by up to 10% and reducing latent heat fluxes which transfer heat out of the biosphere into space.
Reducing the production of the biological precipitation nuclei from forests that would help coalesce the humid haze micro-droplets to form dense clouds with high albedos that reflect 33% of solar radiation out to space, thereby regulating global temperatures.
Preventing the nucleation of haze and cloud droplets into raindrops which lowers rainfalls and the level and longevity of transpiration, photosynthesis and cooling latent heat fluxes.
Impairing the night-time escape of re-radiated heat to space via natural “radiation windows” due to the impaired nucleation and precipitation of such “blocking” humid micro-drop hazes.
Increasing sustained high pressures above the cleared, bare heated soils that prevents the inflow of cool moist air from oceans, its precipitation and the associated cooling heat fluxes.
Extending such high pressure over vast regions and periods to accentuate the aridification of bio-systems which readily collapse to deserts with further human land degradation. [Healthy Soils Australia 2015: 1-2]
Given this reality we need solutions that go beyond just reducing future CO2 emissions but also:
Cool regions and the climate so as to offset dangerous warming and its feedback effects.
Draw down carbon back into its safe soil sinks so as to reduce its greenhouse effect.
Restore the resilience of agro-ecosystems and communities to the extremes and secure their essential water, food and bio-material needs via just, safe low carbon futures. [Healthy Soils Australia 2015: 8]
Regenerate natural hydrological processes by land management which captures water in soils, wetlands, aquifers and biomass
Maintain healthy biodiverse soils to Restore microbial drivers that govern these cooling hydrological processes by emitting condensation nuclei that lead to rainfall.
Support the biological sequestration of carbon from the air into stable soil humates and glomalin to enhance the water held in the soil reservoirs that sustain the cooling latent heat fluxes.
Support the production of microbial precipitation nuclei that coalesce the warming humid hazes into dense high albedo clouds that cool regions and generate critical cooling rainfalls.
Promote the nucleation and enhancement of rainfall in key regions to maintain the latent heat fluxes, green vegetated habitats and the radiation windows that enhance nighttime cooling effects.
Only by regenerating our forests, soils and landscapes can we now restore the hydrological cooling processes that helped govern the natural heat dynamics and buffered climate of the blue planet. Such regeneration is now our only option to offset the dangerous climate feedbacks resulting from the warming induced by our landscape degradation and its associated abnormal rise in CO2 levels.
Fortunately viable practical options exist to enable us to do and directly benefit from this, at grass roots community level: tree by tree, hectare by hectare, region by region. While we face a global emergency and must all take responsibility for it, it can only be addressed locally via practical action on the ground by communities driven by their own self interest in securing a safe climate and future.
The good news is we can still avoid the pending extremes and collapse provided we focus on direct local action urgently to regenerate the health of each square metre of soil and each forest and tree. We have the abundant degraded land, sunshine, CO2, waste biomass and nutrients to do it with. To grow more green areas; by regenerating our soils, forests, rangelands and even re-greening deserts.
We can do this if we enhance the infiltration, retention and availability of each raindrop in our soils so that the regenerated ‘in soil reservoirs’ sustain healthy green growth over larger areas for longer. This will happen naturally, synergistically, as plant growth enhances the structure of the soil by increasing its carbon content which in turn aids its water holding capacity and nutrient dynamics.
Just as nature did over the past 420 million years in colonizing and greening the earth’s land surface, these same processes are now our only option to regenerate our soils, forests and landscape and re-secure our safe climate and future. [Healthy Soils Australia 2015: 7-11]
Water Article Summaries
Ellison 2017. “Trees, forests and water: Cool insights for a hot world” may be one of the few articles in the mainstream literature relating to climate that puts hydrological cooling effects before carbon in importance for addressing global warming, although dynamics of water and carbon are closely intertwined.
Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. [Ellison 2017: 51]
This paper is discussed further under Forests.
Evans, Griggs 2015. Carol Evans is a fisheries biologist and Jon Griggs is a rancher in northeastern Nevada. They have worked together over twenty-five years to restore overgrazed lands to health through planned grazing of cattle, which also brings water, trout, beavers and biodiversity to the riparian areas of Maggie and Susie Creeks. In the driest state in the U.S., with less than ten inches of rain a year, they now have perennial streams and wet meadows after five years of the worst drought in memory.
Susie Creek, ca. 1989 (left) and 2015 (right) after five years of drought. Elko, Nevada
Kravčík 2007. Michal Kravčík and co-authors are Slovakian hydrologists who have developed what they call a new water paradigm for managing water cycles, floods and drought.
In a healthy water cycle, while some rain enters streams and rivers directly and is carried off to sea, most rain water is absorbed by the soils in situ, where it lands. The rain gives life to the soil and sets many biological processes in motion, where it is essential for stable soil carbon storage and cooling the biosphere. This includes evapotranspiration from plants which returns water as vapor to the atmosphere where the water condenses and falls as rain. The cycle then begins anew. Kravčík et al. call this the “small water cycle” (i.e., local water cycle) where most water goes through its cycles in a regional area or smaller. The “large water cycle” is the exchange of water between oceans and land, and “above land water circulates at the same time in many small water cycles which are subsidized with water from the large [continental or global] water cycle.” [Kravčík 2007: 16]
Civilizations disturb healthy water cycles and accelerate the runoff from land by creating impermeable surfaces (including degraded farmlands and rangelands), and preventing water from remaining in place to sink into soils or to forcing it to run off the land, causing floods and often carrying valuable topsoil with it. Furthermore, water systems have been engineered to move water away from its source to the oceans. Water, with its growing use as a means to dispose of farming, industrial and human wastes, is even seen as a nuisance. As a result, less water returns to continents from the oceans than is lost from continents to oceans, which leads to desiccation of soils, severe drought, wildfires, desertification, and a measure of sea-level rise. There is a growing understanding that these phenomena, often attributed to climate change, may in fact also be a function of disrupted water cycles.
Restored urban land, Kosice. November 2005 (left), September 2006 (right).
Heat from the sun drives these earthly water cycles. Small water cycles are local, circulating water within a relatively small area. Latent heat causes water to evaporate; heat is absorbed in the process of evaporating water and does not result in an increase in local temperature. We thus do not experience latent heat as an increase in temperature. However, when there is less water available for evaporation, less solar energy is transformed into latent heat and more solar energy is transformed into sensible heat, heat you can feel as increased temperature. This is the heat that we are increasingly experiencing as global warming.
A great deal of heat is moved from the surface of the earth to the upper atmosphere by evaporation and transpiration of water by plants, contributing to significant cooling of the biosphere – to illustrate it takes 540 calories to turn 1 gram of water to vapor; by comparison it takes only 80 calories to melt 1 gram of ice. Draining of land, that is, runoff and floods, can be reversed through comprehensive conservation of rainwater which maintains the sponge-like absorption capacity of soils and maintains many aspects of soil health, resilience, biodiversity and productivity. Renewal of small water cycles over land can temper extreme weather events and ensure a growth in water reserves by eliminating heat islands and problematic distribution of atmospheric moisture.
Nobre 2010. Antonio Nobre is an Amazon scientist who has studied the biotic pump (see also Makarieva), and tells how he was once told by an indigenous wise man,
“Doesn’t the white man know that, if he destroys the forest, there will be no more rain? And that if there’s no more rain, there will be nothing to drink, or to eat?” I heard that . . . [ and thought], “Oh, my! I’ve been studying this for 20 years, with a super computer; dozens, thousands of scientists, and we are starting to get to this conclusion, which he already knows!” A critical point is the Yanomami have never deforested. How could they know the rain would end? This bugged me and I was befuddled. How could he know that?
Some months later, I met him at another event and said, “Davi, how did you know that if the forest was destroyed, there’d be no more rain?” He replied: “The spirit of the forest told us.”
The equatorial region, in general, and the Amazon specifically, is extremely important for the world’s climate. It’s a powerful engine for evaporation. From a satellite viewpoint, atmospheric water flow can look like a geyser, which is underground water heated by magma transferred into the atmosphere. There are no geysers in the Amazon but trees play the same role. They, like geysers, transfer an enormous amount of water from the ground into the atmosphere. Nobre continues:
There are 600 billion trees in the Amazon forest, 600 billion geysers. That is done with an extraordinary sophistication. They don’t need the heat of magma. They use sunlight to do this process. On a typical sunny day in the Amazon a large tree manages to transpire 1,000 liters of water. If we take all of the Amazon, which is a very large area, and add up all the water that is released by transpiration, “the sweat of the forest,” an incredible amount of water is evaporated into the atmosphere: 20 billion metric tons of water per day. . . . This river of vapor that comes up from the forest and goes into the atmosphere is greater than the Amazon River.”
The Amazon River itself is the largest river on Earth, it carries one fifth of all the fresh water, it releases 17 billion metric tons of water a day into the Atlantic Ocean, smaller than “the river in the sky.” To evaporate the 20 billion tons of water released by trees it would take 50,000 of the largest hydroelectric plant in the world, Itaipus, which generates 14 GW of electricity, 30% of Brazil’s power. The Amazon does this with no technology, at no cost.
Schwartz 2016. Judith Schwartz once again travels the world to collect stories of lands restored, of lives revived, this time to glean insight from restorers of water. She demonstrates that many of our assumptions about managing water are derived from engineering, not biology. When biology is the focus of the water and rainfall question the problem is redefined, and clarified. Solutions that had been invisible become apparent, and provide the opportunity for far more effective responses – even in some of the driest places on earth. Floods and droughts become manageable, even preventable entirely. Two of the innovators mentioned in Water in Plain Sight, Michal Kravčík and Rajendra Singh, spoke at Biodiversity for a Livable Climate’s 2015 Restoring Water Cycles conference, as did Judy.
Singh 2007. Rajendra Singh, the “Water Man of India,” tells the story of how he helped over 1,000 villages restore water and abundance through the use of ancient, low-technology land management. Providing water for people, farms and animals, such efforts countered the ill effects of industrialization and reversed flight to the cities. Says Singh:
I am neither a scientist, nor a professional water engineer nor a climate change expert. I am a small constructive worker of Gandhi and I mobilize the civil society and the community for action on natural resources management and conservation for rural uplift in India. Here I am recording the impact of the above work on the ecology of 6,500 square km area in Alwar district from 1985‐2007. Since 1985, 8,600 small water harvesting talabs [a form of check dam] in 1,068 villages of Alwar district covering 6,500 square km area have been built. This has resulted in the shallow aquifer recharge in groundwater bringing up the water table from about 100‐120 meters depth to 3‐13 meters at present. The area under single cropping increased from 11 per cent to 70 per cent out of which area under double cropping increased from 3 per cent to 50 per cent bringing prosperity to the farmers. The forest cover, which used to be around 7 per cent increased to 40 per cent through agro‐forestry and social forestry, providing sufficient fuel wood and sequestering carbon from atmosphere [Singh 2007: 5].
A dramatic example of how large restoration efforts are built from small, local efforts. In the ten years since this paper, Tarun Bhagat Sangh has continued to expand its work.
Although often overlooked, ignored or taken for granted, earthworms are nevertheless keystone soil species, mediators and moderators for rebuilding healthy, biodiverse, high carbon and moisture rich topsoil [Darwin 1881; Blakemore 2016c]. We depend on soils for more than 99% of our food and 100% of our timber and natural fibres [Blakemore 2012, Pimentel 2013]. As an integral part of organic production, earthworms are key to agricultural sustainability and global ecosystem stability. Ancient in origin (probably pre-Cambrian but certainly more than 500 million years old), the 7,000 known species of earthworms are ubiquitous and invariably associated with topsoil humus. Earthworms are a basis of terrestrial food webs and the ultimate detritivor [Blakemore 2016c], recently reinstated as key players in the International “4 per 1000 Initiative” [4p1000.org, n.d.] to increase soil organic matter to store carbon. In this section, we discuss the abundance and variety of earthworms and their role in soil health and functionality.
Overview
Extrapolating data from Darwin [1881], their population numbers around 1.3 x 1015 or 1.3 quadrillion globally with biomass of 0.4 t/ha x 9.5 Gha of productive land = 3.8 Gt. This is about ten times the biomass of all humanity, and twice that of both all domesticated stock and total global fish [Blakemore, 2017]. Forming possibly the largest beneficial animal resource on the planet, earthworms are yet apparently severely depleted by cultivation and agrichemical excesses of industrial farming, often being absent from such soils [e.g. Lee 1985] with both their populations and biodiversity in decline [Blakemore 2016a, b, c].
In comparison to intensive agrichemical farming, studies by Blakemore, [2000, 2016a,b] show a diverse array of up to 23 earthworm species per organic farm site (mean 13 spp), implicated in 16-80% increased crop or pasture yield (mean +39%) plus an average of 12% extra soil moisture storage (range 7-91%) compared to conventional neighbour farms. Carbon sequestration is restored at rates two to three times higher in pasture. Such findings are highly relevant due to looming species extinction and climate change with requirement to meet the needs of a growing population. Organic farming can thus produce higher yields and sequester more carbon.
Earthworms may number up to 1,000~2,000/m2 (10-20 million/ha, or 4-8 million/ac) in fertile soils with biomass as high as 3-5 t/ha, (1.2-2 t/ac ) so earthworm stocks may outweigh the above ground stock [Lee 1985; Blakemore 2016c, 2017]. Earthworm abundance and diversity increase in a truly sustainable system as they convert all organic ‘wastes’ into humus-rich compost while processing all atmospheric CO2 in 12 yr cycles [Blakemore 2016a]. Their burrows, as long as 9,000 km/ha (2250 mi/ac) [Kretzchmar 1982] and up to 15 m in depth (49.2 ft) [Sims & Gerard, 1999: 27, as cited in Blakemore 2016c] aerate, improve water infiltration and, importantly, provide habitats for many other beneficial organisms and microbes that they help distribute throughout the entire soil profile. All rainfall is filtered through their burrows and water is stored in worm-worked humus. Blakemore [2000] found up to 90% extra water in pasture compared to adjacent arable fields, and organic arable soil stored 40% more water than chemically farmed arable soil [Blakemore, Hochkirch 2017].
Wormless soils are economically and ecologically expensive: they need to be plowed regularly, and require extra irrigation plus subsidized artificial chemical nitrogen fertilizers and biocide sprays to fight off plant infections and infestations [Howard 1945; Balfour 1975]. This toxic burden has severe impact upon non-target organisms and any organism fed the crops – including humans – as well as poisoning the soil, air, waterways and oceans. Such findings are summarized in Lady Eve Balfour’s IFOAM presentation in 1977 [Balfour 1977]. Another compelling reason for earthworm conservation is that it is impossible to “geoengineer” by addressing isolated variables the many benefits and essential irreducible systems services that earthworms freely and relentlessly provide. In other words, we have no viable alternative to earthworms.
Soil and Earthworm Relationships
We face a complexity of inter-relating ecological problems.Intensive chemical agriculture is a major GHG contributor (28-50%) and a major source of extraneous CO2 (currently 10-25% and in total historically up to 40%) [Houghton 2010: 338, 348]:
Globally, the conversion of lands to croplands has been responsible for the largest emissions of carbon from land-use change. . . From 1850 to 2000, land use and land-use change released an estimated 108–188 Gt (billion tons) of carbon to the atmosphere, or about 28–40% of total anthropogenic emissions of carbon (274 Gt C from fossil fuels) [Strassmann 2008].
The FAO [Gerber 2013] found that intensive industrial livestock farming (rather than organic husbandry) contributed 14.5% of human-induced GHG emissions. A newspaper report [Bryce 2013] comments:
The FAO’s last livestock report, a 2006 assessment titled Livestock’s Long Shadow, found that farms breeding chickens, pigs, and cows for meat and dairy products, produced a disconcerting 18% of global greenhouse gas emissions . . . Around 30% of global biodiversity loss can be attributed to livestock production, such as the spread of pasture land or turning over forests and savannahs.
Although these figures vary due to different formulas for budgeting, it’s clear that agriculture in all its forms, including the practice of forest clearance, is a major contributor to GHG emissions.
The traditional, innovative & scientific methods of non-chemical, organic farming and Permaculture appreciate the importance of earthworm conservation [Howard 1945; Balfour 1975; Mollison 1988]. As a key player in natural processes and crucial issues, Darwin’s “lowly earthworm”, although neglected, warrants re-ascendency to its former position as premier farm livestock [Howard 1945]. For our own health and for that of our planet, we urgently need wholly natural vermi-composting at all scales (from kitchen to continent) in order to replace synthetic fertilizers and to facilitate rapid transition to broad-acre organics that also has earthworm livestock at its core. Enabling earthworms to restore healthy soils is vital to stabilizing climate. All organic ‘wastes’ and manures should be recycled via vermi-composting and appropriate management employed to enhance populations of field-working worms.
Earthworm Article Summaries
van Groenigen et al. 2014.In a recent meta-analysis, while not considering organic farming or carbon per se, this study confirmed earthworm presence corresponding to crop yield increases of 25%, which is comparable to average ~39% extra organic yield in soils with earthworm proliferations determined by Blakemore [2000, 2016b]. This supports earlier studies by Wollny [1890: Forschungen auf der Gebiet der Agrikultur-Physik, 13, s. 381] that found addition of earthworms to soil led to a marked increase of cereal grain by 35-50% and of straw by 40%.
Solomon 2013.
Although earthworms are beneficial in gardens and agricultural fields, they are harmful to Michigan’s forests where they are an invasive species. . . . Earthworms are not native to Michigan and the Great Lakes region, at least not since before glaciers covered the region; they were brought here during European settlement in the 1800s or possibly earlier. Plants, wildlife and forests evolved without any of these creatures around. They are now an invasive species that harms forests.
Hardwood forests without earthworms have a thick layer of slowly decomposing leaves, or “duff” that promotes a rich community of wildflowers, tree seedlings and small animals. Earthworms change that environment dramatically by essentially consuming the duff, thereby destroying habitat and reducing fertility. In contrast to their effect in gardens, earthworms cause forest soils to become more compacted. As a result of habitat loss, fertility declines and soil compaction, these forests may be less productive and have poorer new tree regeneration in the long run.
Another view, from oligochaetologist (worm scientist) Rob Blakemore, is as follows:
Regarding popularized concerns about alien Asian invasive worms threatening to destroy American native forests, this may reasonably be regarded as part of a process that is commonly known as Ecological Succession [Odum 2005].
Ironically, the ecological concept of succession started with Thoreau and Cowles on studies of forest succession and on the Lake Michigan dunes. Large parts of the northeastern North America were glaciated up to about 10,000 years ago completely destroying all land surfaces and forming the Great Lakes. When the ice retreated Nature returned in successive waves and, gradually, the soil, vegetation, and animals communities re-established and species continue to evolve.
According to Darwin [1881] earthworms are supremely important for natural productivity and for the recent progress of human civilizations. In this context the woodlands of Michigan seem a relatively minor issue compared to species extinction and climate change. Healthy soils generally harbour earthworms and it appears there had been insufficient time for these slow-moving and flightless organisms to colonize without fast-track via incidental intervention of most-recent human settlers, often as anglers on the Great Lakes.
When exotic crops and stock were introduced around the world 10,000 years ago, so too were attendent earthworms and these have now spread to “pristine,” albeit transitional, woodlands. The many benefits earthworms have for agricultural and unmanaged soils may cause some changes in more natural habitats but this is a virtually unavoidable and irreversible force majeure and fact-of-life.
Certainly there will be a new ecological balance in time, possibly at a different level of productivity and biodiversity. That is the definition of succession.
Héry et al. 2008. Earthworms have been observed to increase methanotrophy (methane metabolic breakdown) in soil covering a landfill; this is most likely “due to the stimulation of bacterial growth or activity than to substantial shifts in the methanotroph community structure” [Hery 2008: 92].
Earthworm-mediated bioturbation has been linked to an increase in methanotrophy in a landfill biocover soil (AC Singer et al., unpublished), but the mechanism of this trophic interaction remains unclear. The aims of this study were to determine the composition of the active methanotroph community and to investigate the interactions between earthworms and bacteria in this landfill biocover soil where the methane oxidation activity was significantly increased by the earthworms [Hery 2008: 92].
And
We proposed the hypothesis that earthworms could stimulate the growth or the activity of methanotrophs. We showed that the earthworm-mediated increase of methane oxidation in the landfill biocover soil only weakly correlated with a shift in the structure of the active methanotroph population. Future work needs to focus on the relationship between this earthworm effect on enhanced methane oxidation in landfill cover soil and this effect on bacterial activity and growth. The possible contribution of an enriched population of nitrifying bacteria to methane oxidation also requires further investigation [Hery 2008: 101].
This compendium is, if nothing else, a testament to the key role soils must play if we are to preserve life on earth through the anthropocene. Soils, the engine of every terrestrial ecosystem, are themselves wildly diverse subterranean ecosystems providing habitat to countless trillions of micro- and macro-organisms. These organisms themselves create the soil and its functionality by ingesting dead organic matter, thereby breaking it down so that nutrients become available to plants which in turn feed everything else up the food chain. In the everyday processes of foraging, reproducing, exchanging, growing, and dying, the biology upon and within the soil regulates the climate by consuming, transforming, and burying molecules of carbon. This buried and embodied carbon would otherwise be atmospheric carbon dioxide. Thus, it’s not the soil as an apparently homogenous substrate that concerns us. It is the living, biodiverse soil ecosystem that matters for maintaining a global climate system hospitable to human life. In this section, we discuss the contents, processes and functions of the soil and the threats to it, with an eye to highlighting the absolutely critical role of soil biology.
Overview
A noteworthy perspective is that a teaspoon of healthy soil holds more microorganisms than there are people on earth. The biodiversity in that teaspoon rivals that of the Amazon rainforest. In fact, it’s these very microorganisms (along with their larger counterparts, like earthworms and mammals) that themselves create the content and structure of soil by breaking down organic material, secreting sticky biomolecules, and burrowing, while also depending on the soil for habitat and food. Because of the interrelatedness of diverse and vital roles played by soil organisms (roles such as fixing nitrogen, suppressing disease, creating channels for water and root penetration, and forming soil aggregates, for example), the soil as a whole could be viewed as a macro-organism, made up of myriad “organs” working together and becoming sick or healthy as if it were a living body (within a field, for instance) [Magdoff 2009].
Furthermore, the particular community composition of microorganisms in a given soil can affect the overall functioning of the soil. It is generally understood that high concentrations of fungi are associated with healthy soil, while soils regularly disturbed by tillage tend to be bacteria-dominant [Magdoff 2009]. Fungal populations are also associated with increased carbon sequestration.
Kallenbach et al [2016] demonstrate that distinct microbial communities, especially those with high fungal concentrations, are a better predictor than clay mineral content of Soil Organic Carbon (SOC) production. Morrien et al. [2016: 1] show “that during nature restoration the efficiency of nutrient cycling and carbon uptake can increase by a shift in fungal composition and/or fungal activity.” Specifically, the proportion of arbuscular mycorrhizal fungi[5] increases over time leading to greater carbon uptake by fungi, “without an increase in fungal biomass or shift in bacterial-to-fungal ratio.” Recent research by David Johnson of New Mexico State University shows that high fungal to bacterial ratios are correlated with strikingly high carbon sequestration and high plant productivity [Johnson n.d.; Johnson 2017] (see also Croplands section).
Due to the exceptional scope and complexity of its biological activity, soil is a major player in the self-regulating system of Earth’s environment by taking up and releasing large quantities of carbon dioxide, oxygen, methane and other gases [Brady 2008]. Indeed, it is the living organisms on and in the soil that breathe these gases in and out. Under favorable conditions balance is maintained due in part to soil organisms holding carbon in place, both within their own biomass and through ongoing carbon-rich soil creation.
Soils store more carbon than is found in the atmosphere and terrestrial vegetation combined [FAO 2017a]. The capacity for soils to store carbon depends on various factors, such as temperature, moisture level, soil type, and topography. Carbon-rich peatland and organic soil[6], which occupy 3% of land but contain an estimated 30% of the world’s soil carbon, are commonly located in cold environments or rainy, humid environments, where productivity is high and decomposition slow. Another third of the world’s soil carbon is in drylands (areas where potential evaporation/transpiration exceeds average rainfall) which cover approximately 40% of Earth’s land area, but tend toward lower productivity, slower carbon accumulation, and susceptibility to erosion when mismanaged [FAO 2017b].
Ecological processes have maintained a balanced carbon cycle over time, keeping the Earth at a relatively stable temperature for hundreds of thousand of years. Although there have been temperature fluctuations before, it is virtually certain that human activity (mainly burning fossil fuels, desertifying and clearing land) is upsetting the carbon cycle, causing atmospheric concentrations of heat-trapping carbon dioxide to have risen from about 280 parts per million (ppm) prior to the 19th century to more than 400 ppm today [NASA 2017]. This, in turn, is changing the temperature at a faster rate than at any time in at least 1,000 years [Smith 2015]. Since the start of industrial era, carbon emissions from the soil due to deforestation and land conversion is estimated to be 136 Gt (Gigatons, or one billion tons), a little less than half the amount of carbon emissions from fossil fuel combustion over the same time period [Lal 2016]. Estimates of carbon lost to the atmosphere from land use since the beginning of agriculture range as high as 537 Gt [Buringh 1984: 91].
It is important to note that soil that has been greatly depleted of carbon can become a carbon sink when managed according to ecological/restorative principles. A carbon sink is anything that absorbs more carbon than it releases as CO2. “Soils that are depleted of SOC have the greatest potential to gain carbon, but also the least propensity to do so.” [FAO 2017b: 7]This is because, while depleted soils can theoretically regain the entire massive amount of carbon they’ve lost, they first need to be biologically re-activated through restorative management practices. It is vital to note that degraded lands (and all lands) have this capacity for renewal as the global community seeks both to reduce CO2 emissions and to draw down excess atmospheric CO2 as quickly as possible. Conversely, organic soils, forests, and other intact ecosystems with large existing stores of carbon in the soil and vegetation have the potential to become new sources of emissions, and must therefore be properly managed and preserved.
According to the Rodale Institute, the fastest, least expensive and most efficient way to rebuild agricultural soils is through “regenerative organic agriculture,” designed to bring carbon and biodiversity back into the soils.
Recent data from farming systems and pasture trials around the globe show that we could sequester more than 100% of current annual CO2 emissions with a switch to widely available and inexpensive organic management practices, which we term “regenerative organic agriculture.” These practices work to maximize carbon fixation while minimizing the loss of that carbon once returned to the soil, reversing the greenhouse effect. [Rodale Institute 2014: 2] (See more in Croplands section.)
Management practices for building carbon in soils involve increasing input and reducing losses of soil organic matter (SOM). SOM is made up of fungi, bacteria, countless microorganisms, decaying plant matter, decomposing animals, and products formed from their decomposition.SOM also contains humus, an organic material believed to contain stable forms of organic carbon. However, the inherent resistance of humus and other compounds to decomposition and release of carbon dioxide into the atmosphere has recently come into question, as the stability of soil compounds may be an ecosystem property and not an intrinsic property of the compound itself [Schmidt 2011]. Thus the understanding of soils is transformed from a consideration of properties of isolated variables to properties of the system as a whole, with its exceedingly complex interactions, a transformation considered throughout this Compendium.
In sum, SOM provides food for soil organisms and plants, enhances microbial biodiversity, creates pore space, increases cation-exchange capacity (CEC)[7], and increases buffering capacity (ability to resist change in acidity or pH). All of these factors affect the soil’s ability to hold carbon [Fenton 2008].
Soil organic carbon (SOC) levels are directly related to the amount of SOM. The key factors in SOC levels are photosynthesis, respiration and decomposition. Photosynthesis is the process used by plants to harness energy from sunlight, combined with the CO2 they take from the air and water from the ground, and convert it into energy storage (in the form of sugars) for their own growth and survival. Respiration is the measure of CO2 released from soil microbes and plant roots into the surrounding soil.
Soil C results both directly from growth and death of plant roots and above-ground biomass, as well as indirectly from transfer of carbon-rich compounds from roots to soil microbes. The roots of almost all plants form symbiotic associations with arbuscular mycorrhizal fungi – the roots provide the fungi with energy in the form of carbon while the fungi provide the plant with nutrients. While decomposition of biomass by soil microbes results in carbon loss as CO2 from the soil due to respiration, a proportion of the original carbon is retained in the soil through the formation of numerous stable soil carbon molecules. Carbon is also stored in soil aggregates, which are formed in collaboration with microorganisms.
The multiple soil organisms providing vital ecosystem functionsare sensitive to various practices and changing conditions, even those occurring above the soil surface. Applying synthetic nitrogen fertilizer, for example, can affect soil pH, which in turn can negatively affect soil organisms. Organic amendments such as manure and compost, on the other hand, provide direct carbon sources for soil organisms and an indirect carbon source via increased plant growth without negative side effects.
Temperature also affects soil microbes. For example, the scottnema lindsayae nematode cannot survive in its native habitat in Antarctica if temperatures become too warm or too cold, and research suggests that a 65% decline in activity of these nematodes over 12 years could lead to a 30% reduction of carbon cycling in soils [Wall 2014]. Similarly, earthworms, described by Darwin [1881] as “nature’s plow,” are damaged both by cultivation and by the use of toxic herbicides and other agrichemicals often used in “conventional” no-till.
A healthy biosphere has the power to draw down carbon and store water and through this support biodiverse life forms, including humans. Most importantly, in partnership with nature, humanity can restore ecosystems, including agro-ecosystems[8], to create a functional global biosphere once again. This begins with the soil, and requires at once protecting intact organic soils and their invaluable concentrations of stored carbon, and restoring carbon-depleted mineral soils so as to activate their capacity as massive carbon sinks. Remarkably, taking action to protect and rebuild soil is almost universally available, low-cost, safe, and has the power to mitigate and reverse climate change in a relatively short period of time.
Eco-restoration has numerous co-benefits such as restoring abundant food production, eliminating floods and droughts, restoring water supplies, building strong local economies and providing shade, beauty, and recreation.
Soil Article Summaries
Crowther et al. 2016. When this paper first appeared in Nature, it seemed to raise serious objections to the hypothesis that soils as carbon sinks could have a significant impact on climate. On further examination, however, it may be more about the perils of isolating variables than about the limitations of soils.
There is growing confidence that warming generally enhances fluxes to and from the soil, but the net global balance between these responses remains uncertain [Crowther 2016: 104].
The authors analyze data on the soil’s response to warming from 49 experimental sites in North America, Europe and Asia, across six biomes. They find that the level of carbon loss from the soil is contingent upon the size of the soil’s original carbon stock, and on the duration and extent of the warming. Extrapolating to a global scale, they estimate that an additional 1oC temperature rise will release anywhere from 30 ± 30 to 203 ± 161 of carbon Gt from the soil.
The authors themselves highlight several limitations in their analysis due to lack of data, including from experimental sites in the tropics, from soil at depths greater than 10cm, and on the effects from biotic responses to warming. It is noteworthy that, as the authors state:
Our current understanding of global feedbacks is dominated by the physical sciences, but changes in the physiology and community compositions of organisms have been shown to have strong effects on the strength of this feedback [107].
In other words, for a more precise prediction of how global warming will affect the soil’s net release of carbon, we need to understand better how plants, animals and microbes may interact and respond to that same warming with respect to their effect on the land-carbon climate feedback.
Like Amundson et al., these authors make projections about the soil’s response to climate change without consideration of the wide spectrum of possible land management practices, ranging from clearcutting and urbanization to reforestation and Holistic Management. Presumably, a more accurate picture of the soil’s response to future warming must incorporate measurements of the soil’s resilience capability, based on its level of exposure to oxidative or erosive elements vs. embeddedness in intact ecosystems.
Grindrod 2017. A brief review for non-scientists of soil microbiology and the growing understanding that soil health is, in many ways, all about microbes and their complex interactions with mineral, plants and animals. It likely that micro-organisms are the key to soil fertility as a result of the universe of microbial chemistry that affects, among many things, availability of nutrients and plant health, rendering other forms of dangerous inputs such as biocides unnecessary. See also David Johnson [2017].
Hart 2015. In a year of severe heat and drought, cover crops and no-till proved their value for North Carolina Farmer Russell Hedrick, reports Southeast Farm Press. In a county where average SOM is 1.5%, Hedrick’s farm has 4.8% SOM after just four years of no-till and cover crops, which have increased the water infiltration in the soil while also cooling the soil surface as much as 20 degrees F. Hedrick also introduced cattle into his system to graze on the cover crops, a strategy his NRCS[9] advisor told him he “couldn’t afford not to do,” due to the ruminants’ stimulating effects on the life of the soil. During this drought year, while Hedrick’s neighbors lost their crop, he was still able to harvest 110 bushels per acre of corn although it is twice that much in a normal year. Furthermore, Hedrick controls weeds with less herbicide due to the thick mat and allelopathic[10] effects of the five-plant cover crop mix, and he no longer uses any insecticides. He’s also started to see earthworms on his land for the first time, helping to break down organic matter and carry carbon deeper into the soil.
Kallenbach 2016.
Soil organic matter (SOM) and the carbon and nutrients therein drive fundamental submicron- to global-scale biogeochemical processes and influence carbon-climate feedbacks. Consensus is emerging that microbial materials are an important constituent of stable SOM, and new conceptual and quantitative SOM models are rapidly incorporating this view. However, direct evidence demonstrating that microbial residues account for the chemistry, stability and abundance of SOM is still lacking. Further, emerging models emphasize the stabilization of microbial-derived SOM by abiotic mechanisms, while the effects of microbial physiology on microbial residue production remain unclear. Here we provide the first direct evidence that soil microbes produce chemically diverse, stable SOM. We show that SOM accumulation is driven by distinct microbial communities more so than clay mineralogy, where microbial- derived SOM accumulation is greatest in soils with higher fungal abundances and more efficient microbial biomass production. [Kallenbach 2016: Abstract]
Schmidt 2011. Complex interactions, not intrinsic chemical properties, may determine the persistence of soil organic carbon molecules and their climate effects:
Most soil carbon derives from below-ground inputs and is transformed, through oxidation by microorganisms, into the substances found in the soil. By moving on from the concept of recalcitrance and making better use of the breadth of relevant research, the emerging conceptual model of soil organic carbon cycling will help to unravel the mysteries surrounding the fate of plant- and fire-derived inputs and how their dynamics vary between sites and soil depths, and to understand feedbacks to climate change. We argue that the persistence of organic matter in soil is largely due to complex interactions between organic matter and its environment, such as the interdependence of compound chemistry, reactive mineral surfaces, climate, water availability, soil acidity, soil redox state and the presence of potential degraders in the immediate microenvironment. This does not mean that compound chemistry is not important for decomposition rates, just that its influence depends on environmental factors. Rather than describing organic matter by decay rate, pool, stability or level of ‘recalcitrance’ – as if these were properties of the compounds themselves – organic matter should be described by quantifiable environmental characteristics governing stabilization, such as solubility, molecular size and functionalization. . . . Because many, if not most, organic molecules in soils are of microbial origin, experiments are needed that identify the long-term drivers of microbial-cell and microbial-product decomposition, rather than focusing on the immediate fate of fresh plant material. (emphasis added) [Schmidt 2011: 52-3]
This study opens many possibilities for new approaches to soil carbon, including assessment from a systemic as opposed to a reductionist perspective:
More generally, though, the major advances in our understanding of soils will come from research grounded in the theory of many disciplines and in the practice of many approaches. The future research agenda for soils will integrate many different fields and have broader goals than it might have had in the past, with longer time horizons, wider spatial coverage, and an imperative to connect carbon, water and nutrient cycles, so as to understand the soil-plant system as a crucial part of our biosphere. [Schmidt 2011: 55]
Do We Have More Soil for Carbon Storage than We Thought?
This is a discussion of a hypothesis by oligochaetologist[11] Rob Blakemore, Ph.D. He suggests that accounting for varying planetary topography may dramatically increase the projections for soil-carbon storage potential. This has significant implications for the importance of eco-restoration efforts to address climate, and the speed with which we can draw carbon from the atmosphere.
We believe that this hypothesis is both reasonable and logical, but support is largely inferential at the current time. It is our hope that this discussion will stimulate further research into the potential of photosynthetic carbon capture and sequestration in soils, and that this development will motivate an intensification of eco-restoration efforts.
Introduction
Area measurements of the earth’s terrestrial surface are conveniently two-dimensional (2-D), yet the true surface has a certain amount of three-dimensional undulation (3-D). Therefore the actual surface and volume of soils on Earth may be underestimated by an unknown factor using current measurements and models.
This leads to the proposition that global soils may have the capacity to store greater quantities of atmospheric carbon than previously assumed. The role of eco-restoration, with soils as the primary storage medium, with above-ground and below-ground biodiversity as the process for incorporating carbon into soils, is therefore a key to any successful climate strategy.
Why “Flat-Earth” soil estimates may be incorrect
The surface of the earth is conventionally calculated based on an assumption of two-dimensional areas within a set of coordinates. Yet, despite decades of satellite imagery, accurate information on the actual surface area of the earth is surprisingly elusive: the land has undulating terrain adding to its extent. The importance of this distinction is that the total volume of global soils, as well as primary productivity, i.e., photosynthesis, relate to actual soil surface area, not to a two-dimensional model projection thereof.
Actual three-dimensional surface area is most important for calculations of our total topsoil resource or, as Darwin [1881: 49] describes in his ground-breaking book on earthworms: “The vegetable mould . . . covers, as with a mantle, the surface of the land . . .”
It is not unreasonable to estimate a flat projection of photosynthesizing terrestrial surface area of 12 billion hectares (gigahectares or Gha).[12]Some multiple will account for surface irregularities that increase that surface area, and for the sake of discussion we will use a conservative factor of 2.
A practical example of an applicable mathematical estimate of irregular surface areas is from a paint manufacturer [Resene, n.d.], who estimates that compared to a flat surface, a 200 m2 corrugated sheet has 10.5% larger surface area (= 221 m2), and that Anaglypta or Stucco textures, i.e., bumpy like Earth, have surface area 40-100% greater than that of the base area.
One can also use geometry and knowledge of fractals in order to estimate a reasonable multiplier of the available flat topography for illustrative purposes. As a paradoxical (i.e., counter-intuitive) fractal, the actual true land surface area may be infinitely expanded at increasingly finer scales of observation as in a 3-D version of the 2-D “Coastline Paradox” that, in practice, increased the linear distance estimate of Britain’s coastal outline more than six fold[13].
Therefore, this revised estimate of a true land surface takes into account the area exposed to the Sun’s irradiation, and includes all topsoil that supports the plants upon which we depend. If that surface area is doubled by irregularities in the surface of the land, so proportionately is capture of the Sun’s energy and resultant soil activity, including carbon and water storage.
These are very rough approximations: we can say with certainty only that current “Flat-Earth” surface area, and therefore soil volume calculations, are under-representations, and likely significant ones. We look forward to more detailed studies from researchers with the resources to pursue them.
Leaf Area Index
The primary productivity providing for most life on Earth operates at the biological scale of a leaf.[14] Average leaf sizes reportedly range from 0.011 to about 39.5 cm2 but no data are readily available for the topographical surface area and volume of underlying topsoil that supports these plants. An alternative estimate of effective terrestrial surface area is possible if we apply a Leaf-Area-Index (LAI).
LAI is a dimensionless quantity that characterizes plant canopies defined as the one-sided green leaf area over the flat unit ground surface area (LAI = leaf area / ground area). In other words, LAI is a factor that derives the effective ground area for which the plant is productive based on how much photosynthesis is actually turning atmospheric carbon dioxide molecules into above- and below-ground biomass. For example, if the surface area on the ground under a tree occupies 10 m2 and the total leaf surface area is 45 m2, the tree is accomplishing 45 m2 worth of photosynthesis which would be underestimated by only accounting for the tree’s ground-level two-dimensional footprint, as is conventionally reported.
LAI’s range from 0 (bare ground) to ~18 (dense forests) and a global average is 4.5. The authors of this source state that “LAI is a key variable for regional and global models of biosphere-atmosphere exchanges of energy, carbon dioxide, water vapour, and other materials.” [Asner et al. 2003: 195]
For our purposes, we apply LAI to the recalculated undulating and rough-surface topography. If we therefore take our hypothetical but reasonable estimate of a flat 12 Gha of photosynthesizing land and multiply it by 2, we have 24 Gha of non-flat photosynthesizing land surface area. If we include the LAI multiplier of 4.5 to those 24 Gha, we arrive at the equivalent of 108 Gha of photosynthesis, or 9 times more carbohydrate production by green plants than would be estimated from flat-surface-area measurements.
Why does it matter?
We are rapidly losing soils, with global topsoil erosion rates reportedly greater than 2,000 tonnes per second[15] [Pimentel 2013: 447]. Soil is further depleted by agri-chemical pollution and urbanization [Blakemore 2017a], that is, land degradation is due in no small measure to loss of natural soil fertility and excess synthetic nitrogen [Rockström 2009: 472, Fig. 1]. Therefore it is clearly in our best interests, and in the interests of the remaining living organisms on planet Earth, to get accurate information about the 3-D topography of the land, which will alter calculations about surface area and volumes of soils. This in turn will increase the potential for sequestration of carbon and for water storage in soils, vital knowledge that could allow more rapid and effective restoration efforts.
How Much Soil Is There on Earth?
In addition to terrain considerations, we now consider Soil Organic Carbon (SOC) weight in Gigatons, as opposed to previously considered surface area in Gigahectares. Blakemore [2016a: 11] noted that:
Soil carbon values require allowance for intractable glomalin adding a further 5-27% to almost all SOC tallies (Comis, 2002). Plus data from deep soils may increase budgets: e.g., Harper & Tibbett (2013) found C up to five times greater in Australian soils at depth >1 m and down to 35 m in some cases. The Walkley-Black method itself underestimates total C by about 20% with a correction factor of ca. 1.3 often required, whereas latest techniques using mid-infrared (MIR) spectroscopy give more accurate readings. These three factors combined would surely increase SOC totals.
Glomalin, only discovered in 1996, is a stable fungal molecule tightly-bound to soil particles as a major component of soil organic matter. Glomalin adds up to 27% of total SOC [Khursheed 2016], and is stable for 7 to 42 years, depending on conditions [Comis 2002: 4].
Thus an answer to “How much soil is there on Earth?” is still elusive. How much soil can be built on Earth through the activity of healthy biological systems and how quickly these transformations can take place remains an intriguing question. Allowing for glomalin, deep soil data and carbon in living or dead roots [Jackson 1997: T1], soil carbon quantities are likely considerably higher than conventional estimates, as is the potential for future carbon sequestration in soils.
Conclusion
More than two millennia ago Aristotle told us the Earth was not flat and he also concurred with Plato in recognizing that soil erosion and loss of humus and earthworms is catastrophic to civilization [Montgomery 2008: 51]. Leonardo da Vinci’s observation 500 years ago that “We know more about the movement of celestial bodies than about the soil underfoot”seemingly still rings true. An essential feasible and achievable solution is to apply what Sir Albert Howard termed Nature’s Law of Return, i.e., to vermi-compost all organic ‘wastes” to restock the topsoil [Howard 1945].
Most crucially vital, we must determine the amount of living topsoil remaining and its potential restoration through organic regenerative land management worldwide applying principles and practices of Permaculture (Mollison 1988; Blakemore 2017). The challenge now is for professional geographers, astronomers and others to provide actual topographic values for land and topsoil contribution to global photosynthesis and the carbon cycle.
This compendium is, if nothing else, a testament to the key role soils must play if we are to preserve life on earth through the anthropocene. Soils, the engine of every terrestrial ecosystem, are themselves wildly diverse subterranean ecosystems providing habitat to countless trillions of micro- and macro-organisms. These organisms themselves create the soil and its functionality by ingesting dead organic matter, thereby breaking it down so that nutrients become available to plants which in turn feed everything else up the food chain. In the everyday processes of foraging, reproducing, exchanging, growing, and dying, the biology upon and within the soil regulates the climate by consuming, transforming, and burying molecules of carbon. This buried and embodied carbon would otherwise be atmospheric carbon dioxide. Thus, it’s not the soil as an apparently homogenous substrate that concerns us. It is the living, biodiverse soil ecosystem that matters for maintaining a global climate system hospitable to human life. In this section, we discuss the contents, processes and functions of the soil and the threats to it, with an eye to highlighting the absolutely critical role of soil biology.
Overview
A noteworthy perspective is that a teaspoon of healthy soil holds more microorganisms than there are people on earth. The biodiversity in that teaspoon rivals that of the Amazon rainforest. In fact, it’s these very microorganisms (along with their larger counterparts, like earthworms and mammals) that themselves create the content and structure of soil by breaking down organic material, secreting sticky biomolecules, and burrowing, while also depending on the soil for habitat and food. Because of the interrelatedness of diverse and vital roles played by soil organisms (roles such as fixing nitrogen, suppressing disease, creating channels for water and root penetration, and forming soil aggregates, for example), the soil as a whole could be viewed as a macro-organism, made up of myriad “organs” working together and becoming sick or healthy as if it were a living body (within a field, for instance) [Magdoff 2009].
Furthermore, the particular community composition of microorganisms in a given soil can affect the overall functioning of the soil. It is generally understood that high concentrations of fungi are associated with healthy soil, while soils regularly disturbed by tillage tend to be bacteria-dominant [Magdoff 2009]. Fungal populations are also associated with increased carbon sequestration.
Kallenbach et al [2016] demonstrate that distinct microbial communities, especially those with high fungal concentrations, are a better predictor than clay mineral content of Soil Organic Carbon (SOC) production. Morrien et al. [2016: 1] show “that during nature restoration the efficiency of nutrient cycling and carbon uptake can increase by a shift in fungal composition and/or fungal activity.” Specifically, the proportion of arbuscular mycorrhizal fungi[5] increases over time leading to greater carbon uptake by fungi, “without an increase in fungal biomass or shift in bacterial-to-fungal ratio.” Recent research by David Johnson of New Mexico State University shows that high fungal to bacterial ratios are correlated with strikingly high carbon sequestration and high plant productivity [Johnson n.d.; Johnson 2017] (see also Croplands section).
Due to the exceptional scope and complexity of its biological activity, soil is a major player in the self-regulating system of Earth’s environment by taking up and releasing large quantities of carbon dioxide, oxygen, methane and other gases [Brady 2008]. Indeed, it is the living organisms on and in the soil that breathe these gases in and out. Under favorable conditions balance is maintained due in part to soil organisms holding carbon in place, both within their own biomass and through ongoing carbon-rich soil creation.
Soils store more carbon than is found in the atmosphere and terrestrial vegetation combined [FAO 2017a]. The capacity for soils to store carbon depends on various factors, such as temperature, moisture level, soil type, and topography. Carbon-rich peatland and organic soil[6], which occupy 3% of land but contain an estimated 30% of the world’s soil carbon, are commonly located in cold environments or rainy, humid environments, where productivity is high and decomposition slow. Another third of the world’s soil carbon is in drylands (areas where potential evaporation/transpiration exceeds average rainfall) which cover approximately 40% of Earth’s land area, but tend toward lower productivity, slower carbon accumulation, and susceptibility to erosion when mismanaged [FAO 2017b].
Ecological processes have maintained a balanced carbon cycle over time, keeping the Earth at a relatively stable temperature for hundreds of thousand of years. Although there have been temperature fluctuations before, it is virtually certain that human activity (mainly burning fossil fuels, desertifying and clearing land) is upsetting the carbon cycle, causing atmospheric concentrations of heat-trapping carbon dioxide to have risen from about 280 parts per million (ppm) prior to the 19th century to more than 400 ppm today [NASA 2017]. This, in turn, is changing the temperature at a faster rate than at any time in at least 1,000 years [Smith 2015]. Since the start of industrial era, carbon emissions from the soil due to deforestation and land conversion is estimated to be 136 Gt (Gigatons, or one billion tons), a little less than half the amount of carbon emissions from fossil fuel combustion over the same time period [Lal 2016]. Estimates of carbon lost to the atmosphere from land use since the beginning of agriculture range as high as 537 Gt [Buringh 1984: 91].
It is important to note that soil that has been greatly depleted of carbon can become a carbon sink when managed according to ecological/restorative principles. A carbon sink is anything that absorbs more carbon than it releases as CO2. “Soils that are depleted of SOC have the greatest potential to gain carbon, but also the least propensity to do so.” [FAO 2017b: 7]This is because, while depleted soils can theoretically regain the entire massive amount of carbon they’ve lost, they first need to be biologically re-activated through restorative management practices. It is vital to note that degraded lands (and all lands) have this capacity for renewal as the global community seeks both to reduce CO2 emissions and to draw down excess atmospheric CO2 as quickly as possible. Conversely, organic soils, forests, and other intact ecosystems with large existing stores of carbon in the soil and vegetation have the potential to become new sources of emissions, and must therefore be properly managed and preserved.
According to the Rodale Institute, the fastest, least expensive and most efficient way to rebuild agricultural soils is through “regenerative organic agriculture,” designed to bring carbon and biodiversity back into the soils.
Recent data from farming systems and pasture trials around the globe show that we could sequester more than 100% of current annual CO2 emissions with a switch to widely available and inexpensive organic management practices, which we term “regenerative organic agriculture.” These practices work to maximize carbon fixation while minimizing the loss of that carbon once returned to the soil, reversing the greenhouse effect. [Rodale Institute 2014: 2] (See more in Croplands section.)
Management practices for building carbon in soils involve increasing input and reducing losses of soil organic matter (SOM). SOM is made up of fungi, bacteria, countless microorganisms, decaying plant matter, decomposing animals, and products formed from their decomposition.SOM also contains humus, an organic material believed to contain stable forms of organic carbon. However, the inherent resistance of humus and other compounds to decomposition and release of carbon dioxide into the atmosphere has recently come into question, as the stability of soil compounds may be an ecosystem property and not an intrinsic property of the compound itself [Schmidt 2011]. Thus the understanding of soils is transformed from a consideration of properties of isolated variables to properties of the system as a whole, with its exceedingly complex interactions, a transformation considered throughout this Compendium.
In sum, SOM provides food for soil organisms and plants, enhances microbial biodiversity, creates pore space, increases cation-exchange capacity (CEC)[7], and increases buffering capacity (ability to resist change in acidity or pH). All of these factors affect the soil’s ability to hold carbon [Fenton 2008].
Soil organic carbon (SOC) levels are directly related to the amount of SOM. The key factors in SOC levels are photosynthesis, respiration and decomposition. Photosynthesis is the process used by plants to harness energy from sunlight, combined with the CO2 they take from the air and water from the ground, and convert it into energy storage (in the form of sugars) for their own growth and survival. Respiration is the measure of CO2 released from soil microbes and plant roots into the surrounding soil.
Soil C results both directly from growth and death of plant roots and above-ground biomass, as well as indirectly from transfer of carbon-rich compounds from roots to soil microbes. The roots of almost all plants form symbiotic associations with arbuscular mycorrhizal fungi – the roots provide the fungi with energy in the form of carbon while the fungi provide the plant with nutrients. While decomposition of biomass by soil microbes results in carbon loss as CO2 from the soil due to respiration, a proportion of the original carbon is retained in the soil through the formation of numerous stable soil carbon molecules. Carbon is also stored in soil aggregates, which are formed in collaboration with microorganisms.
The multiple soil organisms providing vital ecosystem functionsare sensitive to various practices and changing conditions, even those occurring above the soil surface. Applying synthetic nitrogen fertilizer, for example, can affect soil pH, which in turn can negatively affect soil organisms. Organic amendments such as manure and compost, on the other hand, provide direct carbon sources for soil organisms and an indirect carbon source via increased plant growth without negative side effects.
Temperature also affects soil microbes. For example, the scottnema lindsayae nematode cannot survive in its native habitat in Antarctica if temperatures become too warm or too cold, and research suggests that a 65% decline in activity of these nematodes over 12 years could lead to a 30% reduction of carbon cycling in soils [Wall 2014]. Similarly, earthworms, described by Darwin [1881] as “nature’s plow,” are damaged both by cultivation and by the use of toxic herbicides and other agrichemicals often used in “conventional” no-till.
A healthy biosphere has the power to draw down carbon and store water and through this support biodiverse life forms, including humans. Most importantly, in partnership with nature, humanity can restore ecosystems, including agro-ecosystems[8], to create a functional global biosphere once again. This begins with the soil, and requires at once protecting intact organic soils and their invaluable concentrations of stored carbon, and restoring carbon-depleted mineral soils so as to activate their capacity as massive carbon sinks. Remarkably, taking action to protect and rebuild soil is almost universally available, low-cost, safe, and has the power to mitigate and reverse climate change in a relatively short period of time.
Eco-restoration has numerous co-benefits such as restoring abundant food production, eliminating floods and droughts, restoring water supplies, building strong local economies and providing shade, beauty, and recreation.
Soil Article Summaries
Crowther et al. 2016. When this paper first appeared in Nature, it seemed to raise serious objections to the hypothesis that soils as carbon sinks could have a significant impact on climate. On further examination, however, it may be more about the perils of isolating variables than about the limitations of soils.
There is growing confidence that warming generally enhances fluxes to and from the soil, but the net global balance between these responses remains uncertain [Crowther 2016: 104].
The authors analyze data on the soil’s response to warming from 49 experimental sites in North America, Europe and Asia, across six biomes. They find that the level of carbon loss from the soil is contingent upon the size of the soil’s original carbon stock, and on the duration and extent of the warming. Extrapolating to a global scale, they estimate that an additional 1oC temperature rise will release anywhere from 30 ± 30 to 203 ± 161 of carbon Gt from the soil.
The authors themselves highlight several limitations in their analysis due to lack of data, including from experimental sites in the tropics, from soil at depths greater than 10cm, and on the effects from biotic responses to warming. It is noteworthy that, as the authors state:
Our current understanding of global feedbacks is dominated by the physical sciences, but changes in the physiology and community compositions of organisms have been shown to have strong effects on the strength of this feedback [107].
In other words, for a more precise prediction of how global warming will affect the soil’s net release of carbon, we need to understand better how plants, animals and microbes may interact and respond to that same warming with respect to their effect on the land-carbon climate feedback.
Like Amundson et al., these authors make projections about the soil’s response to climate change without consideration of the wide spectrum of possible land management practices, ranging from clearcutting and urbanization to reforestation and Holistic Management. Presumably, a more accurate picture of the soil’s response to future warming must incorporate measurements of the soil’s resilience capability, based on its level of exposure to oxidative or erosive elements vs. embeddedness in intact ecosystems.
Grindrod 2017. A brief review for non-scientists of soil microbiology and the growing understanding that soil health is, in many ways, all about microbes and their complex interactions with mineral, plants and animals. It likely that micro-organisms are the key to soil fertility as a result of the universe of microbial chemistry that affects, among many things, availability of nutrients and plant health, rendering other forms of dangerous inputs such as biocides unnecessary. See also David Johnson [2017].
Hart 2015. In a year of severe heat and drought, cover crops and no-till proved their value for North Carolina Farmer Russell Hedrick, reports Southeast Farm Press. In a county where average SOM is 1.5%, Hedrick’s farm has 4.8% SOM after just four years of no-till and cover crops, which have increased the water infiltration in the soil while also cooling the soil surface as much as 20 degrees F. Hedrick also introduced cattle into his system to graze on the cover crops, a strategy his NRCS[9] advisor told him he “couldn’t afford not to do,” due to the ruminants’ stimulating effects on the life of the soil. During this drought year, while Hedrick’s neighbors lost their crop, he was still able to harvest 110 bushels per acre of corn although it is twice that much in a normal year. Furthermore, Hedrick controls weeds with less herbicide due to the thick mat and allelopathic[10] effects of the five-plant cover crop mix, and he no longer uses any insecticides. He’s also started to see earthworms on his land for the first time, helping to break down organic matter and carry carbon deeper into the soil.
Kallenbach 2016.
Soil organic matter (SOM) and the carbon and nutrients therein drive fundamental submicron- to global-scale biogeochemical processes and influence carbon-climate feedbacks. Consensus is emerging that microbial materials are an important constituent of stable SOM, and new conceptual and quantitative SOM models are rapidly incorporating this view. However, direct evidence demonstrating that microbial residues account for the chemistry, stability and abundance of SOM is still lacking. Further, emerging models emphasize the stabilization of microbial-derived SOM by abiotic mechanisms, while the effects of microbial physiology on microbial residue production remain unclear. Here we provide the first direct evidence that soil microbes produce chemically diverse, stable SOM. We show that SOM accumulation is driven by distinct microbial communities more so than clay mineralogy, where microbial- derived SOM accumulation is greatest in soils with higher fungal abundances and more efficient microbial biomass production. [Kallenbach 2016: Abstract]
Schmidt 2011. Complex interactions, not intrinsic chemical properties, may determine the persistence of soil organic carbon molecules and their climate effects:
Most soil carbon derives from below-ground inputs and is transformed, through oxidation by microorganisms, into the substances found in the soil. By moving on from the concept of recalcitrance and making better use of the breadth of relevant research, the emerging conceptual model of soil organic carbon cycling will help to unravel the mysteries surrounding the fate of plant- and fire-derived inputs and how their dynamics vary between sites and soil depths, and to understand feedbacks to climate change. We argue that the persistence of organic matter in soil is largely due to complex interactions between organic matter and its environment, such as the interdependence of compound chemistry, reactive mineral surfaces, climate, water availability, soil acidity, soil redox state and the presence of potential degraders in the immediate microenvironment. This does not mean that compound chemistry is not important for decomposition rates, just that its influence depends on environmental factors. Rather than describing organic matter by decay rate, pool, stability or level of ‘recalcitrance’ – as if these were properties of the compounds themselves – organic matter should be described by quantifiable environmental characteristics governing stabilization, such as solubility, molecular size and functionalization. . . . Because many, if not most, organic molecules in soils are of microbial origin, experiments are needed that identify the long-term drivers of microbial-cell and microbial-product decomposition, rather than focusing on the immediate fate of fresh plant material. (emphasis added) [Schmidt 2011: 52-3]
This study opens many possibilities for new approaches to soil carbon, including assessment from a systemic as opposed to a reductionist perspective:
More generally, though, the major advances in our understanding of soils will come from research grounded in the theory of many disciplines and in the practice of many approaches. The future research agenda for soils will integrate many different fields and have broader goals than it might have had in the past, with longer time horizons, wider spatial coverage, and an imperative to connect carbon, water and nutrient cycles, so as to understand the soil-plant system as a crucial part of our biosphere. [Schmidt 2011: 55]
Do We Have More Soil for Carbon Storage than We Thought?
This is a discussion of a hypothesis by oligochaetologist[11] Rob Blakemore, Ph.D. He suggests that accounting for varying planetary topography may dramatically increase the projections for soil-carbon storage potential. This has significant implications for the importance of eco-restoration efforts to address climate, and the speed with which we can draw carbon from the atmosphere.
We believe that this hypothesis is both reasonable and logical, but support is largely inferential at the current time. It is our hope that this discussion will stimulate further research into the potential of photosynthetic carbon capture and sequestration in soils, and that this development will motivate an intensification of eco-restoration efforts.
Introduction
Area measurements of the earth’s terrestrial surface are conveniently two-dimensional (2-D), yet the true surface has a certain amount of three-dimensional undulation (3-D). Therefore the actual surface and volume of soils on Earth may be underestimated by an unknown factor using current measurements and models.
This leads to the proposition that global soils may have the capacity to store greater quantities of atmospheric carbon than previously assumed. The role of eco-restoration, with soils as the primary storage medium, with above-ground and below-ground biodiversity as the process for incorporating carbon into soils, is therefore a key to any successful climate strategy.
Why “Flat-Earth” soil estimates may be incorrect
The surface of the earth is conventionally calculated based on an assumption of two-dimensional areas within a set of coordinates. Yet, despite decades of satellite imagery, accurate information on the actual surface area of the earth is surprisingly elusive: the land has undulating terrain adding to its extent. The importance of this distinction is that the total volume of global soils, as well as primary productivity, i.e., photosynthesis, relate to actual soil surface area, not to a two-dimensional model projection thereof.
Actual three-dimensional surface area is most important for calculations of our total topsoil resource or, as Darwin [1881: 49] describes in his ground-breaking book on earthworms: “The vegetable mould . . . covers, as with a mantle, the surface of the land . . .”
It is not unreasonable to estimate a flat projection of photosynthesizing terrestrial surface area of 12 billion hectares (gigahectares or Gha).[12]Some multiple will account for surface irregularities that increase that surface area, and for the sake of discussion we will use a conservative factor of 2.
A practical example of an applicable mathematical estimate of irregular surface areas is from a paint manufacturer [Resene, n.d.], who estimates that compared to a flat surface, a 200 m2 corrugated sheet has 10.5% larger surface area (= 221 m2), and that Anaglypta or Stucco textures, i.e., bumpy like Earth, have surface area 40-100% greater than that of the base area.
One can also use geometry and knowledge of fractals in order to estimate a reasonable multiplier of the available flat topography for illustrative purposes. As a paradoxical (i.e., counter-intuitive) fractal, the actual true land surface area may be infinitely expanded at increasingly finer scales of observation as in a 3-D version of the 2-D “Coastline Paradox” that, in practice, increased the linear distance estimate of Britain’s coastal outline more than six fold[13].
Therefore, this revised estimate of a true land surface takes into account the area exposed to the Sun’s irradiation, and includes all topsoil that supports the plants upon which we depend. If that surface area is doubled by irregularities in the surface of the land, so proportionately is capture of the Sun’s energy and resultant soil activity, including carbon and water storage.
These are very rough approximations: we can say with certainty only that current “Flat-Earth” surface area, and therefore soil volume calculations, are under-representations, and likely significant ones. We look forward to more detailed studies from researchers with the resources to pursue them.
Leaf Area Index
The primary productivity providing for most life on Earth operates at the biological scale of a leaf.[14] Average leaf sizes reportedly range from 0.011 to about 39.5 cm2 but no data are readily available for the topographical surface area and volume of underlying topsoil that supports these plants. An alternative estimate of effective terrestrial surface area is possible if we apply a Leaf-Area-Index (LAI).
LAI is a dimensionless quantity that characterizes plant canopies defined as the one-sided green leaf area over the flat unit ground surface area (LAI = leaf area / ground area). In other words, LAI is a factor that derives the effective ground area for which the plant is productive based on how much photosynthesis is actually turning atmospheric carbon dioxide molecules into above- and below-ground biomass. For example, if the surface area on the ground under a tree occupies 10 m2 and the total leaf surface area is 45 m2, the tree is accomplishing 45 m2 worth of photosynthesis which would be underestimated by only accounting for the tree’s ground-level two-dimensional footprint, as is conventionally reported.
LAI’s range from 0 (bare ground) to ~18 (dense forests) and a global average is 4.5. The authors of this source state that “LAI is a key variable for regional and global models of biosphere-atmosphere exchanges of energy, carbon dioxide, water vapour, and other materials.” [Asner et al. 2003: 195]
For our purposes, we apply LAI to the recalculated undulating and rough-surface topography. If we therefore take our hypothetical but reasonable estimate of a flat 12 Gha of photosynthesizing land and multiply it by 2, we have 24 Gha of non-flat photosynthesizing land surface area. If we include the LAI multiplier of 4.5 to those 24 Gha, we arrive at the equivalent of 108 Gha of photosynthesis, or 9 times more carbohydrate production by green plants than would be estimated from flat-surface-area measurements.
Why does it matter?
We are rapidly losing soils, with global topsoil erosion rates reportedly greater than 2,000 tonnes per second[15] [Pimentel 2013: 447]. Soil is further depleted by agri-chemical pollution and urbanization [Blakemore 2017a], that is, land degradation is due in no small measure to loss of natural soil fertility and excess synthetic nitrogen [Rockström 2009: 472, Fig. 1]. Therefore it is clearly in our best interests, and in the interests of the remaining living organisms on planet Earth, to get accurate information about the 3-D topography of the land, which will alter calculations about surface area and volumes of soils. This in turn will increase the potential for sequestration of carbon and for water storage in soils, vital knowledge that could allow more rapid and effective restoration efforts.
How Much Soil Is There on Earth?
In addition to terrain considerations, we now consider Soil Organic Carbon (SOC) weight in Gigatons, as opposed to previously considered surface area in Gigahectares. Blakemore [2016a: 11] noted that:
Soil carbon values require allowance for intractable glomalin adding a further 5-27% to almost all SOC tallies (Comis, 2002). Plus data from deep soils may increase budgets: e.g., Harper & Tibbett (2013) found C up to five times greater in Australian soils at depth >1 m and down to 35 m in some cases. The Walkley-Black method itself underestimates total C by about 20% with a correction factor of ca. 1.3 often required, whereas latest techniques using mid-infrared (MIR) spectroscopy give more accurate readings. These three factors combined would surely increase SOC totals.
Glomalin, only discovered in 1996, is a stable fungal molecule tightly-bound to soil particles as a major component of soil organic matter. Glomalin adds up to 27% of total SOC [Khursheed 2016], and is stable for 7 to 42 years, depending on conditions [Comis 2002: 4].
Thus an answer to “How much soil is there on Earth?” is still elusive. How much soil can be built on Earth through the activity of healthy biological systems and how quickly these transformations can take place remains an intriguing question. Allowing for glomalin, deep soil data and carbon in living or dead roots [Jackson 1997: T1], soil carbon quantities are likely considerably higher than conventional estimates, as is the potential for future carbon sequestration in soils.
Conclusion
More than two millennia ago Aristotle told us the Earth was not flat and he also concurred with Plato in recognizing that soil erosion and loss of humus and earthworms is catastrophic to civilization [Montgomery 2008: 51]. Leonardo da Vinci’s observation 500 years ago that “We know more about the movement of celestial bodies than about the soil underfoot”seemingly still rings true. An essential feasible and achievable solution is to apply what Sir Albert Howard termed Nature’s Law of Return, i.e., to vermi-compost all organic ‘wastes” to restock the topsoil [Howard 1945].
Most crucially vital, we must determine the amount of living topsoil remaining and its potential restoration through organic regenerative land management worldwide applying principles and practices of Permaculture (Mollison 1988; Blakemore 2017). The challenge now is for professional geographers, astronomers and others to provide actual topographic values for land and topsoil contribution to global photosynthesis and the carbon cycle.
Earthworms
Although often overlooked, ignored or taken for granted, earthworms are nevertheless keystone soil species, mediators and moderators for rebuilding healthy, biodiverse, high carbon and moisture rich topsoil [Darwin 1881; Blakemore 2016c]. We depend on soils for more than 99% of our food and 100% of our timber and natural fibres [Blakemore 2012, Pimentel 2013]. As an integral part of organic production, earthworms are key to agricultural sustainability and global ecosystem stability. Ancient in origin (probably pre-Cambrian but certainly more than 500 million years old), the 7,000 known species of earthworms are ubiquitous and invariably associated with topsoil humus. Earthworms are a basis of terrestrial food webs and the ultimate detritivor [Blakemore 2016c], recently reinstated as key players in the International “4 per 1000 Initiative” [4p1000.org, n.d.] to increase soil organic matter to store carbon. In this section, we discuss the abundance and variety of earthworms and their role in soil health and functionality.
Overview
Extrapolating data from Darwin [1881], their population numbers around 1.3 x 1015 or 1.3 quadrillion globally with biomass of 0.4 t/ha x 9.5 Gha of productive land = 3.8 Gt. This is about ten times the biomass of all humanity, and twice that of both all domesticated stock and total global fish [Blakemore, 2017]. Forming possibly the largest beneficial animal resource on the planet, earthworms are yet apparently severely depleted by cultivation and agrichemical excesses of industrial farming, often being absent from such soils [e.g. Lee 1985] with both their populations and biodiversity in decline [Blakemore 2016a, b, c].
In comparison to intensive agrichemical farming, studies by Blakemore, [2000, 2016a,b] show a diverse array of up to 23 earthworm species per organic farm site (mean 13 spp), implicated in 16-80% increased crop or pasture yield (mean +39%) plus an average of 12% extra soil moisture storage (range 7-91%) compared to conventional neighbour farms. Carbon sequestration is restored at rates two to three times higher in pasture. Such findings are highly relevant due to looming species extinction and climate change with requirement to meet the needs of a growing population. Organic farming can thus produce higher yields and sequester more carbon.
Earthworms may number up to 1,000~2,000/m2 (10-20 million/ha, or 4-8 million/ac) in fertile soils with biomass as high as 3-5 t/ha, (1.2-2 t/ac ) so earthworm stocks may outweigh the above ground stock [Lee 1985; Blakemore 2016c, 2017]. Earthworm abundance and diversity increase in a truly sustainable system as they convert all organic ‘wastes’ into humus-rich compost while processing all atmospheric CO2 in 12 yr cycles [Blakemore 2016a]. Their burrows, as long as 9,000 km/ha (2250 mi/ac) [Kretzchmar 1982] and up to 15 m in depth (49.2 ft) [Sims & Gerard, 1999: 27, as cited in Blakemore 2016c] aerate, improve water infiltration and, importantly, provide habitats for many other beneficial organisms and microbes that they help distribute throughout the entire soil profile. All rainfall is filtered through their burrows and water is stored in worm-worked humus. Blakemore [2000] found up to 90% extra water in pasture compared to adjacent arable fields, and organic arable soil stored 40% more water than chemically farmed arable soil [Blakemore, Hochkirch 2017].
Wormless soils are economically and ecologically expensive: they need to be plowed regularly, and require extra irrigation plus subsidized artificial chemical nitrogen fertilizers and biocide sprays to fight off plant infections and infestations [Howard 1945; Balfour 1975]. This toxic burden has severe impact upon non-target organisms and any organism fed the crops – including humans – as well as poisoning the soil, air, waterways and oceans. Such findings are summarized in Lady Eve Balfour’s IFOAM presentation in 1977 [Balfour 1977]. Another compelling reason for earthworm conservation is that it is impossible to “geoengineer” by addressing isolated variables the many benefits and essential irreducible systems services that earthworms freely and relentlessly provide. In other words, we have no viable alternative to earthworms.
Soil and Earthworm Relationships
We face a complexity of inter-relating ecological problems.Intensive chemical agriculture is a major GHG contributor (28-50%) and a major source of extraneous CO2 (currently 10-25% and in total historically up to 40%) [Houghton 2010: 338, 348]:
Globally, the conversion of lands to croplands has been responsible for the largest emissions of carbon from land-use change. . . From 1850 to 2000, land use and land-use change released an estimated 108–188 Gt (billion tons) of carbon to the atmosphere, or about 28–40% of total anthropogenic emissions of carbon (274 Gt C from fossil fuels) [Strassmann 2008].
The FAO [Gerber 2013] found that intensive industrial livestock farming (rather than organic husbandry) contributed 14.5% of human-induced GHG emissions. A newspaper report [Bryce 2013] comments:
The FAO’s last livestock report, a 2006 assessment titled Livestock’s Long Shadow, found that farms breeding chickens, pigs, and cows for meat and dairy products, produced a disconcerting 18% of global greenhouse gas emissions . . . Around 30% of global biodiversity loss can be attributed to livestock production, such as the spread of pasture land or turning over forests and savannahs.
Although these figures vary due to different formulas for budgeting, it’s clear that agriculture in all its forms, including the practice of forest clearance, is a major contributor to GHG emissions.
The traditional, innovative & scientific methods of non-chemical, organic farming and Permaculture appreciate the importance of earthworm conservation [Howard 1945; Balfour 1975; Mollison 1988]. As a key player in natural processes and crucial issues, Darwin’s “lowly earthworm”, although neglected, warrants re-ascendency to its former position as premier farm livestock [Howard 1945]. For our own health and for that of our planet, we urgently need wholly natural vermi-composting at all scales (from kitchen to continent) in order to replace synthetic fertilizers and to facilitate rapid transition to broad-acre organics that also has earthworm livestock at its core. Enabling earthworms to restore healthy soils is vital to stabilizing climate. All organic ‘wastes’ and manures should be recycled via vermi-composting and appropriate management employed to enhance populations of field-working worms.
Earthworm Article Summaries
van Groenigen et al. 2014.In a recent meta-analysis, while not considering organic farming or carbon per se, this study confirmed earthworm presence corresponding to crop yield increases of 25%, which is comparable to average ~39% extra organic yield in soils with earthworm proliferations determined by Blakemore [2000, 2016b]. This supports earlier studies by Wollny [1890: Forschungen auf der Gebiet der Agrikultur-Physik, 13, s. 381] that found addition of earthworms to soil led to a marked increase of cereal grain by 35-50% and of straw by 40%.
Solomon 2013.
Although earthworms are beneficial in gardens and agricultural fields, they are harmful to Michigan’s forests where they are an invasive species. . . . Earthworms are not native to Michigan and the Great Lakes region, at least not since before glaciers covered the region; they were brought here during European settlement in the 1800s or possibly earlier. Plants, wildlife and forests evolved without any of these creatures around. They are now an invasive species that harms forests.
Hardwood forests without earthworms have a thick layer of slowly decomposing leaves, or “duff” that promotes a rich community of wildflowers, tree seedlings and small animals. Earthworms change that environment dramatically by essentially consuming the duff, thereby destroying habitat and reducing fertility. In contrast to their effect in gardens, earthworms cause forest soils to become more compacted. As a result of habitat loss, fertility declines and soil compaction, these forests may be less productive and have poorer new tree regeneration in the long run.
Another view, from oligochaetologist (worm scientist) Rob Blakemore, is as follows:
Regarding popularized concerns about alien Asian invasive worms threatening to destroy American native forests, this may reasonably be regarded as part of a process that is commonly known as Ecological Succession [Odum 2005].
Ironically, the ecological concept of succession started with Thoreau and Cowles on studies of forest succession and on the Lake Michigan dunes. Large parts of the northeastern North America were glaciated up to about 10,000 years ago completely destroying all land surfaces and forming the Great Lakes. When the ice retreated Nature returned in successive waves and, gradually, the soil, vegetation, and animals communities re-established and species continue to evolve.
According to Darwin [1881] earthworms are supremely important for natural productivity and for the recent progress of human civilizations. In this context the woodlands of Michigan seem a relatively minor issue compared to species extinction and climate change. Healthy soils generally harbour earthworms and it appears there had been insufficient time for these slow-moving and flightless organisms to colonize without fast-track via incidental intervention of most-recent human settlers, often as anglers on the Great Lakes.
When exotic crops and stock were introduced around the world 10,000 years ago, so too were attendent earthworms and these have now spread to “pristine,” albeit transitional, woodlands. The many benefits earthworms have for agricultural and unmanaged soils may cause some changes in more natural habitats but this is a virtually unavoidable and irreversible force majeure and fact-of-life.
Certainly there will be a new ecological balance in time, possibly at a different level of productivity and biodiversity. That is the definition of succession.
Héry et al. 2008. Earthworms have been observed to increase methanotrophy (methane metabolic breakdown) in soil covering a landfill; this is most likely “due to the stimulation of bacterial growth or activity than to substantial shifts in the methanotroph community structure” [Hery 2008: 92].
Earthworm-mediated bioturbation has been linked to an increase in methanotrophy in a landfill biocover soil (AC Singer et al., unpublished), but the mechanism of this trophic interaction remains unclear. The aims of this study were to determine the composition of the active methanotroph community and to investigate the interactions between earthworms and bacteria in this landfill biocover soil where the methane oxidation activity was significantly increased by the earthworms [Hery 2008: 92].
And
We proposed the hypothesis that earthworms could stimulate the growth or the activity of methanotrophs. We showed that the earthworm-mediated increase of methane oxidation in the landfill biocover soil only weakly correlated with a shift in the structure of the active methanotroph population. Future work needs to focus on the relationship between this earthworm effect on enhanced methane oxidation in landfill cover soil and this effect on bacterial activity and growth. The possible contribution of an enriched population of nitrifying bacteria to methane oxidation also requires further investigation [Hery 2008: 101].
Water
With the rise of civilizations, humans began having significant impacts on bodies of water and the water cycle. The early “hydraulic civilizations” appeared along major rivers (Nile, Tigris-Euphrates, Indus, Yellow River and others), changed watercourses and built canals for agriculture and transportation. As populations and cities expanded, demand for food led to soil depletion while the built environment created growing areas of impermeable surfaces. Disruption of water cycles has reached a peak since the industrial revolution, with large areas of land covered with impermeable surfaces, and rainwater and waste rapidly shuttled away from land into the oceans. [Kravčik 2007: 42 ff.]
Eco-restoration requires a shift in thinking about water management and fortunately there are many successful water restoration projects under way, along with a strong theoretical basis to guide them. Water management is the key to cooling the biosphere, regenerating photosynthesis and drawing down carbon on billions of acres. Included in this section is a brief discussion of wetlands, which will be expanded in the next Compendium release.
Several eco-restoration examples are included here, and many were explored at our 2015 Restoring Water Cycles to Reverse Global Warming Conference at Tufts University – all speaker videos are available online. https://bio4climate.org/program-tufts-2015/.
Overview
Healthy Soils Australia, Walter Jehne 2015. Walter Jehne is a microbiologist, soil and climate scientist who has spent the past several years teaching and promoting the use of nature’s tools to address destruction of land and water cycles, and educating on methods important in addressing global warming. The text below represents excerpts (condensed and edited) from the paper, “Restoring Regional Rainfalls: Background Brief for Outcomes, Australia Forum on Vegetation-Rainfall Relationships”:
Contrary to the dominant assumptions that global warming is due to elevated atmospheric carbon concentrations,
Systemic climate changes such as aridification began in the 1970s well before CO2 levels or its temperature effects increased abnormally.
Water-based processes govern most climate effects and over 95% of the earth’s heat dynamics for billions of years, including some 80% of the natural greenhouse effect.
These hydrological heat dynamics have been ignored or deemed “secondary feedbacks” to the CO2 greenhouse effect because water is assumed to be a dominant determinant of our climate, and humans could not possibly have altered these global cycles to cause the anthropogenic climate changes
The hydrological processes are highly complex and difficult to model, and were therefore excluded in IPCC (Intergovernmental Panel on Climate Change) assumptions and models, whereas the minor CO2 component of the greenhouse effect is more readily modelled, and provides a simple marketable explanation of its “cause.”
Because of these IPCC assumptions, policy and response options have largely ignored the dominant hydrological determinants on climate, the effects of land management, and impacts these changes may have on climate, water and bio-system stability.
Yet we have greatly altered the earth’s natural hydrology and thus heat dynamics by:
Clearing over 75% (6.3 billion hectares or 15.75 billion acres) of the earth’s primary forest, greatly altering the land’s albedo and heat reflectance as well as transpiration and latent heat fluxes that cooled vast regions.
Oxidizing and eroding organic matter from some 10 billion hectares of soils thereby reducing the ability of landscapes to infiltrate, retain and supply water to sustain cooling transpiration and latent heat fluxes and the drawdown of carbon from the air by plants via photosynthesis.
Exposing vast areas of such degraded, cropped and bare soils to erosion which has dispersed 3 billion tonnes of additional dust aerosols into the air where it nucleates warming humid hazes that retain heat in the biosphere.
Heating bare exposed soils to greatly increase their re-radiation of heat which massively increases greenhouse warming effects.
Increasing the absorption of solar radiation by humid haze micro-droplets [resulting in] global dimming (while in the liquid phase), as well as the absorption of re-radiated heat (while in the gaseous phase) to warm the lower atmosphere via the water vapour greenhouse effect.
Reducing regional rainfalls often by up to 30% due to the increase in persistent haze micro-droplets which are too small to coalesce into raindrops and precipitate by themselves.
Increasing surface humidity due to the persistent humid hazes, thus lowering evaporation rates by up to 10% and reducing latent heat fluxes which transfer heat out of the biosphere into space.
Reducing the production of the biological precipitation nuclei from forests that would help coalesce the humid haze micro-droplets to form dense clouds with high albedos that reflect 33% of solar radiation out to space, thereby regulating global temperatures.
Preventing the nucleation of haze and cloud droplets into raindrops which lowers rainfalls and the level and longevity of transpiration, photosynthesis and cooling latent heat fluxes.
Impairing the night-time escape of re-radiated heat to space via natural “radiation windows” due to the impaired nucleation and precipitation of such “blocking” humid micro-drop hazes.
Increasing sustained high pressures above the cleared, bare heated soils that prevents the inflow of cool moist air from oceans, its precipitation and the associated cooling heat fluxes.
Extending such high pressure over vast regions and periods to accentuate the aridification of bio-systems which readily collapse to deserts with further human land degradation. [Healthy Soils Australia 2015: 1-2]
Given this reality we need solutions that go beyond just reducing future CO2 emissions but also:
Cool regions and the climate so as to offset dangerous warming and its feedback effects.
Draw down carbon back into its safe soil sinks so as to reduce its greenhouse effect.
Restore the resilience of agro-ecosystems and communities to the extremes and secure their essential water, food and bio-material needs via just, safe low carbon futures. [Healthy Soils Australia 2015: 8]
Regenerate natural hydrological processes by land management which captures water in soils, wetlands, aquifers and biomass
Maintain healthy biodiverse soils to Restore microbial drivers that govern these cooling hydrological processes by emitting condensation nuclei that lead to rainfall.
Support the biological sequestration of carbon from the air into stable soil humates and glomalin to enhance the water held in the soil reservoirs that sustain the cooling latent heat fluxes.
Support the production of microbial precipitation nuclei that coalesce the warming humid hazes into dense high albedo clouds that cool regions and generate critical cooling rainfalls.
Promote the nucleation and enhancement of rainfall in key regions to maintain the latent heat fluxes, green vegetated habitats and the radiation windows that enhance nighttime cooling effects.
Only by regenerating our forests, soils and landscapes can we now restore the hydrological cooling processes that helped govern the natural heat dynamics and buffered climate of the blue planet. Such regeneration is now our only option to offset the dangerous climate feedbacks resulting from the warming induced by our landscape degradation and its associated abnormal rise in CO2 levels.
Fortunately viable practical options exist to enable us to do and directly benefit from this, at grass roots community level: tree by tree, hectare by hectare, region by region. While we face a global emergency and must all take responsibility for it, it can only be addressed locally via practical action on the ground by communities driven by their own self interest in securing a safe climate and future.
The good news is we can still avoid the pending extremes and collapse provided we focus on direct local action urgently to regenerate the health of each square metre of soil and each forest and tree. We have the abundant degraded land, sunshine, CO2, waste biomass and nutrients to do it with. To grow more green areas; by regenerating our soils, forests, rangelands and even re-greening deserts.
We can do this if we enhance the infiltration, retention and availability of each raindrop in our soils so that the regenerated ‘in soil reservoirs’ sustain healthy green growth over larger areas for longer. This will happen naturally, synergistically, as plant growth enhances the structure of the soil by increasing its carbon content which in turn aids its water holding capacity and nutrient dynamics.
Just as nature did over the past 420 million years in colonizing and greening the earth’s land surface, these same processes are now our only option to regenerate our soils, forests and landscape and re-secure our safe climate and future. [Healthy Soils Australia 2015: 7-11]
Water Article Summaries
Ellison 2017. “Trees, forests and water: Cool insights for a hot world” may be one of the few articles in the mainstream literature relating to climate that puts hydrological cooling effects before carbon in importance for addressing global warming, although dynamics of water and carbon are closely intertwined.
Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. [Ellison 2017: 51]
This paper is discussed further under Forests.
Evans, Griggs 2015. Carol Evans is a fisheries biologist and Jon Griggs is a rancher in northeastern Nevada. They have worked together over twenty-five years to restore overgrazed lands to health through planned grazing of cattle, which also brings water, trout, beavers and biodiversity to the riparian areas of Maggie and Susie Creeks. In the driest state in the U.S., with less than ten inches of rain a year, they now have perennial streams and wet meadows after five years of the worst drought in memory.
Susie Creek, ca. 1989 (left) and 2015 (right) after five years of drought. Elko, Nevada
Kravčík 2007. Michal Kravčík and co-authors are Slovakian hydrologists who have developed what they call a new water paradigm for managing water cycles, floods and drought.
In a healthy water cycle, while some rain enters streams and rivers directly and is carried off to sea, most rain water is absorbed by the soils in situ, where it lands. The rain gives life to the soil and sets many biological processes in motion, where it is essential for stable soil carbon storage and cooling the biosphere. This includes evapotranspiration from plants which returns water as vapor to the atmosphere where the water condenses and falls as rain. The cycle then begins anew. Kravčík et al. call this the “small water cycle” (i.e., local water cycle) where most water goes through its cycles in a regional area or smaller. The “large water cycle” is the exchange of water between oceans and land, and “above land water circulates at the same time in many small water cycles which are subsidized with water from the large [continental or global] water cycle.” [Kravčík 2007: 16]
Civilizations disturb healthy water cycles and accelerate the runoff from land by creating impermeable surfaces (including degraded farmlands and rangelands), and preventing water from remaining in place to sink into soils or to forcing it to run off the land, causing floods and often carrying valuable topsoil with it. Furthermore, water systems have been engineered to move water away from its source to the oceans. Water, with its growing use as a means to dispose of farming, industrial and human wastes, is even seen as a nuisance. As a result, less water returns to continents from the oceans than is lost from continents to oceans, which leads to desiccation of soils, severe drought, wildfires, desertification, and a measure of sea-level rise. There is a growing understanding that these phenomena, often attributed to climate change, may in fact also be a function of disrupted water cycles.
Restored urban land, Kosice. November 2005 (left), September 2006 (right).
Heat from the sun drives these earthly water cycles. Small water cycles are local, circulating water within a relatively small area. Latent heat causes water to evaporate; heat is absorbed in the process of evaporating water and does not result in an increase in local temperature. We thus do not experience latent heat as an increase in temperature. However, when there is less water available for evaporation, less solar energy is transformed into latent heat and more solar energy is transformed into sensible heat, heat you can feel as increased temperature. This is the heat that we are increasingly experiencing as global warming.
A great deal of heat is moved from the surface of the earth to the upper atmosphere by evaporation and transpiration of water by plants, contributing to significant cooling of the biosphere – to illustrate it takes 540 calories to turn 1 gram of water to vapor; by comparison it takes only 80 calories to melt 1 gram of ice. Draining of land, that is, runoff and floods, can be reversed through comprehensive conservation of rainwater which maintains the sponge-like absorption capacity of soils and maintains many aspects of soil health, resilience, biodiversity and productivity. Renewal of small water cycles over land can temper extreme weather events and ensure a growth in water reserves by eliminating heat islands and problematic distribution of atmospheric moisture.
Nobre 2010. Antonio Nobre is an Amazon scientist who has studied the biotic pump (see also Makarieva), and tells how he was once told by an indigenous wise man,
“Doesn’t the white man know that, if he destroys the forest, there will be no more rain? And that if there’s no more rain, there will be nothing to drink, or to eat?” I heard that . . . [ and thought], “Oh, my! I’ve been studying this for 20 years, with a super computer; dozens, thousands of scientists, and we are starting to get to this conclusion, which he already knows!” A critical point is the Yanomami have never deforested. How could they know the rain would end? This bugged me and I was befuddled. How could he know that?
Some months later, I met him at another event and said, “Davi, how did you know that if the forest was destroyed, there’d be no more rain?” He replied: “The spirit of the forest told us.”
The equatorial region, in general, and the Amazon specifically, is extremely important for the world’s climate. It’s a powerful engine for evaporation. From a satellite viewpoint, atmospheric water flow can look like a geyser, which is underground water heated by magma transferred into the atmosphere. There are no geysers in the Amazon but trees play the same role. They, like geysers, transfer an enormous amount of water from the ground into the atmosphere. Nobre continues:
There are 600 billion trees in the Amazon forest, 600 billion geysers. That is done with an extraordinary sophistication. They don’t need the heat of magma. They use sunlight to do this process. On a typical sunny day in the Amazon a large tree manages to transpire 1,000 liters of water. If we take all of the Amazon, which is a very large area, and add up all the water that is released by transpiration, “the sweat of the forest,” an incredible amount of water is evaporated into the atmosphere: 20 billion metric tons of water per day. . . . This river of vapor that comes up from the forest and goes into the atmosphere is greater than the Amazon River.”
The Amazon River itself is the largest river on Earth, it carries one fifth of all the fresh water, it releases 17 billion metric tons of water a day into the Atlantic Ocean, smaller than “the river in the sky.” To evaporate the 20 billion tons of water released by trees it would take 50,000 of the largest hydroelectric plant in the world, Itaipus, which generates 14 GW of electricity, 30% of Brazil’s power. The Amazon does this with no technology, at no cost.
Schwartz 2016. Judith Schwartz once again travels the world to collect stories of lands restored, of lives revived, this time to glean insight from restorers of water. She demonstrates that many of our assumptions about managing water are derived from engineering, not biology. When biology is the focus of the water and rainfall question the problem is redefined, and clarified. Solutions that had been invisible become apparent, and provide the opportunity for far more effective responses – even in some of the driest places on earth. Floods and droughts become manageable, even preventable entirely. Two of the innovators mentioned in Water in Plain Sight, Michal Kravčík and Rajendra Singh, spoke at Biodiversity for a Livable Climate’s 2015 Restoring Water Cycles conference, as did Judy.
Singh 2007. Rajendra Singh, the “Water Man of India,” tells the story of how he helped over 1,000 villages restore water and abundance through the use of ancient, low-technology land management. Providing water for people, farms and animals, such efforts countered the ill effects of industrialization and reversed flight to the cities. Says Singh:
I am neither a scientist, nor a professional water engineer nor a climate change expert. I am a small constructive worker of Gandhi and I mobilize the civil society and the community for action on natural resources management and conservation for rural uplift in India. Here I am recording the impact of the above work on the ecology of 6,500 square km area in Alwar district from 1985‐2007. Since 1985, 8,600 small water harvesting talabs [a form of check dam] in 1,068 villages of Alwar district covering 6,500 square km area have been built. This has resulted in the shallow aquifer recharge in groundwater bringing up the water table from about 100‐120 meters depth to 3‐13 meters at present. The area under single cropping increased from 11 per cent to 70 per cent out of which area under double cropping increased from 3 per cent to 50 per cent bringing prosperity to the farmers. The forest cover, which used to be around 7 per cent increased to 40 per cent through agro‐forestry and social forestry, providing sufficient fuel wood and sequestering carbon from atmosphere [Singh 2007: 5].
A dramatic example of how large restoration efforts are built from small, local efforts. In the ten years since this paper, Tarun Bhagat Sangh has continued to expand its work.
Wetlands
Note: As mentioned in the Release notes, we have a small staff and therefore have had to postpone some important material to the next release, scheduled for January 2018. This will include a more thorough exploration of the importance of wetlands in addressing climate.
Wetlands only cover only a small proportion of the terrestrial surface area, with estimates ranging from 5-8% [Mitsch 2007]. Despite this, they store a disproportionate amount of Earth’s soil carbon, with estimates ranging from 20-30% [Lal 2008]. Soil carbon in wetlands can be as high as 40% [Vepraskas and Craft 2016; cf. Nahlik 2016: 2], compared to typical estimates for agricultural soils that range up to 2% [Lal 1995; see the Croplands section of this Compendium for a detailed discussion of agricultural soil carbon]. High carbon storage in wetlands is the result of anoxic conditions in wetland soils that lead to slowed decomposition and a resulting increase in stored organic matter. Wetlands have significant potential to act as carbon sinks under the right circumstances. Potentially reducing the carbon sequestration potential for wetlands are the methane emissions that they produce. Notwithstanding the uncertainty of methane emissions, wetlands globally likely serve as a significant net carbon sink. With the complexity of processes that contribute to wetland carbon exchange and resulting function as GHG sinks or sources, management decisions may be a critical factor in determining the carbon sequestration outcome of wetland ecosystems. [See Nahlik 2016 for review of quantitative data]
Overview
The importance of wetlands cannot be overstated. They exist in all biomes ranging from the tropics to the tundra, and on every continent except Antarctica . Wetlands also tend to host much higher concentrations of biological diversity than other ecosystems. The fact that the oldest intergovernmental environmental convention, the Ramsar Convention of 1971, was focused explicitly on wetland conservation is a testament to this importance [Mitra 2005].
The comparison of carbon dioxide storage to methane emissions is a source of uncertainty in determining the carbon sequestration potential for wetlands. Given methane’s stronger radiative forcing as a greenhouse gas, the IPCC currently uses a ratio of 25:1 to indicate the methane to carbon global warming potential. Despite the high global warming potential presented by methane, it has a relatively short atmospheric “lifetime”, often reported in the range of 8-12 years. Although methanotrophic (methane metabolizing) bacteria are not typically calculated into the equations regarding the atmospheric lifetime of methane, bacteria that metabolize atmospheric methane (e.g., high-affinity oxidation methanotrophs) provide an additional mechanism hastening methane attenuation [Jardine 2009].
Several trends appear to be instructive in wetland carbon storage. Tropical and temperate wetlands generally hold more carbon than northern boreal peatlands [Mitsch 2012: 7-9]. Freshwater inland wetlands may hold more carbon than coastal saline wetlands, though this may be based primarily on their surface area extent. More carbon storage occurs at depths from 30 – 120 cm (1-4 ft), at least as reported by an analysis of U.S. conterminous wetlands in which 65% of the total carbon was deeper than 30 cm (1 ft) [Nahlik 2016: 2-3]. A correlation exists between increased anthropogenic disturbance and decreased carbon storage, although this may be an artefact of settlement patterns rather than a causal relationship [Nahlik 2016: 4]. One study reported high carbon sequestration and low methane emissions in constructed wetlands [Mitsch 2012]; this is an encouraging result considering the many wetlands created as mitigations to compensate for human development impacts.
Wetland Article Summaries
Apfelbaum 1993. Steve Apfelbaum of Applied Ecological Services, Inc. is a restoration ecologist with several decades of experience around the world. This brief paper, “The Role of Landscapes in Stormwater Management,” describes the historical condition of wetlands in the upper midwest, the degrading effects of agriculture and urbanization on water cycles, vegetation and the resultant pollution. Included are recommendations for restoration of healthy wetlands and methods for slowing the movement of water so that it may keep soils hydrated and feed local ecosystems. In addition, restoration of wetlands includes high-capacity carbon storage due to low-oxygen conditions in wetland soils, with subsequent low rates of oxidation and loss of carbon to the atmosphere.
This paper presents evidence that many existing streams did not have conspicuous
channels and were not identified during pre-settlement times (prior to 1830s in the Midwestern United States). Many currently identified first, second, and third-order streams were identified as vegetated swales, wetlands, wet prairies, and swamps in the original land survey records of the U.S. General Land Office.
The data presented show that significant increases in discharge for low, medium, and high flows have occurred since settlement. Stream channels have formed inadvertently or were created to drain land for development and agricultural land uses. Currently, discharges may be 200 to 400 times greater than historical levels, based on data from 1886 to the present for the Des Plaines River in Illinois, a 620-square-mile watershed. Historic data document how this river had no measurable discharge or very low flow conditions for over 60 percent of each year during the period from 1886 to 1904.
This study suggests that land-use changes in the previous upland/prairie watershed have resulted in a change from a diffuse and slow overland flow to increased runoff, concentrated flows, and significantly reduced lag time. Preliminary modeling suggests the following results: reduced infiltration, reduced evaporation and evapotranspiration, greatly increased runoff and hydraulic volatility, and increased sediment yields and instream water quality problems caused by destabilization of streambanks.
The opportunity to emulate historical stormwater behavior by integrating upland landscape features in urban developments and agricultural lands offers stormwater management options that are easier to maintain, less expensive over time, attractive, and possibly more efficient compared with many conventional stormwater management solutions and the use of biofiltration wetlands.
and
Diverse and productive prairies, wetlands, savannas, and other ecological systems occupied hundreds of millions of acres in presettlement North America. These ecological systems have been replaced by a vast acreage of tilled and developed lands. Land-use changes have modified the capability of the upland systems and small depressional wetlands in the uplands to retain water and assimilate nutrients and other materials that now flow from the land into aquatic systems, streams, and wetlands. The historical plant communities that were dominated by deep-rooted, long-lived, and productive species have been primarily replaced by annual species (corn, soybeans, wheat) or shallow rooted non-native species (bluegrass lawns, brome grass fields). The native vegetation was efficient at using water and nutrients, and consequently maintained very high levels of carbon fixation and primary productivity. Modern communities, in turn, are productive but primarily above ground, in contrast to the prairie ecosystem where perhaps 70 percent of the biomass was actually created below ground in highly developed root systems. These changes in the landscape and vegetation coupled with intentional stormwater management have changed the lag time for water to remain in uplands and consequently increased the rate and volume of water leaving the landscape.
Mitsch et al 2012. This study evaluated the carbon storage and methane exchange potential for seven wetlands based on field data collected over several years, and used field data collected at 14 other wetlands globally to model the carbon sequestration and methane emission potential out to 300 years. A total of 21 wetlands were examined. The modeling accounted for the anticipated half-life of methane oxidative degradation in the atmosphere. Results indicated that methane emissions become unimportant within the 300 year model simulation time range, with most wetlands making the shift to net carbon storage by year 100 of the model. The study supports the potential for wetlands as carbon sinks.
Nahlik and Fennessy 2016. The objective of this article was to quantify the carbon stocks present in wetlands of the conterminous United States. To do so, the authors examined empirical field data collected during the 2011 National Wetland Condition Assessment conducted by the U.S. Environmental Protection Agency, and used this to quantitatively extrapolate to larger scale carbon estimates. These estimates were developed at regional and national scales. Results were evaluated by region, wetland type, freshwater or tidal status, and level of anthropogenic disturbance. The article indicated that 11.52 gigatons of carbon are present in the U.S., much of which is in soils deeper than 30 cm (1 ft). Freshwater wetlands located inland held nearly ten-fold as much carbon as intertidal wetlands overall, although this is at least partly due to the much greater aerial extent of inland freshwater wetlands; tidal wetlands still had higher concentrations of carbon storage. The authors also indicate a possible relationship between anthropogenic disturbance and carbon stocks, wherein less disturbed sites store more carbon. Insufficient data was available to determine whether this was a causal effect or an artefact of some kind, such as human preference in settlement patterns. The authors conclude that, due to the substantial carbon stocks that wetlands represent and the potential for anthropogenic impacts, existing intact wetlands should be protected to avoid the risk of further contributing to climate change.
Croplands
Cultivated land covers 1.6 billion hectares globally [FAO 2011]. About 62% of cropland produces food directly for human consumption, while 35% is dedicated to producing animal feed, and 3% to biofuel feedstock, seed and other industrial products [Foley 2011: 338]. Agriculture is a major source of emissions of carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O), contributing 10-12% (including crop and livestock production) of total greenhouse gas emissions [Smith 2007]. Agricultural emissions are driven by the globally dominant industrial model, which favors monocultures and fossil-fuel intensive inputs, and results in soil organic carbon loss and overall soil degradation. However, rather than being a source of carbon emissions globally, agriculture can become a powerful carbon sink. This section looks at the carbon sequestration outcomes of farming practices, such as cover cropping, agroforestry and no-till, which are designed to minimize erosion and boost soil biodiversity, thus restoring soil ecosystems to health and resilience. While more research is needed on holistic approaches that combine multiple soil-building practices, such as permaculture and agroecology, recent research suggests that restorative agriculture could sequester “more than 40% of annual emissions (an estimated 21 Gt CO2 each year [5.7 Gt C/year])” [Rodale 2014], and likely far more, as indicated below.
Cultivation thus began an ongoing slow ignition of
Earth’s largest surficial reservoir of carbon [16]
Overview
The purpose of this compendium, once again, is to emphasize possibilities, the “positive deviants” which lead us to expand our conceptual limits. Only when we can conceive of exceptional and inspiring outcomes may we find the motivation to overcome obstacles to attain them. Fortunately the evidence that supports regenerative land management is rapidly growing, and there are indications that it may outpace climate disruption and provide us with the time and opportunity to address the many difficult circumstances resulting from widespread eco-destruction, including the poster child, global warming. In this section we address the challenges of croplands and their ability to capture atmospheric carbon and recover quickly from millennia of mistreatment.
Under careful human management it is possible for soil organic carbon to reach amounts greater even than under natural, pre-agricultural conditions. A classic example is the Terra Preta soils of the Amazon, “where intensive management and high levels of organic matter additions were practiced over many years, resulting in greatly enhanced soil C” [Paustian 1997: 231].
In spite of a long history of soil carbon loss and a body of scientific literature that views carbon-poor soils as “normal,” many examples of building high levels of soil carbon exist among today’s ecologically minded land managers. California Farmers Paul and Elizabeth Kaiser, for instance, use 5-10 times more compost than average, never till, rotate fields with an extremely diverse mix of vegetable varieties, surround their crops with native trees, shrubs and flowers and have thus built up a thick topsoil containing 10% SOM [Oppenheimer 2015; Kaiser 2017].
In most scientific studies, carbon sequestration rates for croplands measure below 1t C/ha/yr (0.4t C/ac/yr), despite some exceptions as highlighted below. Leading soil scientist Rattan Lal [2016] estimates the global sequestration potential for cropland soils to be 0.8 to 1.2 t C/ha/yr, or “as much as 62 t/ha over the next 50 to 75 years … with a total C sink capacity of ~88 Gt on 1,400 Mha” [Lal 2016: 20A]. That amounts to an average annual global sequestration rate between 1 and 2 Gt C/year, compared to annual carbon emissions from fuel combustion and land use conversion of more than 10Gt C/yr [Lal 2016]. Similarly, Smith [2008] estimates that, under improved management, agriculture could offset 20% of global emissions. Both authors note that conservation-oriented agricultural is a small, albeit crucial, piece of the whole climate mitigation puzzle.
Yet, for a couple of important reasons, these estimates likely greatly underestimate the potential of global croplands to absorb carbon. First, samples are commonly taken to a depth of 30cm or less [Torres-Sallan 2017; Minasny 2017]. This is the default sampling depth recommended in the 2006 IPCC Guidelines for National Greenhouse Gas Inventories, despite acknowledgement in these same guidelines that land use and management is likely to have a major impact on deeper soil layers [FAO 2017b].
Indeed, significant amounts of carbon sequestration occurs in deeper soil profiles – even beyond a 1 m (3 ft) depth [Follett 2012, Liebig 2008, Schmidt 2011: 51]. Harper et al. found that half to three-quarters of total SOC to bedrock was in the surface 5 m with the remainder below that depth. The authors speculate that deep carbon may have been deposited directly by deep-rooting plants. “Where deep soils coincide with deep rooting the biological deposition of carbon from roots (and their associated biota) is inevitable at depths at which SOC has rarely been measured” [Harper 2013: 642].
Second, many studies measure sequestration rates for just one or two soil-building techniques, isolating them from additional, potentially synergistic, practices. In fact, intact ecosystems are based on countless synergistic relationships among organisms and their environment. In other words, many studies measure minor tweaks to conventional, industrial cropping systems.
For example, Minasny et al. [2017] compiled sequestration rates from around the world to assess the viability of the France-led “4 per 1000” initiative (seeking to halt the annual increase in atmospheric CO2 by increasing soil carbon by 0.4% per year). The authors estimate “that an annual rate of 0.2-0.5t C/ha/yr is possible after adoption of best management practices such as reduced tillage in combination with leguminous cover crops.”However, most of some 40 studies of best management practices on arable land assessed only one or two of many – often minimally improved – practices, such as “reduced use of summer fallow,” “rice-rice with NPK,” “inorganic fertilizer,” and “pasture.”
Similarly, an often-cited study by West and Post [2002], compiling 276 paired treatments from 67 long-term experiments, analyzes the sequestration rates for either increased rotation complexity (0.2+/-0.12t C/ha/yr) or a change from conventional tillage to no-till (0.57+/-0.14t C/ha/yr). While both practices were used at some sites, the data were not analyzed according to whether a single practice or combined practices were used. However, the authors suggest that using both practices together can be additive:
Data used in this analysis was stratified separately with regard to a change in tillage or a change in crop rotation. In practice, these changes could occur simultaneously. It can be inferred from our results that if of a decrease in tillage and an enhancement in rotation complexity occur simultaneously, the short-term (15–20yr) increase in SOC will primarily be caused by the change in tillage and subsequent decrease in the rate of SOC decomposition, while the long-term (40–60yr) increase in SOC will be primarily caused by the rotation enhancement and residue input and composition [West & Post 2002: 1943].
If moving to a combination of two restorative practices can increase carbon sequestration somewhat above the use of a single improved practice, then what is possible when many restorative practices are stacked one upon another within an agroecosystem? Permaculture, a design framework with “two broad conceptual criteria: ecosystem mimicry and system optimization,” where multiple restorative practices are indeed combined, represents a counterexample to industrial farming. Yet, sequestration rates from such a system have rarely, if ever, been measured. In fact, very little scientific study of any kind has been conducted in permaculture systems, despite the concept having been developed by scientist Bill Mollison, and adopted to favorable results globally for over 30 years [Ferguson and Lovell 2013].
In light of the centrality of agriculture’s role in ecosystem restoration due to the vast surface area it occupies, we present some literature representing agriculture’s maximum potential contribution to climate change mitigation, and argue that the focus of future research do the same.
Specifically, future studies should consider the effects of greater ecological intensity, diversity and potentially additive and synergistic interactions that can exist among multiple soil-building practices, rather than continuing to pursue measurement of their individual effects, which yield minimal outcomes. Future research must also measure SOC changes to greater depths in the soil horizon in order to capture the full benefit of any given practices. Such changes would likely present both a more accurate and more promising real-world potential for the climate mitigation potential of agriculture.
For a relevant and effective comprehensive assessment of regenerative management practices, one that supports the profound shift necessary in conventional 21st century agriculture, a scientific paradigm shift is necessary so that economics and policy will follow.
For 10,000 years, humans have been clearing patches of forest and grassland to plant crops. While clearing land by burning it visibly turns organic carbon into smoky CO2, plowing and tilling releases soil organic carbon by breaking up soil aggregates that protect carbon. Exposed soil organic carbon is consumed by microbes, and converted to CO2 through respiration. Tilling soil also subjects it to erosion. “Since tillage-based farming began, most agricultural soils have lost 30% to 75% of their soil organic carbon (SOC), with industrial agriculture accelerating these.” [Teague 2016: 157]
Agriculture is a source not only of CO2 emissions, but also of greenhouse gas emissions of methane (CH4) and nitrous oxide (N2O). In cropland soils, CH4 is produced by anaerobic decomposition of organic matter, usually in waterlogged soils like rice paddies. However, soils can also be a methane sink due to the presence of methanotrophic bacteria, which oxidize methane. N2O is produced by microorganisms, which transform excess ammonia fertilizer into nitrate and then N2O. “Upon conversion to NO2– or NO3–, excessive fertilizer N becomes subject to denitrification and thereby contributes to terrestrial emissions of N2O, which have been found to increase with the rate of N fertilization” [Mulvaney 2009: 2296].
Industrial agriculture compensates for soil carbon loss by abandoning degraded land or using chemical inputs for the nutrients and pest resistance that an otherwise carbon-rich, biologically active soil provides. However, the farming methods that rebuild topsoil without relying on synthetic inputs, while also ameliorating the worst effects of drought, are the same ones that can make agriculture a major sink for atmospheric CO2. Such methods, which can be used together as a complementary suite of practices include: no-till; cover-cropping; agro-forestry; diverse crop rotations, including integrating livestock grazing; use of compost, manure, and biochar; and use of deeper-rooting plants and perennials.
Cropland Article Summaries
Cover crops
Cover crops protect the soil during a time of year when no cash crops are growing and the soil would otherwise be bare. “Cover crops, also named inter-crops or catch crops, are crops that replace bare fallow during winter period and are ploughed under as green manure before sowing of the next main crop.” [Poeplau 2015: 34] Cover crops can also be rolled and crimped or mowed, instead of plowed, in preparation for the main crop.
Using cover crops reduces erosion, nutrient leaching, and drought stress, and add carbon through continued plant cover and growth as well as increase biodiversity. Leguminous cover crops also fix nitrogen. Furthermore, “in contrast to other organic amendments, a large part of the C input from cover crop is added as roots, which was found to contribute more effectively to the relatively stable carbon pool than aboveground C-input” [Poeplau 2015: 38].
Vick 2016. This Montana study demonstrates that leaving farmland fallow “depletes carbon stocks and thereby soil quality” [Vick 2016: 129], thus illustrating the importance of keeping land continuously covered with living vegetation. “Fallow” is the stage of crop rotation where no crop is grown.
In this study, a CO2 emissions rate of 1.35 tC/ha/yr (0.54 tC/ac/yr) was measured from land left fallow during the 2014 summer growing season; an adjacent field planted in winter wheat (summer 2013) and spring wheat (summer 2014) was a net carbon sink, measuring carbon input from the atmosphere into the soil at ~2 tC/ha/yr (0.8 tC/ac/yr) and ~1 tC/ha/yr (0.4 tC/ac/yr), respectively. Other parts of this study show a dramatic effect on area cooling as well as increased moisture and rainfall. These results occur only from ending the practice of fallowing.
The researchers observed that a widespread decline of land left fallow in agricultural areas of the Canadian Prairie Provinces coincided with a summertime cooling trend since the 1970s. They noted that extreme temperature events now occur less frequently than in the recent past, maximum summer temperatures have decreased by ca. 2° C (3.6° F), relative humidity has increased by some 7% and summer precipitation has increased by an average of 10 mm/ decade across parts of the Canadian Prairie Provinces. A remarkable 6 W/m2 summer cooling has been observed compared to a ca. 2.5 W/m2 warming globally since the dawn of the Industrial Era.
Even in degraded croplands, relatively small changes can lead to significant differences in rainfall, soil carbon sequestration, and ambient temperature. A 2016 study in Montana demonstrated the effects of reversing the practice of fallowing of wheat fields in the upper midwest. Fallowing is “the practice of keeping a field out of production during the growing season” (Vick 2016:129):
Fallow is a common management practice in the dryland wheat-growing regions of the northern North American Great Plains to conserve water for subsequent crops (Lubowski et al., 2006). Fallow however also increases erosion (Wischmeier, 1959) and soil carbon loss (Cihacek and Ulmer, 1995), and fallow-small grain management strategies are not considered sustainable from the soil conservation perspective (Merrill et al., 1999). [Vick 2016:130].
As a result of farmers’ experience, fallowing has progressively decreased across many areas of the northern midwestern plains since the 1970s, providing an environment suitable for comparison study:
The area of fallow in the Prairie Provinces of Canada has decreased from over 15 Mha in the 1970s to under 2 million ha at the present (Fig. 1) as producers have realized that the water-savings benefit of fallow is outweighed by the economic losses of not planting (Dhuyvetter et al., 1996). The area under fallow in the United States has likewise decreased from 16 Mha to 6 Mha across the same time frame (Lubowski et al., 2006), largely in the northern Great Plains and other areas of the semiarid West . . . Despite the decreasing trend in fallow area across the North American northern Great Plains, fallow remains common in many regions including major land resource area
(MLRA) 52 in north-central Montana – the largest wheat-growing region in the state – where some 40% of agricultural lands may remain in fallow in any given year. In contrast, fallow has been reduced in northeastern Montana (MLRA 53) by hundreds of kha over the past decade (Long et al., 2014, 2013) as producers have adopted continuous cropping or alternate cropping practices. [Vick 2016:130].
The effects of this relatively simple change of practice led to some remarkable results:
The widespread decline of fallow in agricultural areas of the Canadian Prairie Provinces (Fig. 1) has coincided with a summertime cooling trend since the 1970s (Betts et al., 2013a, 2013b; Gameda et al., 2007; Mahmood et al., 2014). Extreme temperature events now occur less frequently than in the recent past, maximum summer temperatures have decreased by ca. 2০C, relative humidity has increased by some 7% (Betts et al., 2013b), and summer precipitation has increased by an average of 10 mm/decade across parts of the Canadian Prairie Provinces (Gameda et al., 2007). A remarkable 6 W/m2 summer cooling has been observed (Betts et al., 2013a); for reference, anthropogenic greenhouse gasses are responsible for a ca. 2.5 W/m2 warming globally since the dawn of the Industrial Era (IPCC, 2007). These climate benefits have only occurred during the growing season; fall, winter, and early spring temperatures have followed global trends (Betts et al., 2013b) . . . In other words, the observed regional climate cooling is broadly consistent with the effects of fallow avoidance on climate processes. [Vick 2016:130-131]
As dramatic as some of these changes are with only reduced fallowing, there are other land-management practices with significant impacts on water cycles, soil carbon, biodiversity and productivity. Such practices hold additional potential, and include cover-cropping and green mulches, pasture cropping, elimination of synthetic inputs which encourage renewed activity of important soil biota, especially worms – and perhaps most importantly on grasslands that co-evolved with animals, the reintroduction of animals themselves.
Finally, it is worth noting that there may be a significant underestimation of surface area and volume of soils in grasslands, as well as in other ecosystems, since natural topographies are not uniformly flat. Topographical variations would add volumes of soil carbon, water, etc. to prior estimates of areas that are typically calculated on the basis of two-dimensional map projections [Blakemore 2016: Fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regenerative management, carbon capture and water storage than is conventionally assumed. Such adjustments to soil volume calculations would positively affect carbon drawdown estimates in considering the potentials of eco-restoration in climate (see section, Do We Have More Soil for Carbon Storage than We Thought?).
Pimentel 2011. Arguing for cover crops as an effective way to reduce erosion and conserve nutrients in soil, Pimentel notes that “Growing cover crops on land before and after a primary crop nearly doubles the quantity of solar energy harvested in the agricultural system per hectare per year. This increased solar energy capture provides additional organic matter, which improves soil quality and productivity.” [Pimentel 2011: 41]
Crop rotation
Crop rotation diversification can enhance pest resistance, nitrogen input (when leguminous crops are added), soil penetration for better water infiltration (when deeper rooting plants are added), and residue input (when crops that produce more biomass are added). The effects on carbon sequestration from increases in crop rotation diversity vary depending on what crops are included. “Crop species can vary significantly in growth patterns, biomass production, water requirements, and decomposition rates, all of which affect net GHG emissions. Therefore, many rotations could be adapted with alternative species or varieties of annual crops to promote soil C sequestration—increasing root and residue biomass, increasing root exudates, or slowing decomposition—or otherwise reduce emissions” [Eagle 2012: 13].
Clearly, crop rotation is something of an umbrella term, describing a variety of practices, and even leaving space for practices that would not seem to offer much in the way of soil restoration. For example, as West and Post [2002] state,
. . . enhancement of rotation complexity refers to (i) a change from monoculture to continuous rotation cropping, (ii) a change from crop–fallow systems to continuous monoculture or rotation cropping, and (iii) an increase in the number of crops used in a rotation cropping system. In this analysis, continuous cropping is a cropping system without a fallow season, monoculture is a system with only one crop grown, and rotation cropping indicates two or more crops rotated over time on the same unit of land. [West & Post 2002: 1931]
Thus, even “continuous monoculture” can be considered as a crop rotation meant to increase carbon sequestration capacity. On the other hand, crop rotation can also involve great diversity, such as at Paul and Elizabeth Kaiser’s farm, where 3-7 crops/year rotate through vegetable beds, sometimes intercropped two crops at a time [Kaiser 2017].
Teague 2016. This study argues for greater use of no-till, cover crops, and crop rotation, including integrating livestock rotation into cropping systems.
Crop production can be managed to maintain permanent ground cover through the rotation of forage and row crop mixes, including cover crops, and legumes to increase soil fertility by fixing N. Grazing livestock can accelerate nutrient cycling through the consumption and decomposition of residual aboveground biomass.” [Teague 2016: 159]
The authors present a set of testable hypothetical scenarios suggesting the adoption of conservation cropping and adaptive management grazing (including grass-finishing cattle).
No-till
No-till (NT) allows farmers to plant without disturbing the soil, thus protecting it from water and wind erosion, leaving soil aggregates intact, and preventing a flush of oxygen from activating microbial breakdown of organic matter and releasing CO2. No-till can contribute to climate mitigation both by reducing emissions from the turnover of soil organic matter caused by tillage, and by sequestering carbon, especially in the surface layer [Mangalassery 2015].
Brown 2016. North Dakota Farmer Gabe Brown began practicing no-till in 1994. Since then, he has added cover crops (a diverse mixture of 70 species), complex crop rotations, orchards, livestock grazing (including cattle, sheep, pork and chicken), vegetable production, and bees. Through a long-term commitment to building the soil through no-till, keeping the ground always covered, and favoring as much biodiversity as possible (including a wide diversity of cash crops), Brown reports SOM has increased from 1.7% in 1993 to 11% in 2013. Furthermore, water infiltration has increased from ½ inch to more than 14 inches over the same time span.
Follett 2012. Measured to a depth of 150 cm (~5 ft), no-till continuous maize grown in eastern Nebraska, fertilized with 120 kg/ha of nitrogen and stover left on the field after grain harvest, sequestered 2.6 tC/ha/yr (1 tC/ac/yr). Notably, more than 50% of sequestered carbon was found below 30 cm (1 ft), illustrating that studies failing to sample below this depth (a common practice) risk greatly underestimating sequestration rates.
Organic vs. synthetic inputs
Organic farming uses “cultural, biological, and mechanical practices that support the cycling of on-farm resources, promote ecological balance, and conserve biodiversity” according to the USDA, which prohibits the use of most synthetic pesticides and fertilizers on certified organic farms. Organic farmers must find alternatives to synthetic inputs for managing pests and fertility. For example, vermi-composting is commonly used in organic farming. It is a natural and proven enhancement of the humification process that uses specific earthworms (e.g. Eudrilus eugeniae [Blakemore 2015]) to rapidly convert all organic “wastes.” Returning this vermicompost to soil renders synthetic fertilizers and pesticides unnecessary, as vermicompost often confers natural resistance to pests [Howard 1945, Balfour 1975] and it enhances resident earthworms [Blakemore 2000, 2016a; see also Earthworms section].
While the organic law provides a baseline for organic practices, the term “organic” encompasses a wide range of approaches to farming. For instance, some organic farmers may do little more than substitute naturally occurring inputs into an otherwise conventional, industrial operation, likely leaving the soil similarly depleted. Other organic farmers put into practice several of the methods mentioned in this section, aiming to truly build the functionality of the soil to resist pests and provide fertility. The studies included below highlight benefits from organic inputs and problems that come with using synthetic fertilizers with respect to soil carbon and biodiversity.
Johnson 2017. Using fungal-dominant compost in a 4.5-year trial at Leyendecker Field Research Site in New Mexico, researchers recorded an annual carbon sequestration rate of 10.7t C/ha/yr (4.8t C/ac/yr). Based on the observed trajectory of increasing productivity, they estimate a potential rate of 19.2t C/ha/yr (7.67t C/ac/yr). Chief investigator David Johnson found that increased plant growth is correlated most closely with the fungal to bacterial ratio. At a fungi:bacteria ratio of 0.04, only 3% of carbon flow went into plant biomass production, with the remainder of the carbon going into other functions, including nitrogen fixation, the soil, and respiration. At a fungi:bacteria ratio of 3.68, plant growth was more efficient with 56% of carbon flow going to biomass production.
Rodale 2014. Compiling data collected from around the world, Rodale Institute concluded that if all cropland were converted to their regenerative model[17], it would sequester 40% of annual CO2 emissions. Adding pastures to that model would add another 71%, effectively exceeding the world’s yearly carbon dioxide emissions.
On-farm soil carbon sequestration can potentially sequester all of our current annual global greenhouse gas emissions of roughly 52 gigatonnes of carbon dioxide equivalent (GtCO2e). Indeed, if sequestration rates attained by exemplar cases were achieved on crop and pastureland across the globe, regenerative agriculture could sequester more than our current annual carbon dioxide (CO2) emissions. Even if modest assumptions about soil’s carbon sequestration potential are made, regenerative agriculture can easily keep annual emissions within the desirable lower end of the 41-47 GtCO2e range by 2020, which is identified as necessary if we are to have a good chance of limiting warming to 1.5°C. [p.5]
Ryals and Silver 2013. This study examined the effects on plant growth and respiration from compost application on annual grassland in both coastal and valley sites in California. They found that a single application of compost during the three-year study resulted in a carbon sequestration rate of 1.45t C/ha/yr (0.58t C/ac/yr) and 0.54t C/ha/yr (0.22t C/ac/yr) at the valley grassland and coastal grassland, respectively. This enhanced net primary productivity was partially offset by CO2 emissions from increased respiration, but the compost did not affect CH4 or N2O fluxes. The authors conclude that:
Our results have important implications for rangeland management in the context of climate change mitigation. Urban and agricultural green waste is often an important source of greenhouse gas emissions (IPCC 2001). Here we show that an alternative fate for that material can significantly increase NPP and slow rates of ecosystem C losses at the field scale. This approach provides important co-benefits to landowners, such as the sustained increase in forage production measured here [Ryals & Silver 2013: 56].
While these results are low compared to some of the other studies noted here, this study illustrates positive use for green waste, and a potential tool that may contribute to climate-positive management.
Khan 2007. This five-decade study of nitrogen fertilization effects on SOC in Illinois shows that, despite progressively greater corn crop residue input during the second half of the 20th Century (increasing from 20,000 or 30,000 to 69,000 plants/ha since 1955), partly due to synthetic fertilizer use, SOC content did not increase, and in most cases declined. SOC declines were most pronounced in subsurface (16-46cm) of the soil horizon, compared to the surface layer (0-15cm). These results are despite crop residue being incorporated, rather than removed, in most plots since 1955, and in all plots since 1967.
These findings implicate fertilizer N in promoting the decomposition of crop residues and soil organic matter and are consistent with data from numerous cropping experiments involving synthetic N fertilization in the USA Corn Belt and elsewhere, although not with the interpretation usually provided. [Khan 2007: 1821]
Perennial systems, agroforestry, and permaculture
Unlike annual plants, perennials live for many years – thousands of years in some cases. Because of their deep (>2m, or 6 ft) and extensive root system, and longer growing seasons, perennials are likely to sequester carbon better than annual cropping systems [Glover 2007].
Agroforestry is the practice of integrating trees (a type of perennial) into a cropping system, including alley cropping, windbreaks, riparian buffers, silvopasture, and forest farming [Eagle 2012; Nair 2009]. Agronomic practices are notable for adding significant amounts of carbon to aboveground biomass, which is often measured separately from soil organic carbon sequestration [Nair 2009]. One of the strengths of agroforestry is its enhancement of an agroecosystem’s functional diversity:
The utilization of the environment by species includes three main components: space, resources, and time. Any species utilizing the same exact combination of these resources as another will be in direct competition which could lead to a reduction in C sequestration. However, if one species differs in utilization of even one of the components, for example light saturation of C3 vs. C4 plants,[18]C sequestration will be enhanced.” [Udawatta 2011: 19]
Toensmeier 2017. Compiling carbon sequestration rates from individual studies, reviews, and expert estimates, and organizing them into groups of annual versus perennial systems, woody versus herbaceous crops, and polyculture versus monoculture, Toensmeier observes that “the general trend is that systems that incorporate trees sequester more carbon.” The highest sequestration rate listed, 18 tC/ha/yr (7.2 tC/ac/yr) falls into the perennial woody polyculture group, and more than half of all sequestration rates listed under perennials are more than 6 tC/ha/year (2.4 tC/ac/yr), while most rates for annual cropping systems are less than 1t C/ha/yr (0.4 tC/ac/yr).
Lawton 2016. On 10 acres of the Arabian Desert in Wadi Rum, Jordan, Permaculture Designer Geoff Lawton built an organic, multi-species food forest on what had previously been bare desert ground. Using wastewater from a nearby irrigated farm to get started, he designed a microclimate that would prevent evaporation in every way possible. Key elements included: date palm trees for wind protection and shade; smaller fruit tree and trellised grapevines for additional shade; a succulent ground cover, which also catches nutrient-rich desert dust; a shaded swale for irrigation; and cut legume trees for mulch.
Lawton sought to “build organic matter within the system as quickly as possible with any living elements that will achieve those ends.” Once the soil came alive, it became productive. Lawton explains that strategic arrangement of the space is especially important in the desert. That’s why crops were grown in two rows in between three slightly wider rows of mixed fruit trees for protection. After four years, this orchard/farm was producing an abundance of fruits and vegetables, showing that it is possible to work with nature and avoid industrial inputs to achieve a productive landscape even in the harshest environment.
DuPont 2010. A Land Institute study measured the effect on soil properties and biota from perennial polyculture systems as compared to annual grain crop systems. Since the latter are typically intensively managed, “the effects of tillage and plant community composition are often confounded” [DuPont 2010: 25]. To control for management effects, this study compared the soil carbon and root biomass outcomes from no-tilled annual crops (rotation of soybean, sorghum and wheat) versus a perennial polyculture. Total root biomass in no-till annual plot measured at only 43% of that in a perennial grass plot in the top 1m of soil. Also, the authors found significantly higher levels of readily oxidizable carbon (ROC) and microbial biomass in the perennial plots compared to the annual crop plots. ROC measures soil carbon that is more available to soil microbes.
Small changes in ROC and other labile fractions of SOC may provide an early indication of soil degradation or improvement in response to management practices. Changes in active carbon pools can be two to four times greater than changes in total C after the initiation of new management practices and they are more highly correlated with other soil quality indicators including microbial respiration, aggregate stability and plant productivity [DuPont 2010: 28].
The authors conclude that “even in the absence of tillage and under best management practices, annual cropping can reduce soil carbon and impact soil biota and food webs important in nutrient cycling after just three years” [DuPont 2010: 25].
Soto-Pinto et al. 2009. In this southern Mexico study of land-use change in various agroforestry systems, the authors show that converting “traditional fallow” (secondary growth woods following cropping, averaging 23.4 years in age) to maize (with beans, squash and pepper) production results in 94% loss of living biomass carbon. However, transitioning to (a) “taungya” (maize, beans, squash and peppers intercropped between rows of timber and multipurpose trees), (b) shaded coffee systems, or (c) “improved fallow” (adding timber trees to traditional fallow plots) preserves living biomass carbon. This study points to the mounting relevance of agroforestry systems that can provide economic benefits to small-scale farmers, while avoiding carbon emissions from land use change from forest to agriculture and livestock production, which accounts for 35% of total emissions in Mexico, according to the authors.
Association for Temperate Agroforestry 2004:
Agroforestry practices are intentional combinations of trees with crops and/or livestock which involve intensive management of the interactions between the components as an integrated agroecosystem.
Intentional: Combinations of trees, crops and/or animals are intentionally designed and managed as a whole unit, rather than as individual elements which may occur in close proximity but are controlled separately.
Intensive: Agroforestry practices are intensively managed to maintain their productive and protective functions, and often involve annual operations such as cultivation, fertilization and irrigation.
Interactive: Agroforestry management seeks to actively manipulate the biological and physical interactions between the tree, crop and animal components. The goal is to enhance the production of more than one harvestable component at a time, while also providing conservation benefits such as non-point source water pollution control or wildlife habitat.
Integrated: The tree, crop and/or animal components are structurally and functionally combined into a single, integrated management unit. Integration may be horizontal or vertical, and above- or below-ground. Such integration utilizes more of the productive capacity of the land and helps to balance economic production with resource conservation.
Liebig 2008. Measured to a depth of 120 cm (~4 ft), switchgrass grown for bioenergy at 10 farms across the Great Plains in the United States sequestered 2.9 tC/ha/yr (1.16 tC/ac/yr). Of that, only 1.1 tC/ha/yr (0.44 tC/ac/yr) was found in the first 30 cm (1 ft) depth, with the remainder measured below 30 cm. The authors explain what makes switchgrass effective in carbon sequestration:
Increases in SOC [soil organic carbon] under switchgrass were likely caused by belowground C input from root biomass and rhizodeposition and decreased soil organic matter losses by erosion. Research conducted by ecologist John Weaver and his graduate students over 60 years ago provide ancillary support for increased SOC under switchgrass. Their detailed surveys of prairie grass roots indicated switchgrass to have the deepest root system of all grasses examined, with roots extending to a soil depth of 3m (~10 ft). This finding, coupled with observations that prairie grass roots regenerate by replacing dying roots with new, live roots indicates the potential for significant C input to the soil under switchgrass.
Montagnini & Nair 2004. Agroforestry systems are multifunctional with respect to carbon capture. Agroforestry can: increase the soil carbon content and fertility of cropland, while allowing for continued food production; create greater sequestration efficiency through diversity of vegetation; and allow for harvest of forest products, potentially keeping carbon sequestered in wood products for many years, and thereby also decreasing pressure on natural forests. And because of the mixed use of agroforestry systems:
[T]he amount of biomass and therefore carbon that is harvested and ‘exported’ from the system is relatively low in relation to the total productivity of the tree (as in the case of shaded perennial systems). Therefore, unlike in tree plantations and other monocultural systems, agroforestry seems to have a unique advantage in terms of C sequestration [Montanigni & Nair 2004: 285].
A few sequestration rates highlighted in this article include: A Costa Rica study of cacao grown under two different species of shade trees Erythrina (a leguminous tree) and Cordia (a timber tree), measured C sequestration in perennial plant biomass at an average of 4.28t C/ha/yr (1.7t C/ac/yr) for the cacao-Cordia system, and 3.08t C/ha/yr (1.2 tC/ac/yr) in the cacao-Erythrina system . In another study, tropical smallholder agroforestry was projected to sequester 1.5-3.5t C/ha/yr (0.6-1.4 tC/ac/yr).
Onim 1990. Tropical agroforestry was observed to increase SOC (soil organic carbon), at the 0-30 cm depth, to a maximum of 8.34 tC/ha/yr (3.38 tC/ac/yr)and minimum of 0.73 tC/ha/yr (0.30 tC/ac/yr).
Biochar
Biocharis organic matter that has been decomposed through pyrolysis (burning) under controlled, low-oxygen conditions, where it emits relatively little CO2. Biochar is then added to the soil for long-term carbon storage and/or enhancing availability of soil nutrients, oxygen and water to plants and microbes. Because charred biomass has been observed to persist in the soil for centuries or millennia, biochar is seen as a stable or recalcitrant form of carbon that that may prove to be a useful tool for reversing climate change. Not only is the biochar itself a stable form of carbon that can remain in soils long-term, but also it helps build healthy soil structure which increases plant growth and therefore photosynthetic capacity, resulting in carbon being removed from the atmosphere and stored in biomass or soils. [McLaughlin 2017; Taylor 2010; Paustian 2016; Weng 2017; Remediation Magazine 2017]
It is worth noting that depending on the pyrolysis technique, the resulting biochar may range in quality from poor to excellent. One hopes that as the industry matures, the understanding of the importance of biochar quality in assessing results will grow as well.
McLaughlin 2017. Hugh McLaughlin, Ph.D., P.E. is an expert on the properties and production of chars created by pyrolyzing biomass, and the subsequent conversion to activated carbons. He has published extensively on biochar and biomass-derived heat production. In this video he gives a short but comprehensive review of the qualities and use of biochar.
Paustian 2016. Biochar application to soils is considered in this article among several activities (such as compost application, cover cropping, residue retention, no-till, and others, as previously mentioned in this compendium) designed to increase soil C stocks by increasing organic matter inputs or reducing decomposition rates. Biochar acts as a soil amendment stimulating plant growth, thereby allowing for greater C storage through greater biomass production, while also embodying a generally stable form of buried carbon.
Biochar mineralizes 10–100 times more slowly than uncharred biomass. Thus a large fraction of added C … can be retained in the soil over several decades or longer, although residence times vary depending on the amendment type, nutrient content and soil conditions (such as moisture, temperature and texture).
However, because the organic matter originates from outside the ecosystem ‘boundary’, a broader life-cycle assessment approach is needed, that considers the GHG impacts of: (1) offsite biomass removal, transport, and processing; (2) alternative end uses of the biomass; (3) interactions with other soil GHG-producing processes; and (4) synergies between these soil amendments and the fixation and retention of in situ plant-derived C. In many cases, net life-cycle emissions will largely depend on whether the biomass used as a soil amendment would have otherwise been burnt (either for fuel, thereby offsetting fossil fuel use, or as waste disposal), added to a landfill, or left in place as living biomass or detritus [Paustian 2016: 50].
Remediation Magazine 2017. A popular report on Weng 2017, quoting the authors:
The project’s leader, DPI [Department of Primary Industries] researcher and SCU [Southern Cross University] adjunct professor Lukas Van Zwieten said the research threw up some unexpected results. “We immediately saw an increase in soil carbon from the biochar, as expected, but what we didn’t expect was that soil carbon content continued to increase. This research demonstrates the ongoing benefits of biochar in farming systems to improve pastures and grasslands and increase farmers’ production and profitability . . . the researchers found that biochar enhanced the below-ground recovery of new root-derived carbon by 20% – that is, more of the carbon photosynthesised by plants was retained in the biochar-amended soil. Biochar accelerated the formation of soil microaggregates via interactions between organic matter and soil minerals, thus stabilising the root-derived carbon. . . . The increased microbial activity and improved physical structure of the soil would also ultimately improve the effectiveness of fertiliser use, making the application of biochar particularly beneficial for high-end, intensive crop production”
“[T]he improved structure of the soil protected the naturally occurring carbon, as well as the carbon added”, said Southern Cross University’s associate professor Terry Rose, a co-author of the study. “Importantly, the biochar also slowed down the natural breakdown of native soil organic carbon by more than 5%.
Taylor 2010. An anthology of articles written by biochar pioneers. Covers biochar history, testing, production, challenges and uses. Suitable reading for general audiences as well as land management and industry professionals.
Weng 2017. Biochar can increase the stable C content of soil. However, studies on the longer-term role of plant–soil–biochar interactions and the consequent changes to native soil organic carbon (SOC) are lacking. . . . We found that biochar accelerates the formation of microaggregates via organo-mineral interactions, resulting in the stabilization and accumulation of SOC in a rhodic ferralsol (s.a. Remediation Magazine 2017).
Grasslands
Grasslands have been estimated to cover approximately 40% of global land surface area, approximately 5.25 bn ha (~13 bn ac ), except for Greenland and Antarctica [Suttie 2005; White 2000:12]. Their deep soils are rich repositories of nutrients, especially carbon and water. Many grasslands are anthropogenic, i.e., resulting from various land management techniques to maintain land for grazing and crop production by humans. Virgin grasslands are increasingly rare, possibly leading to significant underestimations of their potential positive contribution to productivity, and to carbon and water storage. Grasslands are important repositories of biodiversity, and have significant impacts on weather and climate. Here we review research and articles that indicate soil carbon storage potentials of roughly 13 gigatons per year (the equivalent of 6.5 ppm) if global grasslands were managed regeneratively.
Overview
While we have separate sections for Grasslands, Croplands and Soils, there is inevitable overlap. Many croplands are modified grasslands, and both are, of course, based in soils. Yet there are enough differences in each area of study to merit separate sections, keeping in mind that systemic behaviors and interactions are broadly applicable.
Since the onset of agriculture over 10,000 years ago with land management techniques that expose soil to air, estimates of up to 537 gigatons of soil carbon have been oxidized to carbon dioxide and other greenhouse gases [Buringh 1984: 91]. Even so, soils (>2,300 Gt) currently hold almost as much carbon as plants (550 Gt), atmosphere (800 Gt) and ocean surface waters (1,000 Gt) combined [NASA 2011], and almost surely retain the potential to store enough atmospheric carbon to return to pre-industrial levels.
Typical soil studies examine the first 30 cm (1 ft) of soil depth, but more recent investigations indicate that major soil carbon storage takes place deeper than that, often in a more stable form [Liebig 2008, Follett 2012, Harper 2013]. A USDA paper found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr) [Follett 2012]. A study of switchgrass for bioenergy found rates of SOC increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft) [Liebig 2008]. On an intensively grazed former row-crop agriculture land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions [Machmuller 2015]. In addition, the ultimate methane emissions may have been markedly less than measured since the report did not consider methane breakdown into CO2 from methanotrophic bacteria and atmospheric hydroxyl radical oxidation, with a significant reduction of methane’s ultimate greenhouse gas impacts.
These reports demonstrate the potential for massive amounts of soil carbon storage, significant cooling of the biosphere, and dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Grassland Evolution
Grasslands have long been a rich repository of carbon, both stable and labile. The co-evolution of grasslands with grazing ruminants has contributed to dramatic global cooling over the past 50 million years as a result of significant photosynthetic carbon drawdown into grassland soils [Retallack 2013]. Thus, grasslands are more than a consequence of geophysical changes, they are
. . . a biological force in their own right (Retallack 1998), in some ways comparable to the human rise to dominance of planetary resources (Vitousek et al. 1997). Grasslands have long been considered products of the coevolution of grasses and grazers (Kovalevsky 1873). Few plants other than grasses can withstand the high-crowned, enamel-edged teeth and hard hooves of antelope and horses. Yet these same animals are best suited to the abrasive gritty opal phytoliths and dust of flat, open grasslands. Grasses recover readily from fire and nurture large herbivores such as elephants: both fire and elephants promote grassland at the expense of wood land (Retallack 1997b; Jacobs et al. 1999). Grasses suppress insect and fungal attack with secondary metabolites such as cyclic hydroxamic acids (Frey et al. 1997). Grasses create Mollisols, unique soils with fine crumb clods rich in organic matter (Retallack 1997b). (Retallack 2001:407) [Emphasis added.]
Occupying such vast areas of planetary land surface, grasslands have a major influence on the global climate:
CO2 and CH4 (which rapidly oxidizes to CO2 ) are important greenhouse gases, and mechanisms for burial of their C may result in climatic cooling (Berner 1999). The most important long-term C sink from grasslands is their supply by erosion to sedimentary basins of crumb peds, which are unusually rich in organic matter intimately admixed with clay (Pawluk and Bal 1985). Tropical forests, in contrast, yield highly oxidized spherical micropeds with virtually no organic content (Retallack 1991a).” (Retallack 2001:414)
While there are other soils that are greater carbon sinks, such as peat bogs, wetlands and coastal habitats (e.g., mangroves, seagrasses), for volume and depth of carbon storage on vast areas of land, grasslands have enormous potential:
Grasses themselves are C sinks, especially considering their mass of roots and rhizomes underground. . . . [Numerous soil investigations] indicate that past estimates of organic C in tropical grassland soils have been low, in part because soils were not analyzed to sufficient depths. . . . Grassland and woodland soils may have comparable amounts of organic C in the surface 15cm. Beyond that depth, organic C values drop off dramatically in woodland soils but remain high in grassland soils to a meter or more. The fine structure and fertility of grassland soils is in large part due to this large C reservoir. (Retallack 2001:415)
Conventionally, it is estimated that approximately 40% of global land surface area is grasslands (52.5 million square kilometers, or ~5.25 billion hectares, or ~13 billion acres [Suttie 2005]), except for Greenland and Antarctica [White 2000:12; see Figure 1, below]. This is likely a significant underestimation of soil surface area and volume, since grasslands are not uniformly flat, with topographical variations adding carbon, water, etc. to areas that are typically calculated on the basis of a two-dimensional map projection [Blakemore 2016: fig. 5]. The implications are that there may be considerably greater volumes of soil amenable to regeneration, carbon capture and water storage than is conventionally assumed (see the section, “Do We Have More Soil for Carbon Storage than We Thought?”).
Natural grasslands are typically areas of low and seasonal rainfall. Unlike temperate environments with year-round rainfall, semi-arid and arid grasslands are dependent on grazing animals as a keystone species. The habits of grassland plants are as dependent on grazing animals as the animals are dependent on the plants that grasslands provide as food.
Figure 1: Global Extent of Grassland (White 2000:12).
Grazing animals are ruminants and as long as they are grazed in herds that move frequently, as they do in natural habitats in the company of predators, ruminant species are often interchangeable insofar as grassland health is concerned. Bison and antelope, for example, may be the wild ruminants that roamed the prairies and savannahs, but domestic cattle, when properly managed (human herders are the equivalent of wild predators), serve the same ecosystem functions.
Pioneering Work Of Allan Savory
Allan Savory, a wildlife biologist from Zimbabwe, began studying desertification in the 1950s. He pioneered an approach that he has termed Holistic Planned Grazing (HPG) for regenerative management of grasslands. He noted that there are essential differences between temperate grasslands, which he termed “non-brittle” environments, and arid and semi-arid grasslands, which he referred to as “brittle.” These distinctions are critical in understanding how different habitats require different management approaches.
Non-brittle environments, because of year-round rainfall, are relatively forgiving of mismanagement that destroys soil biota and exposes soil to sunlight, air and the elements. Recovery from soil degradation can be be rapid. Brittle environments, to the contrary, are fragile and easily desiccated, and when poorly managed, either from overgrazing or undergrazing, may take decades or centuries to recover or even ultimately turn to desert.
Brittle environments particularly need ruminant evolutionary partners. Their hooves are designed to open the soils to air and water, and their digestive systems deposit a feast for soil organisms. The ruminant gut is a moist refuge during the dry season for soil microbes which are essential to the health of the land. Grasses need to be bitten lest they shade out their own new growth.
What Savory discovered is that the same land may either flourish or die depending on how it is grazed. When ruminants are kept in check by predatorsthey graze an area in tight herds for protection and then move to the next patch of fresh grasses and other plants, providing up to two years of recovery and regrowth time for the recently grazed pasture. On the other hand, when they are provided the safety of fencing and left to graze large areas at will, they return to their favorite plants and overgraze those areas, eventually compacting the soil, preventing water infiltration and proper aeration, killing the plants, and leading to desertification.
The difference in land health is dramatic.[19] The pictures below illustrate:
Mexico
Arizona
Zimbabwe
Fig. 1. These pictures are of neighboring properties in Mexico, Arizona and Zimbabwe. In each area they were taken on the same day, have similar soils, and the same precipitation. The pictures on the right are examples of properly managed livestock through Holistic Planned Grazing to restore grasslands. On the left we see examples of improperly managed livestock as well as exclusion from grazing (“resting the land”) [Savory Institute 2015:12]
Savory’s work, after decades of successful application on ranches in Africa, Asia, Australia and North and South America, garnered global attention (and controversy) after his TED Talk in 2013 [Savory 2013].
The primary point is that If well-managed, grassland soils can not only sequester annual greenhouse gas emissions but can alsobegin to draw down legacy atmospheric carbon as well. In addition, they provide human and other predator food, converting grasses inedible to non-ruminant mammals to meat.
Grasslands As Ecosystems
It is useful to understand how grasslands work as intact ecosystems, thereby providing a solid theoretical basis for observations of grasslands as vast carbon and water sinks.What appears above-ground is only a hint of grassland ecosystem dynamics. Soils are the planet’s most complex and least understood terrestrial ecosystem, yet soils are where most of the action takes place on grasslands as well. While all soils are built on the foundation of minerals provided by weathering of rock, the soils on grasslands are primarily biological soils. The kingdoms of life are the active agents in soil creation and it is the interactions among life forms that create the rich and productive grassland soils (see Soils section).
The basis for all life is the microbial kingdom. These smallest of cells, with their complex biochemistry, morphology and behavior, are active players in creating stable soil molecules, storing abundant carbon and water. In addition to microbes, soil ecosystems are built from exchanges among fungi, insects, worms, green plants, birds and small and large mammals. It is this set of rich interactions that creates the biodiverse, abundant and resilient environment of global grasslands.
For the sake of illustration, let us start the discussion with the ruminant gut during a dry season. Microbes survive in a warm, moist environment while constantly being cycled into the soil through digestion and elimination. The short-term hoof disturbance with minimal compaction while animals are constantly moving opens the soils to available moisture from precipitation, urination and condensation. During the rainy season the water is more effectively absorbed into opened soils, nourishing plants, raising the water table and eventually even leading to perennial streams and ponds. Even limited rainfall goes a long way in spongy soils that are covered with grasses and other plants to keep the ground cool and moist [Byck 2014: 8’38”]
Methane
Methane, a relatively short-lived but powerful greenhouse gas, is often raised as a serious concern with beef production. This is surely true when animals are left to roam freely and overgraze, and then moved to concentrated animal feeding operations with large manure lagoons. However, it is important to consider the whole of ecosystem functions in assessing methane emissions {Savory Institute 2015]. This includes the conversion of the methane molecule into carbon dioxide by bacteria (metanotrophs) that live in healthy soils and literally eat energy-rich methane, and the oxidation of methane by hydroxyl radicals present in the lower atmosphere. It may include other ecosystem processes, such as the effects of earthworms increasing methanotrophic bacterial activity in landfills and pastures [Héry 2008; Kernecker 2014]. The result is a virtuous cycle where plants can then take up that carbon dioxide through photosynthesis and send some of the carbon back underground through their root systems. Throughout their life cycle under conventional industrial management, cattle are rarely if ever exposed to such healthy, biodiverse soils.
Historical methane data indicates that in the United States, for example, pre-settlement wild ruminants generated approximately 86% of the methane of current farmed ruminants (Hristov 2012:1371). Yet even with vast numbers of ruminants on grasslands across the planet, atmospheric methane remained constant until the global dependence on widespread use of fossil fuels and its effects on agriculture and animal husbandry began to grow rapidly in the 18th century (Fig. 2).
Thus, results are very different with animals grazed in a manner that mimics nature. Rowntree et al. describe the importance of accounting for the beneficial ecosystem services that well-managed grazing systems can provide.
. . . LCA’s [Life Cycle Assessments] often consider soil C to be in dynamic equilibrium. However, empirical data suggest otherwise (e.g. Machmuller et al., 2015; Teague et al., 2011). Recent studies such as Ripple et al. (2014) and Eshel et al. (2014) have reported the emissions from ruminants in food production without accounting for the beneficial ecosystem services that well-managed grazing systems can provide. In our study, we used 3 tC/ha/yr (1.2 tC/ac/yr) as a potential C sequestration figure, which is relatively high (Conant et al., 2001) but viable based on existing studies (Teague et al., 2011; Delgado et al., 2011; Machmuller et al., 2015; Teague et al., 2016). Importantly, the results presented here suggest that with appropriately managed grazing, a grass-finished beef model can not only contribute to food provisioning but also be ecologically regenerative as well. [Rowntree 2016:36]
This excerpt illustrates a paradigm shift in action, wherein investigators within the dominant paradigm are constrained from evaluating the possibilities offered by “beneficial ecosystem services” because it doesn’t occur to them to consider them. Such biological processes are invisible due to limiting assumptions of the paradigm.
Research into systems implications of holistically managed grass-finished beef is growing. It is only recently that mainstream researchers are beginning to understand that the biological function of animals in an ecosystem is as dependent on the ecosystem as it is on the biology of the animal.[20] Studying animals in isolation or as part of a synthetic system such as industrial agriculture often leads to incorrect conclusions.
Fig. 2. Historical Concentrations of Greenhouse Gases [IPPC 2007]
Review of some studies of grasslands
Soil creation (pedogenesis) is conventionally defined as the weathering of rock; it may take 3,000 years or longer to create a foot of soil through geological processes. Soil created through biological activity, on the other hand, happens orders of magnitude faster, up to several inches per year.
Australian soil scientist Christine Jones notes that
The rates of soil formation provided in the scientific literature usually refer to the weathering of parent material and the differentiation of soil profiles. These are extremely slow processes, sometimes taking thousands of years. Topsoil formation is different and can occur rapidly under appropriate conditions. . . .
The late P.A. Yeomans, developer of the Keyline system of land management, recognised that the sustainability of the whole farm was dependent on living, vibrant topsoil. The formation of new topsoil using Keyline principles, at rates not previously considered possible, was due to the use of a tillage implement designed to increase soil oxygen and moisture levels, combined with a rest/recovery form of grazing and pasture slashing, to prune grass roots and feed soil biota, especially endemic earthworms. Yeomans was able to produce 10 cm of friable black soil within three years, on what was previously bare weathered red shale on his North Richmond farm (Hill 2002).
Bennett (1939) calculated a rate of topsoil formation of just over 11 t/ha/yr (4.4 t/ac/yr) for soils in which organic material was intermixed into surface layers. In situations where plant root mass is high, rates of topsoil formation of 15-20 t/ha/yr (6-8 t/ha/yr) have been indicated (Brady 1984). Healthy groundcover, high root biomass and high levels of associated microbial activity, are fundamental to the success of any technique for building new topsoil.
If the land management is appropriate, evidence of new topsoil formation can be seen within 12 months, with quite dramatic effects often observed within three years. Many people have built new topsoil in their vegetable or flower gardens. Some have started to build new topsoil on their farms. If you have not seen new soil being formed, make a point of doing so. (Jones 2003:19-20)
Healthy biodiverse grasslands with abundant animal populations provide favorable circumstances for biological soil accumulation and carbon sequestration, including opening soils to air and water, fertilizing soil life and stimulating growth of grasses.
Using current best land management practices, recent research has begun to confirm the importance of studying soil organic carbon accumulation on grasslands. In a 9-year study of bioenergy crops, investigators found that switchgrass and maize stored 50% of their soil organic carbon (SOC) below 30 cm (1 foot), up to 4 times more than used in models in use at that time (Follett 2012:866):
Most of the research on SOC in agricultural production systems focused on C in the 0 to 30 cm depth [22–27]. A few studies in which soil sampling has been conducted at greater depths indicate that production agriculture affects soil C deeper in the soil profile [28,29]. (Follett 2012:867)
Concerns about soil depth measurement are not new [Liebig 2008]; depth of soil measurement in estimating soil carbon storage potential is a significant issue. Conventional soil science, which largely addresses agricultural soils managed in industrial agricultural contexts, typically measures soil carbon down to around 30 – 40 cm (12 – 16 inches). Yet roots of native prairie plants may reach 5 times that depth (see Fig. 1), storing carbon in stable molecules for centuries and millennia as long as the soils are undisturbed and not exposed to light or desiccation. [See Soils section.]
Methodological issues for assessment of SOC have thus been problematic, and have likely led to serious overall underestimation of soil sequestration capacity. This is particularly relevant because these soils will not be able play their appropriate critical role in addressing climate until mainstream science and policy recognize and promote the potential of best practices in land management in all ecosystems. Harper & Tibbett found up to five times more soil carbon in Australian soils at depths greater than 1 meter (~3 feet) than is conventionally estimated:
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths (Table 1), it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5 m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm −2 or 21 % of the total store to bedrock. [Harper 2013: 645]
We discuss the dynamics of water cycling and forests elsewhere, but it is worth noting a recent paper suggesting that a more holistic view of ecosystem dynamics is in order. A paradigm shift prioritizing water over carbon as the driving climate force more accurately and effectively guides climate recovery strategies and offers more tactical and regenerative options [Ellison 2017; s.a. Schmidt 2017]. Water is more tangible to most people than carbon, and water recovery is more visible and rapid, offering hope and encouragement in a generally grim scenario. In addition there are numerous other benefits to people and landscapes with improved water management.
THE IMPORTANCE OF ANECDOTAL EVIDENCE
While anecdotal evidence is often disdained in academic science, in many scientific pursuits it forms the foundation of future study. This is especially true in such predominantly observational pursuits such as naturalist biology, ecology, rangeland science and agronomy, where study of the visible vagaries of the natural world inspires questions that may reach far beyond what a happenstance occurrence would imply. It is the weight of such observations that leads to formal protocols, hypotheses and theories to explore the details of a field. Isolating variables can be a very helpful tool; unfortunately, modern science practice has generally lost sight of systems contexts, and that system behavior can be very different from the behavior of any of its isolated parts.
Therefore, it is essential to embrace both analytical and holistic evidence in order to build a full understanding of how environmental systems work as wholes. Together these complementary approaches provide a more comprehensive picture of the systems in question, as well as much clearer guidance for how to proceed in current global ecosystem urgencies.
A small selection of a growing literature of informative anecdotal reports of grassland eco-restoration is included among formal studies below. They reflect the extensive experience of farmers, ranchers and other land managers, and demonstrate the potential positive effects of regenerative land management and eco-restoration on climate and the biosphere as a whole. See for example Stigge 2016, Oppenheimer 2015, Byck 2014, Brown n.d., Brown 2016.
Grassland Article Summaries
Byck 2014. This 12-minute video relates the experiences of three ranchers who manage cattle and land according to regenerative land management principles. They discuss their transition to Holistic Planned Grazing, where for two of them, in areas of ~15 inches of rainfall, their alternative had been bankruptcy. The video vividly illustrates the benefits of cover-cropping and organics, the improvement in lifestyle and economics, and the dramatic improvement in biodiversity and water management. [Byck 2014]
Follett 2012. A USDA study found unexpectedly high quantities of soil organic carbon (SOC) between 30-150 cm (1-5 ft) below the surface, exceeding 2.25 tC/ha/yr (0.9 tC/ac/yr). Ausmus reports that
. . . a 9-year project that evaluated the effects of nitrogen fertilizer and harvest treatments on soil organic carbon sequestration in switchgrass and no-till maize crops managed for biofeedstock production [found that] more than 50 percent of the soil carbon was found between 1 and 5 feet below the soil surface. The average annual increase of soil organic carbon throughout the first 5 feet of subsoil also exceeded 0.9 tons per acre per year [Ausmus 2014: 4-5].
Of interest were the difficulties the authors faced in getting the study through peer review and published since their results were so unexpected, as reported in Ausmus 2014.[21] It originally appeared in Bioenergy Research in 2012. [Follett 2012].
Note too that the Follett study was performed on already degraded soils, “Perennial grasses could be used as bioenergy crops on about 20 million ha (ha = 10,000 m2 or 2.5 acres) of marginal or idle cropland in the USA alone [18]” (Follett 2012:867). [Emphasis added.] A well-supported inference is that healthy, biodiverse soils will yield even better results.
Degraded soils may be less effective carbon sinks than virgin soils even though they have lost most of their carbon because the soil life that creates long-lasting stable carbon molecules is damaged or destroyed by synthetic inputs, tilling and other forms of mismanagement. Nonetheless, results were dramatic:
In the first 9 years of a long-term C sequestration study in eastern Nebraska, USA, switchgrass and maize with best management practices had average annual increases in SOC per hectare that exceed 2 tC/yr for the 0 to 150 cmsoil depth. For both switchgrass and maize, over 50 % of the increase in SOC was below the 30 cm depth. SOC sequestration by switchgrass was twofold to fourfold greater than that used in models to date which also assumed no SOC sequestration by maize. (Follett 2012:866) . . . .
Our results clearly show that significant amounts of C were sequestered deep in the soil profile by switchgrass grown and managed as a biomass energy crop and maize grown continuously in a no-tillage production system for the cultivar Trailblazer array of N fertility and harvest treatments for a 9-year period. For almost all other C sequestration reports used in bioenergy models, studies designed for other purposes were adapted to obtain soil C sequestration estimates, initial soil samples were not available, and control samples were from adjacent fields or non-treatment areas. Our results are supported by similar results reported by Liebig et al. [28] for four switchgrass fields managed with uniform N rates and harvest treatments for 5 years in the USA western Corn Belt. They are supported by the recent work of Varvel and Wilhelm [29] for maize in which significant increases in soil C occurred in soil layers up to 150 cm in depth in maize no-till plots as compared to tilled plots. The soil C that is sequestered deeper than 30 cm is expected to be more stable over time since it is below the tillage zone. Even in the top 30 cm of soil, sequestered C may be stable for extended periods in no-till production systems as reported previously by Follett et al. [33]. (Follett 2012:873)
Harper 2013. This study suggests that the standard sampling depth of 30cm vastly underestimates the global store of soil organic carbon, and thus, presumably, the potential of future soil carbon storage that could result from eco-restoration efforts. “Hypothesizing that SOC retained in soils below the top half metre (in highly weathered deep profiles) would account for the major proportion of SOC in the landscape” [Harper 2013: 642], this southwestern Australia study took deep soil samples in 38 spots across 5 locations. They found that 79% of total carbon store to bedrock occurred below a half meter depth, and 41% occurred below 5 meters depth. “There are two possible sources for the deep carbon; that produced in situ by roots or dissolved carbon that has moved downward from nearer the surface” [Harper 2013: 645]. The sample sites were recently reforested or were under agriculture, and previously had been “covered in a range of xerophytic plants, with root systems that extended to depths of 40 m, such as reported for a Eucalyptus marginata forest” [Harper 2013: 642]. The authors note that more research is needed to understand how deep SOC is affected by land-used changes and climate change.
When the SOC storage within the deep profiles was compared with what would have been reported from conventional sampling depths, it is clear that considerably more SOC was stored in the soils than is normally reported. Across all samples, the surface 0.5m, which is deeper than the standard IPCC sampling depth of 0.3 m (Aalde et al. 2006), contained 5.8± 0.57 kgCm−2 or 21 % of the total store to bedrock. If this is adjusted to 0.3 m depth, using an exponential function based on the samples in the surface metre, the value decreases to 5.6 kgCm−2. For the individual sites this ranged from 3.6 to 8.0 kgCm−2, or 14 –37 % of the total store. …the surface 5 m contained 16.3±1.38 kgCm−2 or 59 % of the total store to bedrock, with this proportion varying from 47 to 77% across the five sampling locations. The amount of carbon stored in the soils can also be contrasted with the biomass carbon storage of 11.0–16.0kg Cm−2 expected at equilibrium following reforestation for these sites (Harper et al. 2007) and likely previously removed from the sites by deforestation in advance of agriculture [p.645].
Liebig 2008. A study of switchgrass for bioenergy found rates of SOC (Soil Organic Carbon) increase of up to 2.75 tC/ha/yr (1.1 tC/ac/yr) when measured to depths of up to 120 cm (4 ft). “In this study, switchgrass significantly affected change in SOC. . . Across sites, SOC increased significantly at 0–30 cm (1 ft) and 0–120 cm (4 ft), with accrual rates of 1.1 and 2.9 tC/ha (0.44 and 1.16 tC/ac), respectively.” [Liebig 2008:215] This indicates the chronic underestimation of soil carbon capacity in the many studies which by convention only measure SOC down to 30 cm (1 ft).
Machmuller 2015. On intensively grazed former row-crop agricultural land converted to dairy farms in the Southeastern U.S., Machmuller et al. found many improvements in the sandy soil, including ~1.25 tC/ha/yr (~0.5 tC/ac/yr) sequestration after accounting for ruminant methane emissions. The study “sought to determine how fast and how much soil C accumulates following conversion of row crop agriculture to management-intensive grazed pastures in the southeastern United States. . . . The highest rates of belowground C accumulation occur when land is converted to grassland ecosystems” [Machmuller 2015: 2]. These intensively grazed managed systems led to an approximately 75% increase in soil carbon within six years,
[a] high C accumulation rate [that] stems from year round intensive forage/grazing management techniques on sandy soils with an initially low soil C content due to past conventional-till row crop agriculture. . . . These forage-management techniques are precisely those suggested to increase SOM in pasture systems and when they are applied to soils with degraded SOC content, such as soils in the southeastern United States, rapid C accumulation ensues. . . .
On the basis of a whole farm C-cycle analysis, C accumulation appears to offset methane emissions during the rapid soil C accumulation phase . . . As the C accumulation rate declines these farms will become net C-emitting—similar to all dairy production—because of ruminant methane emissions. However, the substantial soil-quality benefits of higher organic matter remain and will likely increase the sustainability of dairy production using management-intensive grazing [Machmuller 2015:3].
The eventual methane emissions may be markedly less than suggested, however, since the report did not consider methane breakdown from methanotrophic bacteria and atmospheric hydroxyl radical oxidation.
The authors conclude
that pasture-based intensively grazed dairy systems may provide a near-term solution for agricultural lands that have experienced soil-C loss from previous management practices. Emerging land uses, such as management-intensive grazing, offer profitable and sustainable solutions to our needs for pairing food production with soil restoration and C sequestration. [Machmuller 2015: 2-3]
McCosker 2000. A discussion of the introduction of what the author calls “cell grazing,” framed as a paradigm shift in Thomas Kuhn’s terms [Kuhn 1962] over the years 1990-1999. McCosker reviews the dichotomy between researchers and producers, and travels to see actual results in the U.S., Zimbabwe, Namibia, and South Africa. He states, “Only after seeing the outcomes time and again in all possible environments was I finally convinced that the principles could not be faulted.” This kind of paradigm shift has been experienced repeatedly as practitioners must develop the courage to challenge prevailing assumptions in order to take the requisite transitional steps.
Oppenheimer 2015. Farmers Paul and Elizabeth Kaiser own eight acres in Sebastopol in Northern California and they farm three of them, developing a model that other farmers are beginning to use.
[Kaiser] farms a mere eight acres, and harvests fewer than three of them. Nonetheless, his methods are at the forefront of a farming movement that is so new (at least in the U.S.), and so built for a climate-changed world of diminishing rains, that it opens up gargantuan possibilities. One might call this methodology sustainability on steroids, because it can generate substantial profits. Last year, Kaiser’s Sonoma County farm grossed more than $100,000 an acre, which is 10 times the average per-acre income of comparable California farms. This includes Sonoma’s legendary vineyards, which have been overtaking farmland for decades, largely because wine grapes have become much more lucrative these days than food, at least the way most farmers grow it.
Kaiser manages all of this without plowing an inch of his ground, without doing any weeding, and without using any sprays—either chemical or organic. And while most farmers, even on model organic farms, constantly tinker with various fertilizer cocktails, Kaiser concentrates on just one: a pile of rotten food and plants, commonly known as compost, and lots of it. Kaiser then adds this compost to a rare blend of farming practices, both old and new, all aimed at returning dirt to the richest, most fertile seedbed possible.
They use permaculture, agroforestry and other intensive techniques, have built deep, healthy soils by keeping the ground covered and spongy to capture water and carbon, all of which provide solid protection from droughts and floods.
Retallack 2001, 2013. “Grassland expansion initiated increased organic C storage in soils, soil water retention, speed of nutrient exploitation, surface albedo, and C burial in sediments eroded from their soils. These changes had many consequences, including long-term global cooling.” [Retallack 2001:422] and “This climatic zone is not only the most widespread, but also the most fertile region of our planet.” [Retallack 2013:78] The paleohistory of grasslands provides the basis for considering the potential of grasslands as huge biological carbon sinks that may be realized again with regenerative land management
Rodale Institute 2014.
Rodale reports that regenerative grazing practiced on a global scale could sequester 71% of annual emissions of 14 Gt C/yr. Combined results from regenerative grazing and agriculture techniques could, if practiced globally, lead to a net reduction of atmospheric carbon dioxide of 1.7 gigatons per year, or 0.85 ppm/year. This represents a potential sequestration of approx. 3t C/ha/yr (1.2t C/ac/yr) on grasslands and croplands. [Rodale Institute 2014:9] This does not include the considerable contributions of non-agricultural lands, nor recent developments in intensive regenerative practices such as permaculture and biochar.
Moreover, Rodale’s side-by-side trial after 30+ years showed that, following the three-year transition period, organic yields match conventional yields, except in drought years, when organic yields surpass conventional yields. Furthermore, energy input and greenhouse gas emissions were lower in organic systems, and profits were higher.
Rowntree 2016. Examining ruminant methane and net carbon sequestration for grassfed beef in a systems context, Rowntree found net sequestration rates of up to 2.11 tons/ha/yr (0.84 tC/ac/yr) for non-irrigated, lightly stocked grazing.
Therefore, including soil carbon sequestration (SCS) potential brings the differences in grazing environments into focus, and significantly changes the outcome. When comparing two well-managed grazing strategies, grass-finished (MOB) and conventional (IRG), each strategy could be an overall carbon sink, but the MOB grazing would only need sequester half as much carbon (1 tC/ha/yr) as IRG grazing (2 tC/ha/yr) for a net zero greenhouse gas footprint. Methane emissions were similar in both grazing environments, but MOB grazing offered significant benefits in increased carbon sequestration. [Rowntree 2016:36]
It is unlikely that such SCS would take place in the absence of a healthy biodiverse ecosystem, one that is supported through MOB grazing. The result is higher net methane emissions under conventional grazing practices. The opposite occurs with properly managed grazing practices, where grasslands as a system actively build more soil carbon for years, leading to a net increase in soil carbon despite enteric methane production by ruminants.[22]
The recent call for improved management of grazing systems as part of an international climate change mitigation strategy is critical, particularly in light of many existing beef LCAs [Life Cycle Assessments] that have concluded that beef cattle produced in grazing systems are a particularly large sources of GHG emissions. To identify the best opportunities to reduce GHG emissions from beef production, a systems approach that considers the potential to increase soil C and reduce ecosystem-level GHG emissions is essential… [W]e generated an LCA that indicates highly-managed grass-finished beef systems in the Upper Midwestern United States can mitigate GHG emissions through SCS while contributing to food provisioning at stocking rates as high as 2.5 Animal Units (AU) per hectare. From this data, we conclude that well-managed grazing and grass-finishing systems in environmentally appropriate settings can positively contribute to reducing the carbon footprint of beef cattle, while lowering overall atmospheric CO2 concentrations. [Rowntree 2016:36]
Schwartz 2013. Cows Save the Planet was a landmark book that helped launch the regenerative agriculture and eco-restoration climate movements. The author gathers stories from practitioners around the world and paints a picture of broad possibilities for addressing global warming, floods, droughts, desertification, malnutrition and many other seemingly unrelated problems that have a single common cause: human mismanagement of lands across the planet. An excellent introduction for a general readership as well as for scientists unfamiliar with the potentials that nature provides.
Shinn n.d. Ridge Shinn is a rancher in Central Massachusetts (Big Picture Beef) who introduced Allan Savory’s methods to the state. For over ten years he has been investigating the holistic context for bringing healthy meat through the local supply chain, and its relationship to human health and global warming. He states,
Since the 1990’s, science has discovered important connections between rotational grazing, soil health, and healthy food. Big Picture Beef’s methods for raising 100% grass-fed cattle offer huge benefits for the environment and for society. The long term goal of the program is Northeast beef for Northeast markets, carbon sequestration, soil fertility and biodiversity, energy savings, and a revitalized rural economy.
Teague et al. 2016. In a review of the literature, the authors conclude that regenerative conservation cropping and adaptive multi-paddock grazing can turn agricultural soils from a carbon source in conventional agriculture into a carbon sink at rate of ~3 tC/ha/yr (~1.2 tC/ac/yr). Key factors include the use of no-till, cover crops, managed grazing, organic soil amendments and biotic fertilizer formulations. These practices can result in elimination of soil erosion and loss, the greatest agricultural contribution to global warming (1 Gt C/yr). Benefits may include “increased water infiltration, improved water catchment, greater biodiversity, increased ecosystem stability and resilience, and improved C sequestration.” [Teague 2016:158]
Conclusion
These reports demonstrate the promising potential for storage of massive amounts of soil carbon to address both the atmospheric and eco-destruction aspects of climate, along with dramatic improvements in ecosystem health using regenerative approaches to grassland management.
Forests
Note: As mentioned in the Release notes, we have a small staff, and therefore have had to postpone some important material for the next release, scheduled for January 2018. This is particularly true of forests and we will include a more thorough examination of their importance in addressing climate moving forward. Nonetheless, we felt that the investigations here were innovative and interesting, and we wanted to make them available to our readers sooner rather than later.
Forests cover nearly 31% of Earth’s total land area [FAO 2016], and remain one of the major terrestrial ecosystems on the planet. Forests play a significant role in the global ecosystem through cooling, evapotranspiration, covering/shading/sheltering, providing fuel and fiber, aiding cloud formation, and creating wind. Because global forests and wooded lands store an estimated 485 Gt of carbon, forest conservation and afforestation are recognized in the United Nations Framework Convention on Climate Change (UNFCCC) as key strategies for climate change mitigation [UNFCCC 2017].
Despite this acknowledgment, “for the world as a whole, carbon stocks in forest biomass decreased by an estimated 0.22 Gt annually during the period 2011–2015. This was mainly because of a reduction in the global forest area” [UNFCCC 2017].Indeed, humanity has been in the business of clearing forests for thousands of years, and this continues today. However, rapid reductions in deforestation could abate further carbon emissions and thus extreme results of climate change. Moreover, reductions in deforestation and implementation of agroforestry practices together could restore biodiversity in damaged ecosystems, repair local and global water cycles, and, ultimately, help restore carbon levels to pre-industrial levels. Here we present several articles illustrating the impact of forests on global climate, as well as the potential for restorative afforestation and agroforestry practices to sequester large amounts of carbon.
Forest Article Summaries
Ellison 2017. This paper takes the innovative and paradigm-shifting position that carbon is not the primary consideration in climate; rather, it is water that should be a central focus in assessing climate processes and effects. It considers forests from a systems perspective.
Forest-driven water and energy cycles are poorly integrated into regional, national, continental and global decision-making on climate change adaptation, mitigation, land use and water management. This constrains humanity’s ability to protect our planet’s climate and life-sustaining functions. The substantial body of research we review reveals that forest, water and energy interactions provide the foundations for carbon storage, for cooling terrestrial surfaces and for distributing water resources. Forests and trees must be recognized as prime regulators within the water, energy and carbon cycles. If these functions are ignored, planners will be unable to assess, adapt to or mitigate the impacts of changing land cover and climate. Our call to action targets a reversal of paradigms, from a carbon-centric model to one that treats the hydrologic and climate-cooling effects of trees and forests as the first order of priority. For reasons of sustainability, carbon storage must remain a secondary, though valuable, by-product. The effects of tree cover on climate at local, regional and continental scales offer benefits that demand wider recognition. The forest- and tree-centered research insights we review and analyze provide a knowledge-base for improving plans, policies and actions. Our understanding of how trees and forests influence water, energy and carbon cycles has important implications, both for the structure of planning, management and governance institutions, as well as for how trees and forests might be used to improve sustainability, adaptation and mitigation efforts. [Ellison 2017: Abstract]
Ford 2017. Structural Complexity Enhancement (SCE) is part of a larger ecological concept: nature tends to complexity, providing its resiliency, flexibility and inventiveness. SCE in treatment of forests is a management approach that promotes development of late-successional structure, including elevated levels of coarse woody debris. It adds variety to tree ages (favoring older trees), and variations in available sunlight and habitat.
Large trees, previously assumed to slow in both productivity and growth rate (Weiner and Thomas 2001, Meinzer et al. 2011), function as long-term carbon sinks (Carey et al. 2001). These findings further support the significance of structural retention as a co-benefit to forest carbon storage. Adaptive silvicultural practices promoting multiple co-benefits, for instance, by integrating carbon with production of harvestable commodities, can contribute to efforts to dampen the intensity of future climate change while maintaining resilient ecosystems (Millar et al. 2007). Prescriptions that enhance in situ forest biomass and thus carbon storage offer one such alternative (Ducey et al. 2013). U.S. forests currently offset approximately 16% of the nation’s anthropogenic CO2 emissions, but this has the potential to decline as a result of land-use conversion and lack of management (EPA 2012, Joyce et al. 2014). While passive or low-intensity management options have been found to yield the greatest carbon storage benefit, assuming no inclusion of substitution effects (Nunery and Keeton 2010) or elevated disturbance risks (Hurteau et al. 2016), we suggest the consideration of SCE to enhance carbon storage. Multiple studies have explored co-benefits provided by management for or retention of elements of stand structural complexity, including residual large living and dead trees, horizontal variability, and downed CWM (Angers et al. 2005, Schwartz et al. 2005, Dyer et al. 2010, Gronewold et al. 2012, Chen et al. 2015). Silvicultural treatments can effectively integrate both carbon and late-successional biodiversity objectives through SCE based on this study and previous research (e.g., Dove and Keeton 2015). Remaining cognizant of the potential for old-growth compositional and structural baselines to shift over time and space with global change—climate impacts on forest growth and disturbance regimes, altered species ranges, and the effects of invasive species—will be important for adaptive management for late-successional functions such as carbon storage. [Ford 2017: 16]
Healing Harvest Forest Foundation.
The spot compaction of animal feet is far less damaging to the forest soil and tree roots than the continuous track created by a wheel or track driven machine. Small sized tracts of timber can not be harvested with conventional methods that require higher capitalization and expensive moving cost. The economic pressure in conventional forest harvesting operations influences most loggers to feel that they must cut all the trees to make their work cost effective. This restricts the silvicultural prescriptions available for the management of the forest….Our method of selecting individual trees on a “worst first” basis and limiting removal to no more than 30% retains the forested condition and is indeed improvement forestry…. The holes created in the forest canopy are substantial enough for “shade intolerant” species to regenerate naturally from seedlings of the superior specimens that are left in a healthy “good growing” condition. We believe that basically the repair of the forest from previous “high grading” is best accomplished through several successive “low grading” harvests.[Healing Harvest 1999]
Makarieva 2007. The authors examine ecological and geophysical principles to explain how land far inland away from the ocean can remain moist, given that gravity continuously pulls surface and groundwater into the ocean over time.
All freshwater on land originates in the ocean from which it has evaporated, is carried on air flux, and precipitates over the land. Coastal regions benefit from this cycle by their proximity to the ocean, yet in the absence of natural forests in coastal regions precipitation weakens as distance from the ocean increases, leaving inland areas arid. The authors propose the concept of a biotic pump to explain how large continents can be sufficiently moist deep into the interior and abundant with rivers and lakes.
Air and moisture are pulled horizontally by evapotranspiration from coastal forests. When water vapor from plants condenses, it creates a partial vacuum which pulls water evaporating from the ocean into the continental interior where it rains in forest. By contrast, deserts are unable to pull ocean evaporation to them because they lack any evaporative force.
Such ongoing deforestation, and crucially coastal deforestation on a large scale, threatens to cut off rain to the interiors of Earth’s continents thereby creating new deserts. The Amazonian rainforest is the prime example. Deforestation of the eastern coast of South America has led to changes in the rainforest that is resulting in drying and desertification of the interior, with unprecedented fires and loss of rivers. Historically, Australia’s interior became a desert around the time the first humans arrived on the continent, and the authors speculate that early coastal deforestation was the cause. On the other hand, restoring natural coastal forests can also restore inland water cycles and reverse desertification.
This article illustrates the importance of biological relationships that are ecologically complex and poorly understood. It highlights the significance of the precautionary principle in assessing what we don’t know (and what we don’t know that we don’t know) when altering ecological processes, and taking preventive action in the face of uncertainty.
Going back almost 4 billion years, a scant half-billion years since the formation of planet earth from cosmic dust, life began to appear. It persisted through eons of celestial, tectonic and climatic upheaval. Around a billion years later, life, in the form of microbes, found the driver’s seat and has taken over the world ever since. In an anthropocentric culture that creates gods in its own image, we are not generally aware that millions of species of living things have molded this planet, turned it blue and green, and created most of its features, from an oxygen atmosphere to geological formations to proliferation of millions of other kinds of living things. Without life, Earth would be merely another rock flying through space, like Mars or Venus.
The power of life is especially important in discussions of and action on climate change, since mainstream climate science views living things as victims of global warming, not primary drivers of potential climate solutions, as mentioned above. This is most unfortunate since our current obsession with greenhouse gas emissionsas a root cause of climate disruption has led us to a dead end. For even if we were to go to zero emissions immediately, due to positive feedback loops and a seriously degraded biosphere, climate chaos would likely continue to accelerate and rage out of control.[3],[4] It is therefore not unreasonable to pursue the possibility that living things are able to remove the requisite carbon from the atmosphere, cool the biosphere as well as return biodiverse life to dead and dying ecosystems, and in fact there is ample evidence that such is the case – as we shall see.
In order to re-evaluate our approach to climate change in the anthropocene and to find solutions in addition to reducing carbon emissions, we do well to consider the situation from a systems perspective. That is, to acknowledge that we’re not simply dealing with recent energy imbalances disrupting millions of years of relative stability in planetary temperatures, and to acknowledge as well that the prevailing belief that these changes are driven primarily by geophysical phenomena unrelated to biological systems is most likely erroneous. To the contrary, we emphasize the point that anthropogenic global warming is an extremely complex phenomenon, a function driven primarily by the biology of all kingdoms of life.
There is no question that planet earth is seriously warming at accelerating rates; however, it is increasingly apparent that the greenhouse gas premise of climate science is problematic. That global warming is simply a consequence of atmospheric concentrations of greenhouse gases, especially carbon dioxide, is a persistent hypothesis that has its origins in the modern physical sciences beginning around two hundred years ago, although climate-related observations were made by the ancient Greeks and undoubtedly by many hunter-gatherer cultures. This greenhouse gas hypothesis must now be subject to closer examination.
It is well established that greenhouse gases do trap heat in the earth’s atmosphere. Furthermore, the resultant energy imbalance in the form of trapped heat from the burning of fossil fuels has increased greenhouse gas concentrations and planetary warming. However, global warming is largely treated as a geophysical phenomenon isolated from planet earth as a living, dynamic system. Isolating variables, a primary tool in mainstream science, often leads to overlooking systems behavior that bears no resemblance to the behavior of any of its parts.
Biological systems are generally regarded as victims of global warming; they are less generally regarded as the primary potential solution. This view is starting to change, but slowly, whereas global warming is a pressing emergency not allowing for the normal course of a paradigm shift to take place over at least one generation if not more.
We must therefore develop a new paradigm, that of global warming as a phenomenon of biology and ecosystem function. From this perspective we would regard greenhouse gases as a symptom of human mismanagement and destruction of billions of acres of land and waters – a highly problematic symptom, to be sure, but not the root cause. Such a perspective brings an entirely new set of potential solutions to bear, powerful land management solutions well within our grasp, as we seek to demonstrate.
The challenge in promoting eco-restoration as the primary approach to addressing global warming is that the scientific literature supporting such efforts is diffuse, spread across many disciplines, sometimes with few obvious connections. There currently exists no dedicated journal that intentionally collects studies from fields as disparate as rangeland science, paleontology, soil science, microbiology, agronomy, evolutionary biology, mycology, entomology, oceanography, limnology, and many many others – not to mention human psychology – and relates findings directly to climate, its effects, mechanisms, and solutions.
This Compendium will begin to look at some of the elements of planetary climate and systems function, and attempt to weave them together to create a more comprehensive systems view. We attempt to offer opportunities for many different and powerful nature-based approaches for dealing with changes in planetary function, particularly climate.
There are a number of assumptions that we need to reconsider if we are to see our way clear to a new, more effective climate paradigm. Once we have drawn a picture of the new paradigm and the previously unrecognized connections among investigations that this paradigm enables, the research and data will be positioned to acquire new meaning, sense and purpose.
Going back almost 4 billion years, a scant half-billion years since the formation of planet earth from cosmic dust, life began to appear. It persisted through eons of celestial, tectonic and climatic upheaval. Around a billion years later, life, in the form of microbes, found the driver’s seat and has taken over the world ever since. In an anthropocentric culture that creates gods in its own image, we are not generally aware that millions of species of living things have molded this planet, turned it blue and green, and created most of its features, from an oxygen atmosphere to geological formations to proliferation of millions of other kinds of living things. Without life, Earth would be merely another rock flying through space, like Mars or Venus.
The power of life is especially important in discussions of and action on climate change, since mainstream climate science views living things as victims of global warming, not primary drivers of potential climate solutions, as mentioned above. This is most unfortunate since our current obsession with greenhouse gas emissionsas a root cause of climate disruption has led us to a dead end. For even if we were to go to zero emissions immediately, due to positive feedback loops and a seriously degraded biosphere, climate chaos would likely continue to accelerate and rage out of control.[3],[4] It is therefore not unreasonable to pursue the possibility that living things are able to remove the requisite carbon from the atmosphere, cool the biosphere as well as return biodiverse life to dead and dying ecosystems, and in fact there is ample evidence that such is the case – as we shall see.
There is substantial evidence that we can address the climate crisis by intensive global eco-restoration: drawing down vast amounts of carbon from the atmosphere into global soils through photosynthesis; managing water cycles to cool the biosphere; restoring biodiversity and degraded terrestrial and aquatic ecosystems.
Support for an eco-restoration hypothesis is solid and comes from a wide variety sources, both in academic science and modern and traditional land management practice. Eco-restoration may be applied in numerous ecosystems: croplands/agroecosystems; estuaries; forests; marine ecosystems; shorelines; pastures and rangelands; wetlands; and others.
One of the challenges at this point in time is to collect available evidence from sources spread across many disciplines, in different formats, synthesize it, and present a comprehensive, logical and compelling case that there are practical steps we can take to regenerate large areas of the planet in order to address global warming successfully and rapidly.
In this paper we attempt to connect these disparate sources and create a constructive narrative to move from the current climate paradigm, where global warming is narrowly defined as a problem of excessive greenhouse gases, to a new climate paradigm, where global warming is defined as a systemic problem resulting from global anthropogenic destruction of the natural world.
We include in Appendix A an essential discussion of how paradigms both promote and constrain research and discovery. A key point is that a shift in paradigms opens many positive possibilities for addressing climate through eco-restoration, possibilities that are outside the scope of the current greenhouse gas paradigm. The latter is limited to reducing fossil fuels emissions and has little if any success to date based on annual increases in atmospheric greenhouse gas burdens and rising global temperatures. Notwithstanding technological advances, it furthermore has uncertain future prospects, especially considering the accelerating warming we are seeing today.
We further explore an idea that has been overwhelmed by our current preoccupation with powerful technologies, i.e., that living systems are the most powerful force affecting planet earth throughout the biosphere. Therefore it is in living systems, not technology, where the solutions to global warming reside.
We also address historically healthy natural systems that were bountiful in ways that are mostly lost to modern human experience. Collectively, humans have gradually whittled away at the power of the natural world through environmental overshoot and destruction over hundreds of generations, until widespread environmental collapse – including mass extinction, desertification and global warming – appears inevitable. When we begin to grasp the potential productivity and broad benefits of healthy ecosystems, we discover a hopeful new roadmap for addressing present dilemmas.
We also discuss in Appendix A the conceptual and psychological obstacles to a paradigm shift, as postulated by Thomas Kuhn in his highly influential 1962 book, The Structure of Scientific Revolutions [Kuhn 1962]. It is our hope that Kuhn’s insights will assist in understanding the current scientific and cultural roadblocks and in proceeding with the necessary transitions.
We conclude that it is possible, even this deep into climate, extinction and eco-destruction crises, for successful environmental outcomes for a biodiverse spectrum of species, including Homo sapiens. The challenge is largely overcoming resistance inherent in human dominant culture, including scientific, technological, social, political and economic beliefs. Such resistance is the primary obstacle. Otherwise we can solve these problems with readily available resources and little or no technology, provide for satisfying and productive lives in local habitats worldwide, make ample food and water available to existing populations while reducing both global population and non-essential consumptive behaviors. Not to mention preventing droughts, floods and conflicts over resources, and all for relatively little expense.
While this may all sound too good to be true, these are not separate problems. By solving the one key problem, a natural world in utter anthropogenic disarray, it is possible for all the pieces to fall into place.
We have undertaken a far more ambitious enterprise than we had initially envisioned.
One reason is that, just as in a forest, whenever you turn over a leaf or a log you discover a universe. The universe of knowledge about healing a devastated earth is vast, from indigenous wisdom to systems science and everything in-between.
The second reason is that this body of knowledge is in a phase of exponential growth, as are the life-support issues that we attempt to address. It is virtually impossible to keep up with the almost daily discoveries and surprises, some terrifying, some extraordinarily hopeful.
We have a small staff, and therefore have had to postpone some very important material for the next release, scheduled for December 2018. We expect that these under-represented areas of discussion, which also hold great potential for addressing eco-devastation in general and climate in particular, will receive a more comprehensive review: forests and wetlands; cities and suburbs; oceans; shorelines; microbial life; dynamics of ice; and more (including the surprises we haven’t thought of yet).
This Compendium of Scientific and Practical Findings Supporting Eco-Restoration to Address Global Warming (the “Compendium”) is a fully referenced compilation of the evidence outlining the power, benefits and necessity of eco-restoration to address global warming. Bringing together findings from the scientific literature, government and industry reports, and journalistic investigations, this is a public, open-access document that is housed on the website of Biodiversity for a Livable Climate (https://bio4climate.org/resources/compendium).
Welcome to the First Edition of this Compendium. We are developing and continuing to refine our editorial process, and we invite comments and suggestions from you, the reader, to help make this document as useful as possible.
The Compendium is intended as a living document, and there will be regular additions between releases as the relevant literature evolves. In the past few years there has been a dramatic growth of information that supports the power of the natural world to address the causes and effects of climate change. The climate conversation has expanded dramatically beyond greenhouse gases, and is repositioning global loss of biodiversity and destruction of ecosystems as root causes of global warming and many associated environmental problems.
There is no central field of study that aggregates the information relevant to eco-restoration and climate; there are, however, many fields that contribute. The result is a solid body of evidence that argues compellingly for a focused effort on the part of governments, civic organizations, NGOs and, especially, local communities and individuals to take the lead on regenerating degraded and desertified land and waters worldwide, while also preserving carbon-rich wetlands, coastal seagrasses and other vital intact ecosystems.
The evidence is abundant, and our goal is to begin to gather it in one place to make it readily available for public scrutiny. We will present information from papers in the peer-reviewed literature, non-profit organizations, government bodies, commercial publications, and the popular press. All have valuable contributions to make from different perspectives that together paint a picture of a new, healthy and attainable world, a portrait of the people who are helping us to get there, how to get there using nature’s low-tech tools, and the arguments that propel this pressing journey forward.
It behooves us to recognize that an exclusive focus on greenhouse gas emissions is problematic. This is not a statement made lightly, as longstanding bodies of knowledge should not be dismissed capriciously. And yet, when paradigms fail to reconcile reality with assumption, they should be retired in the service of scientific progress.
This is the situation we are in today with two competing though not mutually exclusive paradigms (worldviews):
Old paradigm: Climate change is primarily a consequence of elevated greenhouse gas emissions, largely from the burning of fossil fuels.
Compared to
New paradigm: Climate change is a consequence of global ecological destruction, especially of soils, soil biota, above-ground flora and fauna, and bodies of water; including disruptions of carbon, water, and energy cycles.
It’s important to note that the old paradigm has provided many important insights critical to the new paradigm, and we should learn from the investigations into the old paradigm even as the paradigm as a whole may come into question. We need to work together towards a common goal, a healthy and abundant planet, while keeping in mind that each paradigm leads to a very different set of outlooks, studies, behaviors and outcomes.
Evidence for paradigm shifts builds slowly, acceptance even more so. Indeed, one of the problems we face with this Compendium is that we’re not just considering changing land management practices, we’re of necessity examining and questioning some of the core assumptions of current mainstream science. We are facing a culture shift of difficult proportions.
We are aware that there are many studies that present significantly lower estimates of the potential for building soil carbon, managing water cycles, and eco-restoration in general. It is our position that for the most part, while those studies may present useful data, their perspective is limited by mechanistic assumptions and reductionist, non-systemic methodologies. Therefore we mostly do not include these studies in this Compendium (some, however, are illustrative and helpful).
This is admittedly a bias on our part, an intentional one aimed at offsetting the virtually ubiquitous bias of the mainstream paradigm. The reader may readily peruse the mainstream literature for extensive review, and we welcome critical examination of both the mainstream literature and of the literature presented here in an effort to promote the best possible outcomes for biodiverse life on Earth.
In sum, the clear intent of this Compendium is to fortify the case for eco-restoration as a primary and essential solution to global warming, one that potentially yields benefits more quickly and safely than any other solution currently being proposed, and to move it forward with all due haste.
Finally, we’re not attempting a definitive “proof,” an elusive pursuit in a scientific arena in any case. Rather, we’re presenting evidence of real and practical possibilities, along with solid research from many disparate fields, some of which are newly discovering (with occasional surprise) that they’re related in mutually productive ways in a kind of scientific symbiosis.
It is truly time to move science and practice beyond present assumptions. We provide examples from a variety of regenerative approaches that illustrate how we may expand the current boundaries of mainstream evidence and paradigms – and perhaps even use our innovative and growing practical and scientific understandings to reverse global warming.[1]
Finally, we invite our readers to submit summaries of relevant findings for the next release of this Compendium. Please e-mail all contributions and correspondence to compendium@bio4climate.org.
About Biodiversity for a Livable Climate
Biodiversity for a Livable Climate, bio4climate.org, is a 501(c)(3) non-profit founded in 2013 whose mission is to support the restoration of ecosystems to reverse global warming. We are:
A think tank, creating research and reports (such as this Compendium), and presenting conferences on the science and practice of eco-restoration with speakers from around the world.
An educational organization, offering presentations, courses and materials, including over 170 videos of speakers from our 9 conferences since November 2014 (bio4climate.org/conferences), with many restoration and climate-positive examples from both scientists and practitioners.
An advocate that reaches out to other organizations to encourage and facilitate the incorporation of eco-restoration as a climate solution into their own messaging and actions. We seek to connect to other groups and projects to help nourish and advance their own growth in a healthy direction, and carry messages among groups to collaboratively learn and build on each other’s efforts, and occasionally facilitate the emergence of new groups. Since climate affects everyone, every organization has to deal with it in its own way, and we help with the transition.
An activist groupthat engages in non-partisan political processes. For example, we helped shepherd a bill through the legislative process in 2017 to establish a Maryland Healthy Soils Program and are pursuing similar efforts in the Massachusetts legislature.
We are a small 501(c)(3) non-profit with a major impact in addressing climate, and we rely on your generous contributions! Please go to www.Bio4Climate.org/Donate to join our monthly donor program, or to make a one-time donation, all tax deductible. Many thanks!
Suggested Citation
Compendium of Scientific and Practical Findings Supporting Eco-Restoration to Address Global Warming, https://bio4climate.org/resources/compendium/. This is a collection of references that will grow as new literature becomes available, and as older literature is re-discovered.
Acknowledgements
Current reviewers and contributors to this collection are Hannah Lewis, Adam Sacks, Robert Blakemore, Erica Antill, Andrew Blair, Gina Angiola, Philip Bogdonoff, Annita Seckinger, Paula Phipps and Fred Jennings. The contributions from our many speakers and collaborators cannot be overstated. We invite our readers to review our collection of conference videos on the program page of each of our nine conferences to date.
We are most appreciative of the support from our sponsors over the past three years. In particular, the 11th Hour Project provided significant funding for our first two years, and the new and important institution that it helped create, the Regenerative Agriculture Foundation, is continuing its strong moral and financial support. We are also pleased to acknowledge generous conference sponsorship from the Organic Consumers Association, Regeneration International, the Virgin Earth Challenge, Bristol Community College, and the Tufts Institute of the Environment. Additional important support has been kindly provided by the Nutiva Foundation, the Savory Institute, Irving House and the Bionutrient Food Association.
We also gratefully acknowledge support from several institutions, including Tufts University, Bristol Community College and the University of the District of Columbia.
We have undertaken a far more ambitious enterprise than we had initially envisioned.
One reason is that, just as in a forest, whenever you turn over a leaf or a log you discover a universe. The universe of knowledge about healing a devastated earth is vast, from indigenous wisdom to systems science and everything in-between.
The second reason is that this body of knowledge is in a phase of exponential growth, as are the life-support issues that we attempt to address. It is virtually impossible to keep up with the almost daily discoveries and surprises, some terrifying, some extraordinarily hopeful.
We have a small staff, and therefore have had to postpone some very important material for the next release, scheduled for December 2018. We expect that these under-represented areas of discussion, which also hold great potential for addressing eco-devastation in general and climate in particular, will receive a more comprehensive review: forests and wetlands; cities and suburbs; oceans; shorelines; microbial life; dynamics of ice; and more (including the surprises we haven’t thought of yet).